
Therapeutic Properties of Edible Mushrooms and Herbal Teas in Gut Microbiota Modulation



Abstract
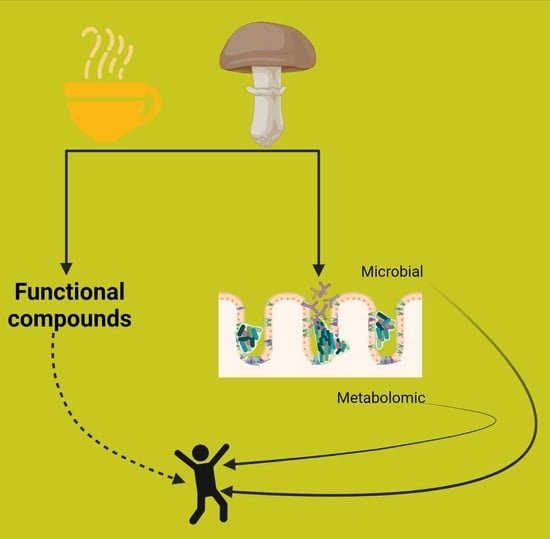
:
1. Introduction
2. Interaction of Mushrooms with Human Microbiota
3. Interaction of Microbiota with Herbal Teas
4. Effect of Plant miRNAs on Gut Microbiota Modulation with Impact on the Human Health
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kesarwani, K.; Gupta, R. Bioavailability enhancers of herbal origin: An overview. Asian Pac. J. Trop. Biomed. 2013, 3, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Kesarla, R.; Omri, A. Formulation Strategies to Improve the Bioavailability of Poorly Absorbed Drugs with Special Emphasis on Self-Emulsifying Systems. ISRN Pharm. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vamanu, E.; Gatea, F.; Sârbu, I.; Pelinescu, D. An In Vitro Study of the Influence of Curcuma longa Extracts on the Microbiota Modulation Process, in Patients with Hypertension. Pharmaceutics 2019, 11, 191. [Google Scholar] [CrossRef] [Green Version]
- Clement, Y.N. Factors affecting the pharmacokinetics of herbal preparations and their impact on the outcome of clinical trials. Focus Altern. Complement. Ther. 2009, 14, 87–91. [Google Scholar] [CrossRef]
- Tang, G.-Y.; Meng, X.; Gan, R.-Y.; Zhao, C.-N.; Liu, Q.; Feng, Y.-B.; Li, S.; Wei, X.-L.; Atanasov, A.G.; Corke, H.; et al. Health Functions and Related Molecular Mechanisms of Tea Components: An Update Review. Int. J. Mol. Sci. 2019, 20, 6196. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Perera, C.; Hemar, Y. Antitumor activity of mushroom polysaccharides: A review. Food Funct. 2012, 3, 1118–1130. [Google Scholar] [CrossRef]
- Spisni, E.; Turroni, S.; Shahaj, S.; Spigarelli, R.; Ayala, D.; Valerii, M.C. Natural Compounds in the Modulation of the Intestinal Microbiota: Implications in Human Physiology and Pathology. Altern. Med. 2020. [Google Scholar] [CrossRef]
- Büntgen, U.; Kauserud, H.; Egli, S. Linking climate variability to mushroom productivity and phenology. Front. Ecol. Environ. 2012, 10, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Vamanu, E.; Gatea, F.; Pelinescu, D.R. Bioavailability and Bioactivities of Polyphenols Eco Extracts from Coffee Grounds after In Vitro Digestion. Foods 2020, 9, 1281. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- MetaHIT. Available online: https://www.gutmicrobiotaforhealth.com/metahit/ (accessed on 11 May 2021).
- Boziki, M.K.; Kesidou, E.; Theotokis, P.; Mentis, A.-F.A.; Karafoulidou, E.; Melnikov, M.; Sviridova, A.; Rogovski, V.; Boyko, A.; Grigoriadis, N. Microbiome in Multiple Sclerosis: Where Are We, What We Know and Do Not Know. Brain Sci. 2020, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-level analysis of gut microbiome variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- McLeod, A.; Zagorec, M.; Champomier-Vergès, M.-C.; Naterstad, K.; Axelsson, L. Primary metabolism in Lactobacillus sakei food isolates by proteomic analysis. BMC Microbiol. 2010, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Drossman, D.A. Functional Gastrointestinal Disorders: History, Pathophysiology, Clinical Features, and Rome IV. Gastroenterology 2016, 150, 1262–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armuzzi, A.; Cremonini, F.; Ojetti, V.; Bartolozzi, F.; Canducci, F.; Candelli, M.; Santarelli, L.; Cammarota, G.; De Lorenzo, A.; Pola, P.; et al. Effect of Lactobacillus GG supplementation on antibiotic-associated gastrointestinal side effects during Helicobacter pylori eradication therapy: A pilot study. Digestion 2001, 63, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, K.; Ozga, A.T.; Warinner, C.; Tito, R.Y.; Obregon-Tito, A.J.; Xu, J.; Gaffney, P.M.; Jervis, L.L.; Cox, D.; Stephens, L.; et al. Gut Microbiome Diversity among Cheyenne and Arapaho Individuals from Western Oklahoma. Curr. Biol. 2015, 25, 3161–3169. [Google Scholar] [CrossRef] [Green Version]
- Sik, B.; Kapcsándi, V.; Székelyhidi, R.; Hanczné, E.L.; Ajtony, Z. Recent Advances in the Analysis of Rosmarinic Acid from Herbs in the Lamiaceae Family. Nat. Prod. Commun. 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hajimehdipoor, H.; Saeidnia, S.; Gohari, A.R.; Hamedani, M.P.; Shekarchi, M. Comparative study of rosmarinic acid content in some plants of Labiatae family. Pharmacogn. Mag. 2012, 8, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Vamanu, E.; Nita, S. Antioxidant Capacity and the Correlation with Major Phenolic Compounds, Anthocyanin, and Tocopherol Content in Various Extracts from the Wild EdibleBoletus edulisMushroom. BioMed Res. Int. 2012, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vamanu, E.; Pelinescu, D.; Avram, I.; Nita, S. AnIn VitroEvaluation of Antioxidant and Colonic Microbial Profile Levels following Mushroom Consumption. BioMed Res. Int. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, X.; Chen, Q.; Luo, L.; Ma, M.; Xiao, B.; Zeng, L. Camellia sinensis and Litsea coreana Ameliorate Intestinal Inflammation and Modulate Gut Microbiota in Dextran Sulfate Sodium-Induced Colitis Mice. Mol. Nutr. Food Res. 2020, 64, e1900943. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L. Health Effects and Sources of Prebiotic Dietary Fiber. Curr. Dev. Nutr. 2018, 2, nzy005. [Google Scholar] [CrossRef] [Green Version]
- Diling, C.; Xin, Y.; Chaoqun, Z.; Jian, Y.; Xiaocui, T.; Jun, C.; Ou, S.; Yizhen, X. Extracts from Hericium erinaceus relieve inflammatory bowel disease by regulating immunity and gut microbiota. Oncotarget 2017, 8, 85838–85857. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Su, L.; Li, D.; Shuai, O.; Zhang, Y.; Liang, H.; Jiao, C.; Xu, Z.; Lai, Y.; Xie, Y. Antitumor Activity of Extract from the Sporoderm-Breaking Spore of Ganoderma lucidum: Restoration on Exhausted Cytotoxic T Cell With Gut Microbiota Remodeling. Front. Immunol. 2018, 9, 1765. [Google Scholar] [CrossRef]
