
First Report of blaNDM-1 Bearing IncX3 Plasmid in Clinically Isolated ST11 Klebsiella pneumoniae from Pakistan

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Samples Collection and Identification
2.2. Antibiotic Susceptibility Testing
2.3. Screening of Carbapenemases and ESBL
2.4. Detection of Antibiotic Resistance Genes
2.5. Multi Locus Sequence Typing (MLST)
2.6. Restriction Enzyme Analysis with Pulsed Field Gel Electrophoresis (REA-PFGE)
2.7. Transconjugation
2.8. Plasmids Analysis
3. Results
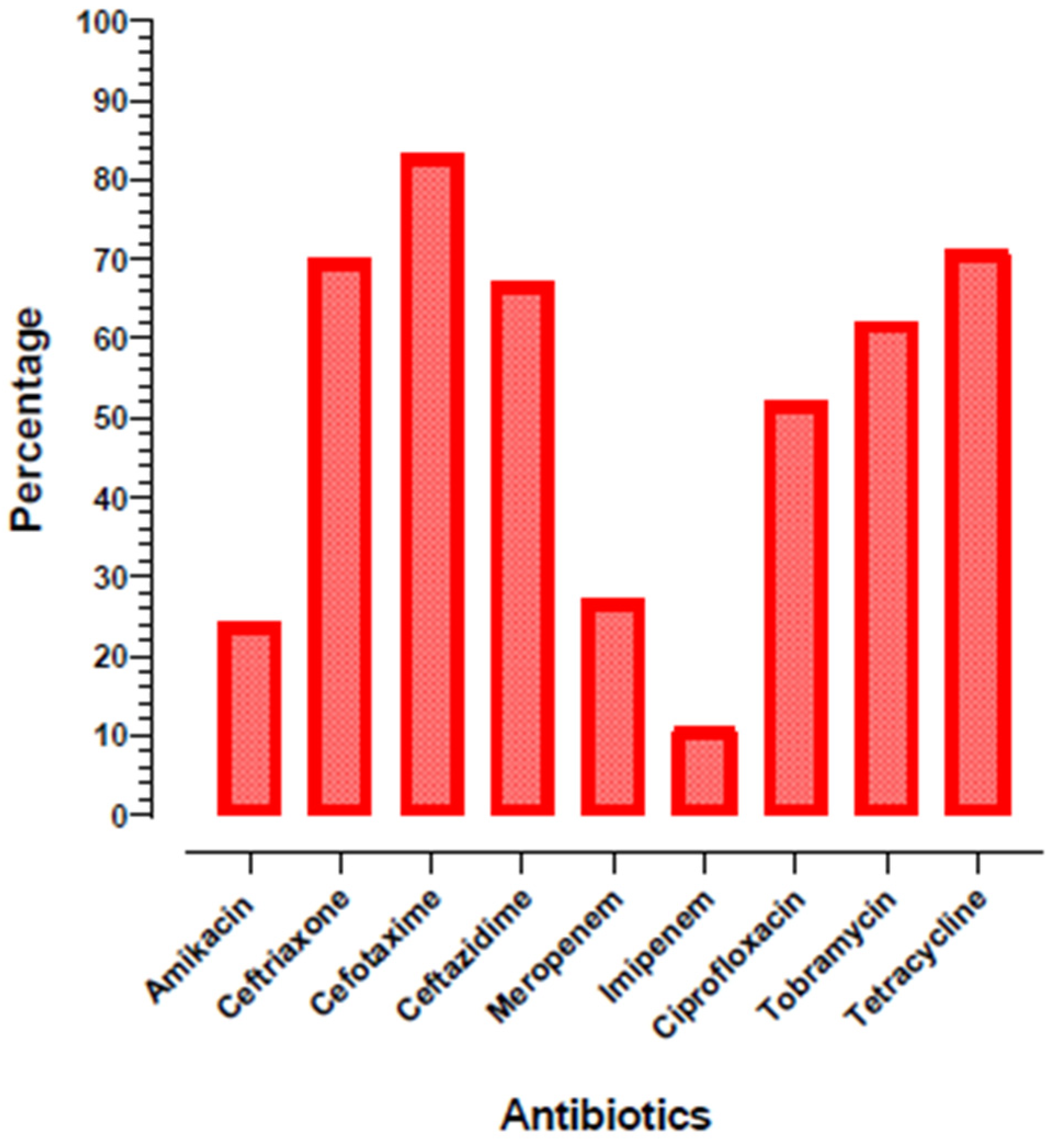
3.1. Bacterial Isolates and Antibiotic Sensitivity Profile
3.2. Carbapenemases Producing Isolates
3.3. Antibiotic-Resistant Genes
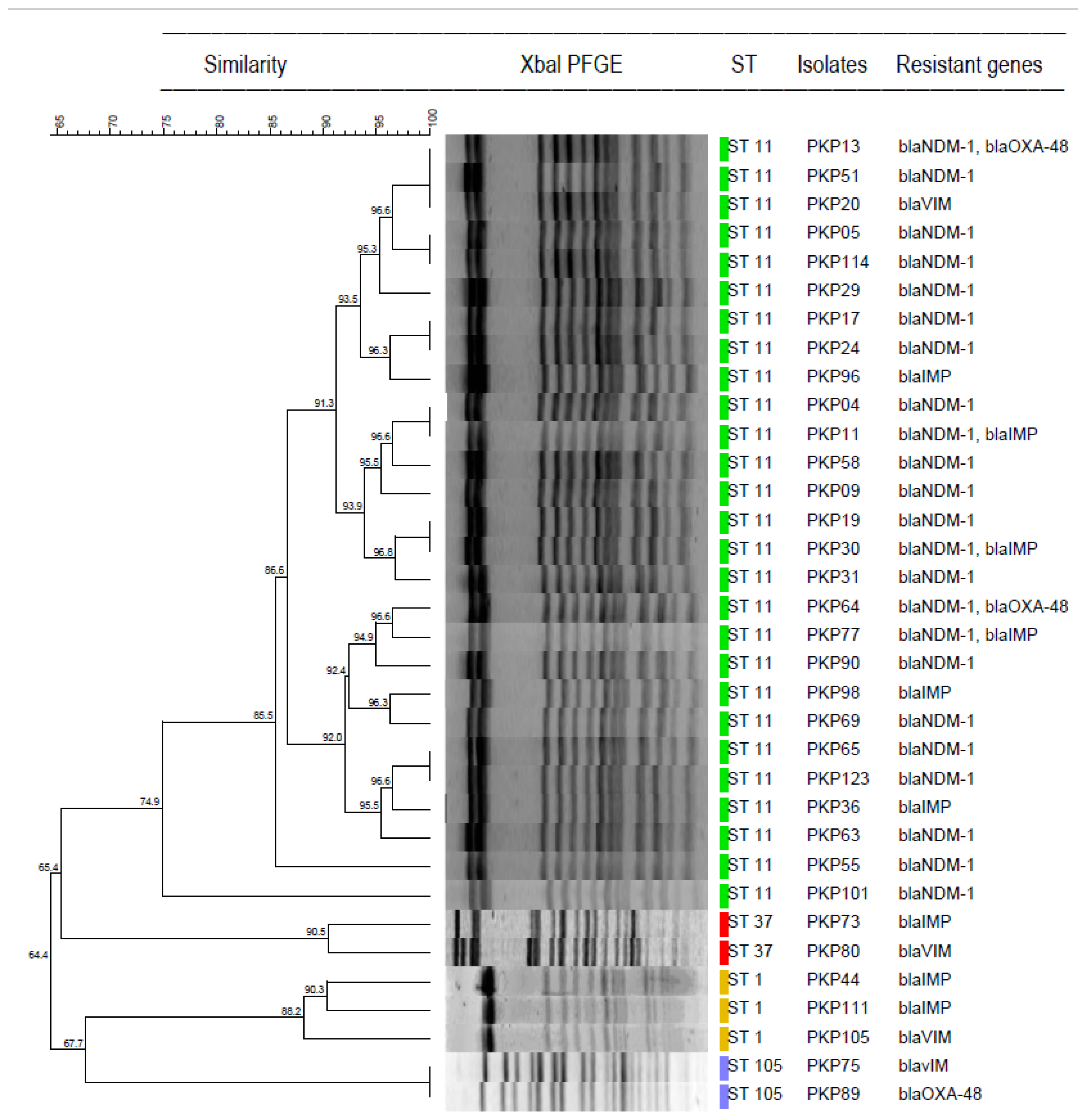
3.4. MLST
3.5. REA-PFGE
3.6. Transconjugation
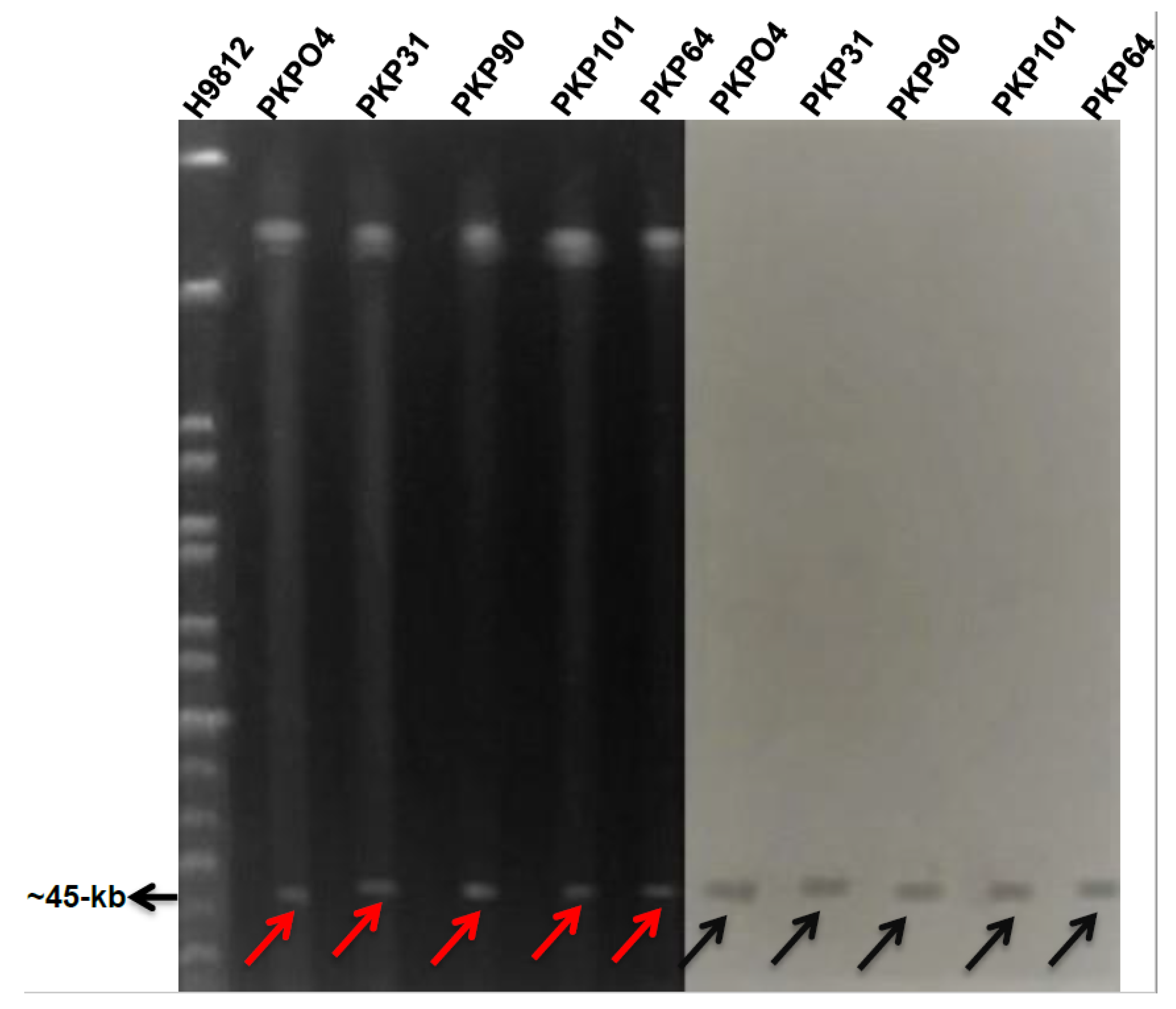
3.7. Plasmid Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dortet, L.; Poirel, L.; Nordmann, P. Worldwide dissemination of the NDM-type carbapenemases in Gram-negative bacteria. Biomed. Res. Int. 2014, 2014, 249856. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.D.; Sebastian, S.C.; Rekha, I.; Irshad, P.; Mudigonda, A.; Suresh, J.; Choudhary, S.; Tewari, R.; Ganaie, F.; Shome, B.R. Molecular detection of the New Delhi metallo-β-lactamase clinical variant with double mutation- V88L and M154L in Escherichia coli isolates from South India. Gene Rep. 2020, 21, 100880. [Google Scholar] [CrossRef]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-lactamase (NDM): A threat to public health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogaerts, P.; Verroken, A.; Jans, B.; Denis, O.; Glupczynski, Y. Global spread of New Delhi metallo-β-lactamase 1. Lancet Infect. Dis. 2010, 10, 831–832. [Google Scholar] [CrossRef]