- Vamanu, E.; Pelinescu, D. Effects of mushroom consumption on the microbiota of different target groups—Impact of polyphenolic composition and mitigation on the microbiome fingerprint. LWT Food Sci. Technol. 2017, 85, 262–268. [Google Scholar] [CrossRef]
- Mitsou, E.K.; Saxami, G.; Stamoulou, E.; Kerezoudi, E.; Terzi, E.; Koutrotsios, G.; Bekiaris, G.; Zervakis, G.I.; Mountzouris, K.C.; Pletsa, V.; et al. Effects of Rich in Β-Glucans Edible Mushrooms on Aging Gut Microbiota Characteristics: An In Vitro Study. Molecules 2020, 25, 2806. [Google Scholar] [CrossRef]
- Shimizu, T.; Mori, K.; Ouchi, K.; Kushida, M.; Tsuduki, T. Effects of Dietary Intake of Japanese Mushrooms on Visceral Fat Accumulation and Gut Microbiota in Mice. Nutrients 2018, 10, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Santamarina, A.; Mondragon, A.D.C.; Lamas, A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Animal-Origin Prebiotics Based on Chitin: An Alternative for the Future? A Critical Review. Foods 2020, 9, 782. [Google Scholar] [CrossRef]
- Hisano, M.; Bruschini, H.; Nicodemo, A.C.; Srougi, M. Cranberries and lower urinary tract infection prevention. Clinics 2012, 67, 661–667. [Google Scholar] [CrossRef]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Lull, C.; Wichers, H.; Savelkoul, H.F.J. Antiinflammatory and Immunomodulating Properties of Fungal Metabolites. Mediat. Inflamm. 2005, 2005, 63–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, V.; Ferrão, J.; Chaquisse, E.; Fernandes, T. Host-Microbial Gut Interactions and Mushroom Nutrition. J. Food Nutr. Res. 2018, 6, 576–583. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta Glucan: Supplement or Drug? From Laboratory to Clinical Trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verywell Health. Available online: www.verywellhealth.com (accessed on 8 October 2020).
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [Green Version]
- Conlon, M.A.; Bird, A.R. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef]
- Hills, J.R.D.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Ma, Q.; Chen, Y.; Lu, Y.; Wang, Y.; Jia, Y.; Zhang, M.; Chen, H. Structure characterization of soluble dietary fiber fractions from mushroom Lentinula edodes (Berk.) Pegler and the effects on fermentation and human gut microbiota in vitro. Food Res. Int. 2020, 129, 108870. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, Y.-T.; Pan, C.-H.; Chien, Y.-W.; Huang, H.-Y. Edible Mushrooms: Novel Medicinal Agents to Combat Metabolic Syndrome and Associated Diseases. Curr. Pharm. Des. 2020, 26, 4970–4981. [Google Scholar] [CrossRef]
- Giavasis, I. Polysaccharides from Medicinal Mushrooms for Potential Use as Nutraceuticals, Polysaccharides Natural Fibers in Food and Nutrition, 1st ed.; Benkeblia, N., Ed.; CRC Press: Boca Raton, FL, USA, 2014; Chapter 8; pp. 171–206. [Google Scholar] [CrossRef]
- Tu, J.; Brennan, M.; Brennan, C. An insight into the mechanism of interactions between mushroom polysaccharides and starch. Curr. Opin. Food Sci. 2021, 37, 17–25. [Google Scholar] [CrossRef]
- Yin, C.; Noratto, G.D.; Fan, X.; Chen, Z.; Yao, F.; Shi, D.; Gao, H. The Impact of Mushroom Polysaccharides on Gut Microbiota and Its Beneficial Effects to Host: A Review. Carbohydr. Polym. 2020, 250, 116942. [Google Scholar] [CrossRef]
- El Kaoutari, A.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Genet. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Xue, Z.; Chen, Y.; Jia, Y.; Wang, Y.; Lu, Y.; Chen, H.; Zhang, M. Structure, thermal and rheological properties of different soluble dietary fiber fractions from mushroom Lentinula edodes (Berk.) Pegler residues. Food Hydrocoll. 2019, 95, 10–18. [Google Scholar] [CrossRef]
- Kodio, A.; Menu, E.; Ranque, S. Eukaryotic and Prokaryotic Microbiota Interactions. Microorganisms 2020, 8, 2018. [Google Scholar] [CrossRef]
- Zhao, R.; Yang, W.; Pei, F.; Zhao, L.; Hu, Q. In vitro fermentation of six kinds of edible mushrooms and its effects on fecal microbiota composition. LWT Food Sci. Technol. 2018, 96, 627–635. [Google Scholar] [CrossRef]
- Fuke, N.; Nagata, N.; Suganuma, H.; Ota, T. Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors. Nutrients 2019, 11, 2277. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Y.; Ke, Y.; Li, C.; Zhang, Z.; Wu, Y.; Hu, B.; Liu, A.; Luo, Q.; Wu, W. In vitro saliva-gastrointestinal digestion and fecal fermentation of Oudemansiella radicata polysaccharides reveal its digestion profile and effect on the modulation of the gut microbiota. Carbohydr. Polym. 2021, 251, 117041. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Su, A.; Ma, G.; Tao, T.; Fang, D.; Zhao, L.; Hu, Q. Comparison of bioactive constituents and effects on gut microbiota by in vitro fermentation between Ophicordyceps sinensis and Cordyceps militaris. J. Funct. Foods 2020, 68, 103901. [Google Scholar] [CrossRef]
- Yu, J.; Xiang, H.; Xie, Q. The difference of regulatory effect of two Inonotus obliquus extracts on high-fat diet mice in relation to the fatty acid elongation function of gut microbiota. Food Sci. Nutr. 2021, 9, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, C.; Li, J.; Li, T.; Zhang, Y.; Liang, Y.; Mei, Y. Phellinus linteus polysaccharide extract improves insulin resistance by regulating gut microbiota composition. FASEB J. 2019, 34, 1065–1078. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.; Zhu, H.; Hu, Q.; Liu, Y.; Zhao, S.; Peng, N.; Liang, Y. A novel polysaccharide from mycelia of cultured Phellinus linteus displays antitumor activity through apoptosis. Carbohydr. Polym. 2015, 124, 90–97. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, L.; Wang, W.; Qiu, W.; Liu, L.; Ning, A.; Cao, J.; Huang, M.; Zhong, M. Polysaccharides isolated from Cordyceps Sinensis contribute to the progression of NASH by modifying the gut microbiota in mice fed a high-fat diet. PLoS ONE 2020, 15, e0232972. [Google Scholar] [CrossRef]
- Nakahara, D.; Nan, C.; Mori, K.; Hanayama, M.; Kikuchi, H.; Hirai, S.; Egashira, Y. Effect of mushroom polysaccharides from Pleurotus eryngii on obesity and gut microbiota in mice fed a high-fat diet. Eur. J. Nutr. 2019, 59, 3231–3244. [Google Scholar] [CrossRef]
- Chen, L.; Tai, W.C.; Hsiao, W.W. Dietary saponins from four popular herbal tea exert prebiotic-like effects on gut microbiota in C57BL/6 mice. J. Funct. Foods 2015, 17, 892–902. [Google Scholar] [CrossRef]
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Zhang, Z.; Yang, C.; Hu, M.; Sun, Y.; He, C.-N.; et al. The chemopreventive effects of Huangqin-tea against AOM-induced preneoplastic colonic aberrant crypt foci in rats and omics analysis. Food Funct. 2020, 11, 9634–9650. [Google Scholar] [CrossRef] [PubMed]
- Bond, T.; Derbyshire, E. Tea Compounds and the Gut Microbiome: Findings from Trials and Mechanistic Studies. Nutrients 2019, 11, 2364. [Google Scholar] [CrossRef] [Green Version]
- Dabulici, C.M.; Sârbu, I.; Vamanu, E. The Bioactive Potential of Functional Products and Bioavailability of Phenolic Compounds. Foods 2020, 9, 953. [Google Scholar] [CrossRef]
- Reyes, C.M.; Cornelis, M.C. Caffeine in the Diet: Country-Level Consumption and Guidelines. Nutrients 2018, 10, 1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Chen, Z.; Wang, D.; Chen, G.; Sun, X.; He, Q.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; et al. Effects of Fermented Herbal Tea Residues on the Intestinal Microbiota Characteristics of Holstein Heifers Under Heat Stress. Front. Microbiol. 2020, 11, 1014. [Google Scholar] [CrossRef]
- McKay, D.L.; Blumberg, J.B. A review of the bioactivity and potential health benefits of peppermint tea (Mentha piperita L.). Phytother. Res. 2006, 20, 619–633. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.L.; Blumberg, J.B. A Review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.). Phytother. Res. 2006, 20, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Gruenwald, J.; Freder, J.; Armbruester, N. Cinnamon and Health. Crit. Rev. Food Sci. Nutr. 2010, 50, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Abbasabadi, Z.; Ardekani, M.R.S.; Rahimi, R.; Farzaei, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 33, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Burcuș, A.; Vamanu, E.; Sarbu, I.; Petre, A. Antioxidant, Anti-Inflammatory, and Antibacterial Potential of Different Drinks Based on Matcha Tea. IOP Conf. Ser. Mater. Sci. Eng. 2018, 374, 012072. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The Two-Way Polyphenols-Microbiota Interactions and Their Effects on Obesity and Related Metabolic Diseases. Front. Nutr. 2019, 6, 188. [Google Scholar] [CrossRef] [Green Version]
- Dong, R.; Liu, S.; Xie, J.; Chen, Y.; Zheng, Y.; Zhang, X.; Zhao, E.; Wang, Z.; Xu, H.; Yu, Q. The recovery, catabolism and potential bioactivity of polyphenols from carrot subjected to in vitro simulated digestion and colonic fermentation. Food Res. Int. 2021, 143, 110263. [Google Scholar] [CrossRef]
- Song, C.; Xie, C.; Zhou, Z.; Yu, S.; Fang, N. Antidiabetic Effect of an Active Components Group fromIlex kudingchaand Its Chemical Composition. Evid. Based Complement. Altern. Med. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Xie, M.; Dai, Z.; Wan, P.; Ye, H.; Zeng, X.; Sun, Y. Kudingcha and Fuzhuan Brick Tea Prevent Obesity and Modulate Gut Microbiota in High-Fat Diet Fed Mice. Mol. Nutr. Food Res. 2018, 62, e1700485. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Y.; Xiao, Y.; Zhong, K.; Bai, J.-R.; Wu, Y.-P.; Zhang, J.-Q.; Gao, H. Characteristics and chemical compositions of Pingwu Fuzhuan brick-tea, a distinctive post-fermentation tea in Sichuan province of China. Int. J. Food Prop. 2019, 22, 878–889. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, J.M.; Munekata, P.E.S. Phenolic compounds of green tea: Health benefits and technological application in food. Asian Pac. J. Trop. Biomed. 2016, 6, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Jung, E.S.; Park, J.I.; Park, H.; Holzapfel, W.; Hwang, J.S.; Lee, C.H. Seven-day Green Tea Supplementation Revamps Gut Microbiome and Caecum/Skin Metabolome in Mice from Stress. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Zheng, J.; Zou, X.; Ye, C.; Xia, H.; Yang, M.; Gao, Q.; Yang, Q.; Liu, H. Ligustrum robustum (Roxb.) blume extract modulates gut microbiota and prevents metabolic syndrome in high-fat diet-fed mice. J. Ethnopharmacol. 2021, 268, 113695. [Google Scholar] [CrossRef]
- Popowski, D.; Czerwińska, M.E.; Kruk, A.; Pawłowska, K.A.; Zentek, J.; Melzig, M.F.; Piwowarski, J.P.; Granica, S. Gut microbiota metabolism and the permeability of natural products contained in infusions from herb of European goldenrod Solidago virgaurea L. J. Ethnopharmacol. 2021, 273, 113924. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hao, Y.; Gao, B.; Geng, P.; Huang, H.; Yu, L.; Choe, U.; Liu, J.; Sun, J.; Chen, P.; et al. Chemical profile and in vitro gut microbiota modulatory, anti-inflammatory and free radical scavenging properties of chrysanthemum morifolium cv. Fubaiju. J. Funct. Foods 2019, 58, 114–122. [Google Scholar] [CrossRef]