- Rolain, J.M.; Parola, P.; Cornaglia, G. New Delhi metallo-beta-lactamase (NDM-1): Towards a new pandemia? Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2010, 16, 1699–1701. [Google Scholar] [CrossRef] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem Resistance: A Review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Baraniak, A.; Izdebski, R.; Fiett, J.; Gawryszewska, I.; Bojarska, K.; Herda, M.; Literacka, E.; Żabicka, D.; Tomczak, H.; Pewińska, N.; et al. NDM-producing Enterobacteriaceae in Poland, 2012–2014: Inter-regional outbreak of Klebsiella pneumoniae ST11 and sporadic cases. J. Antimicrob. Chemother. 2016, 71, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, E.; Irfan, S.; Sultan, B.A.; Nasir, A.; Hasan, R. Dissemination and spread of New Delhi Metallo-beta-lactamase-1 Superbugs in hospital settings. JPMA J. Pak. Med. Assoc. 2016, 66, 999–1004. [Google Scholar] [PubMed]
- Versalovic, J.; Carroll, K.C.; Funke, G.; Jorgensen, J.H.; Landry, M.L.; Warnock, D.W. Manual of Clinical Microbiology, 10th ed.; American Society of Microbiology: Washington, DC, USA, 2011. [Google Scholar]
- ISO. 20776-1: 2019-Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices—Part 1: Broth Micro-Dilution Reference Method for Testing the In Vitro Activity of Antimicrobial Agents Against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases; ISO: Geneva, Switzerland, 2019. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; CLSI Supplement M100, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Vingataramin, L.; Frost, E.H. A single protocol for extraction of gDNA from bacteria and yeast. BioTechniques 2015, 58, 120–125. [Google Scholar] [CrossRef]