- Song, D.; Ho, C.-T.; Zhang, X.; Wu, Z.; Cao, J. Modulatory effect of Cyclocarya paliurus flavonoids on the intestinal microbiota and liver clock genes of circadian rhythm disorder mice model. Food Res. Int. 2020, 138, 109769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, H.; Zhao, H.; Geng, Y.; Ren, Y.; Guo, L.; Shi, J.; Xu, Z. Edgeworthia gardneri (Wall.) Meisn. water extract improves diabetes and modulates gut microbiota. J. Ethnopharmacol. 2019, 239, 111854. [Google Scholar] [CrossRef]
- Gu, J.-F.; Su, S.-L.; Guo, J.-M.; Zhu, Y.; Zhao, M.; Duan, J.-A. The aerial parts of Salvia miltiorrhiza Bge. strengthen intestinal barrier and modulate gut microbiota imbalance in streptozocin-induced diabetic mice. J. Funct. Foods 2017, 36, 362–374. [Google Scholar] [CrossRef]
- Jin, D.-X.; He, J.-F.; Luo, X.-G.; Zhang, T.-C. Hypoglycemic effect of Hypericum attenuatum Choisy extracts on type 2 diabetes by regulating glucolipid metabolism and modulating gut microbiota. J. Funct. Foods 2019, 52, 479–491. [Google Scholar] [CrossRef]
- Henning, S.M.; Yang, J.; Hsu, M.; Lee, R.-P.; Grojean, E.M.; Ly, A.; Tseng, C.-H.; Heber, D.; Li, Z. Decaffeinated green and black tea polyphenols decrease weight gain and alter microbiome populations and function in diet-induced obese mice. Eur. J. Nutr. 2018, 57, 2759–2769. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-S.; Xu, J.; Zhu, H.; Wu, J.; Xu, J.-D.; Yan, R.; Li, X.-Y.; Liu, H.-H.; Duan, S.-M.; Wang, Z.; et al. Gut microbiota-involved mechanisms in enhancing systemic exposure of ginsenosides by coexisting polysaccharides in ginseng decoction. Sci. Rep. 2016, 6, 22474. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Liu, J.; Lv, Y.-J.; Jiang, Y.-L.; Pan, J.-X.; Zhu, Y.-J.; Huang, M.-G.; Zhang, S.-K. Changes in Intestinal Microbiota of Type 2 Diabetes in Mice in Response to Dietary Supplementation with Instant Tea or Matcha. Can. J. Diabetes 2020, 44, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Foster, M.T.; Gentile, C.L.; Cox-York, K.; Wei, Y.; Wang, D.; Estrada, A.L.; Reese, L.; Miller, T.; Pagliassotti, M.J.; Weir, T.L. Fuzhuan tea consumption imparts hepatoprotective effects and alters intestinal microbiota in high saturated fat diet-fed rats. Mol. Nutr. Food Res. 2016, 60, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ni, J.; Li, H. Effect of green tea and mulberry leaf powders on the gut microbiota of chicken. BMC Vet. Res. 2019, 15, 77. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Cai, R.; Tan, Y.; Wu, X.; Wen, Q.; Liu, Z.; Ouyang, S.-H.; Yin, Z.; Yang, H. Preventive consumption of green tea modifies the gut microbiota and provides persistent protection from high-fat diet-induced obesity. J. Funct. Foods 2020, 64, 103621. [Google Scholar] [CrossRef]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial Properties of Green Tea Catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef] [Green Version]
- Takagaki, A.; Nanjo, F. Catabolism of (+)-Catechin and (−)-Epicatechin by Rat Intestinal Microbiota. J. Agric. Food Chem. 2013, 61, 4927–4935. [Google Scholar] [CrossRef]
- Vandeponseele, A.; Draye, M.; Piot, C.; Chatel, G. Study of Influential Parameters of the Caffeine Extraction from Spent Coffee Grounds: From Brewing Coffee Method to the Waste Treatment Conditions. Clean Technol. 2021, 3, 19. [Google Scholar] [CrossRef]
- Shin, B.K.; Kwon, S.W.; Park, J.H. Chemical diversity of ginseng saponins from Panax ginseng. J. Ginseng Res. 2015, 39, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Luo, L.; Luo, Y.; Zhang, J.; Wang, X.; Sun, K.; Zeng, L. Prebiotic Properties of Green and Dark Tea Contribute to Protective Effects in Chemical-Induced Colitis in Mice: A Fecal Microbiota Transplantation Study. J. Agric. Food Chem. 2020, 68, 6368–6380. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D. The Gut Microbiome and Its Role in Obesity. Nutr. Today 2016, 51, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-S.; Kim, Y. Chrysanthemum morifolium Flower Extract Inhibits Adipogenesis of 3T3-L1 Cells via AMPK/SIRT1 Pathway Activation. Nutrients 2020, 12, 2726. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Zhang, J.; Wang, L.; Li, F.; Li, Z.; Xiang, W.; Bie, S.; Wang, C.; Li, Z. Ten New Dammarane-Type Saponins with Hypolipidemia Activity from a Functional Herbal Tea—Gynostemma pentaphyllum. Molecules 2020, 25, 3737. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Postler, T.; Ghosh, S. Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System. Cell Metab. 2017, 26, 110–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front. Nutr. 2021, 8, 586564. [Google Scholar] [CrossRef]
- Li, M.; Chen, W.-D.; Wang, Y.-D. The roles of the gut microbiota–miRNA interaction in the host pathophysiology. Mol. Med. 2020, 26, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Musolino, C.; Tonacci, A.; Pioggia, G.; Gangemi, S. Interactions between the MicroRNAs and Microbiota in Cancer Development: Roles and Therapeutic Opportunities. Cancers 2020, 12, 805. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Su, M.; Qiu, Y.; Chen, M.; Liu, Y.; Zhao, A.; Jia, W. Metabolic profiling using combined GC-MS and LC-MS provides a systems understanding of aristolochic acid-induced nephrotoxicity in rat. FEBS Lett. 2007, 581, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, X.; Bao, Q.; Di, S.; Zhao, Y.; Zhao, S.; Zhang, H.; Lian, F.; Tong, X. The interaction between the gut Microbiota and herbal medicines. Biomed. Pharmacother. 2019, 118, 109252. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Long, Y.; Ji, Z.; Gao, J.; Fu, T.; Yan, M.; Zhang, L.; Su, H.; Zhang, W.; Wen, X.; et al. Green Tea Liquid Consumption Alters the Human Intestinal and Oral Microbiome. Mol. Nutr. Food Res. 2018, 62, e1800178. [Google Scholar] [CrossRef] [Green Version]
- MiRBase: The microRNA Database. Available online: http://www.mirbase.org (accessed on 5 March 2021).
- Li, Z.; Xu, R.; Li, N. MicroRNAs from plants to animals, do they define a new messenger for communication? Nutr. Metab. 2018, 15, 1–21. [Google Scholar] [CrossRef]
- Zhao, Y.; Cong, L.; Lukiw, W.J. Plant and Animal microRNAs (miRNAs) and Their Potential for Inter-kingdom Communication. Cell. Mol. Neurobiol. 2017, 38, 133–140. [Google Scholar] [CrossRef]
- Bi, K.; Zhang, X.; Chen, W.; Diao, H. MicroRNAs Regulate Intestinal Immunity and Gut Microbiota for Gastrointestinal Health: A Comprehensive Review. Genes 2020, 11, 1075. [Google Scholar] [CrossRef]
- Alnuqaydan, A.M. Targeting micro-RNAs by natural products: A novel future therapeutic strategy to combat cancer. Am. J. Transl. Res 2020, 12, 3531–3556. [Google Scholar]
- Zhang, B.; Tian, L.; Xie, J.; Chen, G.; Wang, F. Targeting miRNAs by natural products: A new way for cancer therapy. Biomed. Pharmacother. 2020, 130, 110546. [Google Scholar] [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runtsch, M.C.; Round, J.L.; O’Connell, R.M. MicroRNAs and the regulation of intestinal homeostasis. Front. Genet. 2014, 5, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinler, J.K.; Oezguen, N.; Runge, J.K.; Luna, R.A.; Karri, V.; Yang, J.; Hirschi, K.D. Dietary impact of a plant-derived microRNA on the gut microbiome. ExRNA 2020, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Paz, H.A.; Sadri, M.; Cui, J.; Kachman, S.D.; Fernando, S.C.; Zempleni, J. Dietary bovine milk exosomes elicit changes in bacterial communities in C57BL/6 mice. Am. J. Physiol. Liver Physiol. 2019, 317, G618–G624. [Google Scholar] [CrossRef]
- Xie, W.; Weng, A.; Melzig, M.F. MicroRNAs as New Bioactive Components in Medicinal Plants. Planta Med. 2016, 82, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Cheng, X.; Zhang, T.; Liu, L.; Nie, Z.; Sheng, Q. The identification of microRNAs in Ganoderma lingzhi sporocarp. Mycoscience 2016, 57, 271–278. [Google Scholar] [CrossRef]
- Xie, W.; Melzig, M.F. The Stability of Medicinal Plant microRNAs in the Herb Preparation Process. Molecules 2018, 23, 919. [Google Scholar] [CrossRef] [Green Version]
- Petre, M.; Teodorescu, A.; Pătrulescu, F. Biotechnology of submerged fermentation to produce nutritive mycelial bio-mass through controlled cultivation of edible and medicinal mushrooms. Sci. Bull. Ser. F. Biotechnol. 2012, 16, 89–93. [Google Scholar]


{kind=link}
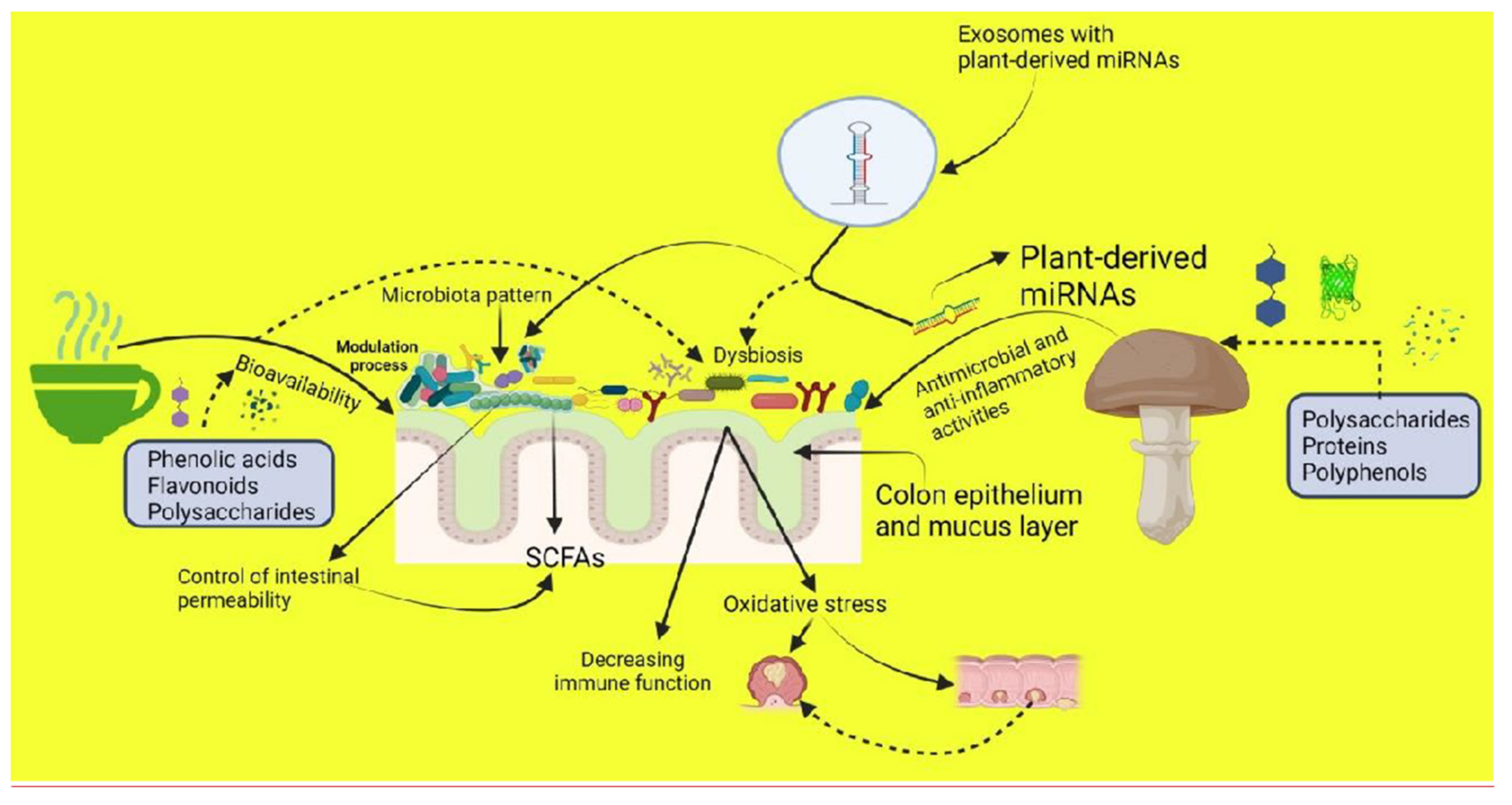{kind=link}
| Mushrooms | Gut Bacteria Increased | Gut Bacteria Depleted | Human/ Animal | References |
|---|---|---|---|---|
| Hericium erinaceus (Lion’s mane mushroom) | Bifidobacterium, Coprococcus, Desulfovibrio, Lactobacillus, Parabacteroides, Prevotella | Corynebacterium, Dorea, Roseburia, Ruminococcus, Staphylococcus, Sutterella | Animal (rat) | [25] |
| Ganoderma lucidum (Reishi) | Firmicutes, Proteobacteria (Helicobacter), Rikenella | Acinetobacter, Actinobacteria (Arthrobacter, Corynebacterium), Bacteroidetes (Bacteroides, Parabacteroides, Prevotella), Blautia, Brevundimonas, Clostridium, Coprobacillus, Cyanobacteria, Facklamia, Jeotgalicoccus, Sporosarcina, Staphylococcus, Streptococcus | Animal (mice) | [26] |
| Boletus edulis, Boletus pinophilus, Boletus aureus (Porcini), Armillaria mellea (Honey fungus), Lactarius piperatus (blancaccio), Pleurotus eryngii (King oyster) | Bifidobacterium and Lactobacillus genera | - | Human | [27] |
| Cyclocybe cylindracea (poplar mushroom), Hericium erinaceus, Pleurotus eryngii, Pleurotus ostreatus (Oyster mushroom) | Bifidobacterium spp. Faecalibacterium prausnitzii (Ruminococcaceae), Eubacterium rectale/Roseburia spp. | - | In vitro study using human faeces | [28] |
| Flammulina velutipes (Enoki), Hypsizygus marmoreus, (White beech mushroom), Lentinula edodes (Shiitake), Grifola frondosa, (Maitake) Pleurotus eryngii | Allobaculum, Bifidobacterium, Ruminococcus, Lactobacillus, Lactococcus, Streptococcus | Bacteroides, Prevotella, Mucispirillum, Dorea, Roseburia, Anaerotruncus, Oscillospira, Escherichia and Akkermansia | Animal (mice) | [29] |
| Mushrooms (Common Name) | Bioactive Compounds | Type of Study | Gut Bacteria Effects | References |