- Han, H.; Zhou, H.; Li, H.; Gao, Y.; Lu, Z.; Hu, K.; Xu, B. Optimization of pulse-field gel electrophoresis for subtyping of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2013, 10, 2720–2731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göttig, S.; Riedel-Christ, S.; Saleh, A.; Kempf, V.A.J.; Hamprecht, A. Impact of blaNDM-1 on fitness and pathogenicity of Escherichia coli and Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2016, 47, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Chinali, G. A modified alkaline lysis method for the preparation of highly purified plasmid DNA from Escherichia coli. Anal. Biochem. 1993, 212, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Barton, B.M.; Harding, G.P.; Zuccarelli, A.J. A general method for detecting and sizing large plasmids. Anal. Biochem. 1995, 226, 235–240. [Google Scholar] [CrossRef]
- Southern, E. Southern blotting. Nat. Protoc. 2006, 1, 518–525. [Google Scholar] [CrossRef]
- Pesesky, M.W.; Hussain, T.; Wallace, M.; Wang, B.; Andleeb, S.; Burnham, C.A.; Dantas, G. KPC and NDM-1 genes in related Enterobacteriaceae strains and plasmids from Pakistan and the United States. Emerg. Infect. Dis. 2015, 21, 1034–1037. [Google Scholar] [CrossRef]