|---|---|---|---|---|
| Lentinula edodes (Shiitake) | Lentinula edodes soluble dietary fiber fractions (LESDF): LESDF-1: →6)-β-D-Glcp-(1→,→4)-β-D-Glcp-(1→,→5)-β-D-Arap-(1→,→4)-β-D-Xylp-(1→,→4)-α-D-Manp,→3)-α-L-Rhap-(1→,→6)-β-D-Galp-(1→ LESDF-2: →6)-β-D-Glcp-1→,→4)-α-D-Glcp-(1→,β-DGlcp-(1→,→5)-β-D-Arap-(1→,→2)-α-L-Rhap-(1→,→3,6)-α-D-Manp-(1→,→6)-β-D-Galp-(1→,→4)-β-D-Xylp-(1→ LESDF-3: β-D-Arap-(1→,→3)-α-D-Galp-(1→,→3,6)-α-D-Manp-(1→, →4)- β-D-Xylp-(1→, and →2,4)-α-D-Glcp-(1→ | Human gut microbiota in vitro study | LEDS-2 increase microbial communities LEDS-3 causes an increase in the abundance of Parasutterella, Bacteroides, Parabacteroides and Lachnospira | [42,49,50] |
| Auricularia auricula-judae (wood ear) Flammulina velutipes (velvet shank) | Mushrooms dried powder after in vitro digestion with α-amylase, pepsin, and pancreatin. The main bioactive compounds are carbohydrates and proteins. | Human gut microbiota in vitro study | They led to an increase in the abundance of groups of Actinobacteria, Bacteroidetes, Proteobacteria and inhibited the growth of Fusobacteria and Firmicutes | [51] |
| Lentinus edodes Pleurotus eryngii (King Oyster) | Promoted the growth of Bacteroidetes, Actinobacteria and inhibited the development of Proteobacteria, Fusobacteria, and Firmicutes | |||
| Pleurotus osteratus (Oyster mushroom) | Promoted the growth of Actinobacteria, Bacteroidetes, and Fusobacteria, inhibited the growth of Proteobacteria and Firmicutes. | |||
| Agaricus bispours (champigno n) | Positively influences Actinobacteria, Fusobacteria, and Firmicutes and inhibit the growth of Bacteroidetes and Proteobacteria | |||
| Hericium erinaceus (lion’s mane mushroom) | Polysaccharides, alcoholic extracts, and whole extracts alcoholic extracts, and whole extracts | Rats with inflammatory bowel disease | Reduce the amount of lipopolysaccharide toxins, increase the abundance of Bifidobacterium; | [25,52] |
| Oudemansiella radicata (Rooted Collybia) | Polysaccharide extract | Human gut microbiota in vitro study | Reduce the Firmicutes/Bacteroidetes ratio Increase Bacteroides abundance | [53] |
| Ophicordyceps sinensis (Rooted Collybia) | Mushrooms dried powder after in vitro digestion with α-amylase, pepsin, and pancreatin | Human gut microbiota in vitro study | Increase abundance of Bifidobacteriales, Selenomonadales | [54] |
| Cordyceps militaris (Chinese caterpillar fungus) | Increase the relative abundance Bacteroidales. Implicitly decreasing the ratio of Firmicutes/Bacteroidetes ratio | |||
| Inonotus obliquus (chaga) | Polysaccharides Ethanolic extract | High-fat diet mice (HFD-mice) | Increase Akkermansia abundance and fatty acid elongation | [55] |
| Phellinus linteus (black hoof mushroom) | Polysaccharide total extract (two fractions were characterized, PLPS-1: α-D-glucose (1→4)-α-D-glucose (1→6) units and PLPS-2: α-(1→3)- D-glucose and α-(1→6)-D-glucose) | Sprague Dawley rats with Type 2 diabetes | Causes an increase in the abundance of Lachnospiraceae-NK4A1 36, Blautia, Ruminiclostridium-9, Eubacterium xylanophilum, Anaerotruncus, Oscillibacter Lachnospiraceae-UCG-00 6, Roseburia, Prevotella and improves microbial balance | [56,57] |
| Cordycepssinensis (Cordyceps mushroom) | Polysaccharide fraction | HFD mice | The relative abundances of Actinobacteria (in particular Olsenella bacteria) and Acidobacterias were increased and those of Bacteroidetes decreased. At the genus/cluster level, decreases of Barnesiella, Prevotellaceae and the Lachnospiraceae incertae sedis and increases of Christensenella, Clostridium_XVIII cluster and Pseudomonas | [58] |
| Pleurotus eryngii | Soluble polysaccharide fraction | HFD mice | Supplementation causes changes only at the genus level, an increase of Anaerostipes, Clostridium, Lactococcus, and a decrease of Roseburia and Lactobacillus | [59] |
| Herbals | Beneficial Effects | Reference |
|---|---|---|
| Matricaria recutita (Chamomile) | Anti-inflammatory, antispasmodic, antioxidative activity, antiplatelet activity | [66] |
| Mentha piperita (Peppermint) | Antimicrobial and antiviral activity, analgesic and anesthetic effects, immunomodulatory activity | [67] |
| Cinnamomum zeylanicum (Cinnamon) | Antimicrobials activity, anti-inflammatory, antiviral, antioxidant, antitumoral activity, Cholesterol- and Lipid-Lowering Properties | [68] |
| Petroselinum crispum (Parsley) | Antioxidant, hepatoprotective, anti-diabetic, analgesic, immunosuppressant, anti-platelet, gastroprotective, cytoprotective, laxative, estrogenic, diuretic, hypotensive, antibacterial, and antifungal activities | [69] |
| Camellia sinensis (Macha tea) | Antimicrobial activity, anti-inflammatory | [70] |
| Tea | Bioactive Compounds | Type of Study | Gut Bacteria Effects | References |
|---|---|---|---|---|
| Kudingcha (KDC) from Ilex latifolia Thun and Ilex kudingcha C.J. Tseng (large-leaved Kudingcha) | Neochlorogenic acid, chlorogenic acid, cryptochlorogenic acid, dicaffeoylquinic acids isomers, quercetin with different glycosides triterpenoid saponins, polysaccharides, monosaccharides, proteins, simple organic acid. | HFD mice | Administration of KDC led to a reduction in abundance of Erysipelotrichaceae. | [73,74,75] |
| Fuzhuanbrick Tea post-ferment ed tea (dark tea leaves of Camellia sinensis var. sinensis and C. sinensis var. assamica) (border-selling tea or border-tea) | Gallic acid, catechins, free amino acids, alkaloids and volatile components. | Administration of the tea reduces the Firmicutes/Bacteroidetes ratio and has led to the increased of relative abundance in Bifidobacteriaceae | ||
| KDC and FBT | Led to an decreased of Clostridium, Bilophila, Oscillibacter, Lactonifactor, Eisenbergiella, Olsenella, Leuconostoc, Pseudoflavonifractor and Streptococcus. | |||
| Green tea (Camellia sinensis) | Standardized green tea extract: catechins (49.9%), including epigallocatechin (9.7%), epicatechin (5.4%), epigallocatechin-gallate (28.4%), and epicatechin-gallate (6.4%), as well as caffeine (4.5%) and theanine (0.4%). Other possible biologically active compounds: gallic acid, p-coumaric acid and quinic acid derivatives, caffeoylquinic acid isomers, and caffeoyl, kaempferol 3-O-p-coumaroylglucoside and kaempferol 3-O-p-coumaroyldirhamnosylhexosi de | Mice under UV stress | 7-day supplementation of green tea extract Firmicutes/Bacteroidetes ratio and increased levels of Lactobacillus spp. and Bifidobacteriumspp. | [76,77] |
| Ligustrum robustum (Roxb.) Blume (bora-bora, Ceylon privét, privet, tree privet, troene) | Ligistrum robustum ethanol extract (LRE)—glycosides extract. Ligupurpuroside, acteoside, isoacteoside, ligupurpuroside A, ligupurpuroside B, ligupurpuroside C, ligupurpuroside D, and osmanthuside B | HFD mice | After 16 weeks of LRE administration, the ratio of Firmicutes/Bacteroidetes ratio increased. LRC contributed to growth stimulation of belonging of Streptococcus, Lactobacillus, and Eubacterium coprostanoligenes groups and a Coriobacteriaceae_U CG-002, and Lachnospiraceae groups. | [78] |
| Solidago virgaurea L. (European goldenrod, Woundwort) | Solidago v. Infusion extract Mainly caffeoylquinic acid derivatives (caffeic acid, p-coumaric acid, chlorogenic acid, neochlorogenic acid, cryptochlorogenic acid) flavonoids (quercetin rhamnohexoside, rutin, isoquercetin, kaempferol) and some phenylpropanoids. | Human and swine gut microbiota in vitro study | In human and swine cultures gut microbiota takes place hydrolysis of caffeoylquinic acid derivatives and deglycosylation of flavonoids. | [79] |
| Chrysanthem um morifolium (florist’s daisy and hardy garden mum or juhua) | Hot-water extract. Chlorogenic acid, tuberonic acid glucoside, diglucosylapigenin isomer, naringenin-6,8-di-C-glucoside Isookanin-7-O-β-diglucopyranoside, Quercetin-3-O-galactoside, diglucosyapigenin isomer, luteolin-7-O-glucuronide, luteolin-7-O-glucoside, dicaffeoylquinic acid isomer, apigenin-7-O-rutinoside, apigenin-7-O-glucoside, Kaempferol-3-O-acetyl-glucoside, diosmetin 7-O-rutinoside, diosmetin 7-glucuronide, acacetin-7-O-6″-malonylgactoside, apigenin 7-O-acetylglucoside isomer, apigenin 7-O-acetylglucoside isomer, Ombuin-3β-rutinoside, luteolin, apigenin 7-O-acetylglucoside isomer acacetin-7-O-glucuronide, acacetin, apigenina, diosmetin, eupatorin, casticina. | Regular chow diet fed C57BL/6J mice study | Hot water extract administration has shaped gut microbiota by increasing Bacteroidetes, Firmicutes (Prevotella) Bifidobacterium 6J mice with gut Colonized by microbiota coming from healthy human volunteers. | [80] |
| Cyclocarya paliurus (Wheel Wingnut; sweet tea tree) | Water extract of C. paliurus leaves. Cyclocarya paliurus flavonoids: Kaempferol-3-O-β-glucuronide, kaempferol-3-O-α-L-rhamnopyranoside, isoquercitrin, quercetin | Adult male C57BL | The administration of CPF led to an increase of microbial diversity, reduction in the relative abundance of Faecalibacterium, Mitsuokella, Ruminococcus, Desulfovibrio and Megamonas. | [81] |
| Edgeworthia gardneri (papertree, paperbush; Argelee) | Water extract, phloroglucinol, swainonine, trigonelline, coumalic acid, Coumarin, scopolamine, 7,8-Dihydroxy-4-methylcoumarin, chlorogenic acid, berberine, psoralen, apigenin, caffeic acid, γ-Terpinene, rutin, 4-methylumbelliferone, scopoletin, kaempferol-3-O-rutinoside, α-Pinene, daidzein, bergapten, glycitein, cytosine, α-Linolenic acid, ferulic acid, palmitoleic acid, linoleic acid, stearic acid, trans-vaccenic acid, arachidonic acid. | HFD mice | The microbial diversity was improved. The extract decreases the number of Proteobacteria and Deferribacteres and reverses the levels of Clostridiales, Lachnospiraceae,S24–7, Rikenellaceae, and Dorea in diabetic mice. | [82] |
| Salvia miltiorrhiza Bge (red sage or Danshen) | Ethanolic extract, Danshensu, protocatechualdehyden, caffeic acid, rutin, isoquercitrin, astragalin, rosmarinic acid, lithospermic acid, salvianolic acid B, salvianolic acid A, and salvianolic acid C | C57BL/6J diabetic mice | Decrease of Proteus hauseri and Helicobacter winghamensis abundance and growth stimulation of Anaerotruncus colihominis, Mucispirillum schaedleri, and Butyricimonas virosa. The extract increases the biodiversity and species of the gut microbiota and reduces the Firmicutes/Bacteroidetes ratio. | [83] |
| Hypericum attenuatum Choisy (St. John’s Wort) | Ethanolic extract, rutin, Quercetin-3-O-β-D-glucuronide | Male KM diabetic mice | The extract reverses dysbiosis induced by diabet, increases levels of Clostridiaceae, Erysipelotrichaceae and Lactobacillaceae | [84] |
| Decaffeinated green tea (GT) and black tea (BT) | Ethanolic extracts of green tea (GTP) and black tea (BTP), gallic acid, epigallocatechin gallate, epicatechin gallate, epigallocatechin, epicatechin | Male mouse C57BL | 6J mice (strain JAX 000664), low-fat/high sucros e diet (LF/HSD), HFD/HSD, HFD/HSD supplemented with GTP and BTP. HFD/HSD-GTP and BTP diets lead to a significant increase in the relative proportion of Parabacteroides, Bacteroides, and Prevotella and an increase of Roseburia, Bryantella, Lactococcus, Lactobacillus, Blautia, Anaerostipes, Shuttleworthia, and Acetitomaculum, Collinsella. GTP administration leads to an increase cell number of bacterial strains belonging to Clostridium and Coprococcus and a decrease of Turicibacter and Marvinbryantia. BTP consumption was increase in Oscillibacter, Anaerotruncus, and Pseudobutyrivibrio | [85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vamanu, E.; Dinu, L.D.; Pelinescu, D.R.; Gatea, F. Therapeutic Properties of Edible Mushrooms and Herbal Teas in Gut Microbiota Modulation. Microorganisms 2021, 9, 1262. https://doi.org/10.3390/microorganisms9061262
Vamanu E, Dinu LD, Pelinescu DR, Gatea F. Therapeutic Properties of Edible Mushrooms and Herbal Teas in Gut Microbiota Modulation. Microorganisms. 2021; 9(6):1262. https://doi.org/10.3390/microorganisms9061262
Chicago/Turabian StyleVamanu, Emanuel, Laura Dorina Dinu, Diana Roxana Pelinescu, and Florentina Gatea. 2021. "Therapeutic Properties of Edible Mushrooms and Herbal Teas in Gut Microbiota Modulation" Microorganisms 9, no. 6: 1262. https://doi.org/10.3390/microorganisms9061262