- Qamar, M.U.; Walsh, T.R.; Toleman, M.A.; Tyrrell, J.M.; Saleem, S.; Aboklaish, A.; Jahan, S. Dissemination of genetically diverse NDM-1, -5, -7 producing-Gram-negative pathogens isolated from pediatric patients in Pakistan. Future Microbiol. 2019, 14, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Gondal, A.J.; Saleem, S.; Jahan, S. Novel Carbapenem-Resistant Klebsiella pneumoniae ST147 Coharboring bla (NDM-1), bla (OXA-48) and Extended-Spectrum β-Lactamases from Pakistan. Infect. Drug Resist. 2020, 13, 2105–2115. [Google Scholar] [CrossRef] [PubMed]
- Masseron, A.; Poirel, L.; Jamil Ali, B.; Syed, M.A.; Nordmann, P. Molecular characterization of multidrug-resistance in Gram-negative bacteria from the Peshawar teaching hospital, Pakistan. New Microbes New Infect. 2019, 32, 100605. [Google Scholar] [CrossRef]
- Humayun, A.; Siddiqui, F.M.; Akram, N.; Saleem, S.; Ali, A.; Iqbal, T.; Kumar, A.; Kamran, R.; Bokhari, H. Incidence of metallo-beta-lactamase-producing Klebsiella pneumoniae isolates from hospital setting in Pakistan. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2018, 21, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Nahid, F.; Khan, A.A.; Rehman, S.; Zahra, R. Prevalence of metallo-β-lactamase NDM-1-producing multi-drug resistant bacteria at two Pakistani hospitals and implications for public health. J. Infect. Public Health 2013, 6, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Voulgari, E.; Gartzonika, C.; Vrioni, G.; Politi, L.; Priavali, E.; Levidiotou-Stefanou, S.; Tsakris, A. The Balkan region: NDM-1-producing Klebsiella pneumoniae ST11 clonal strain causing outbreaks in Greece. J. Antimicrob. Chemother. 2014, 69, 2091–2097. [Google Scholar] [CrossRef] [PubMed]
- Markovska, R.; Stoeva, T.; Boyanova, L.; Stankova, P.; Schneider, I.; Keuleyan, E.; Mihova, K.; Murdjeva, M.; Sredkova, M.; Lesseva, M.; et al. Multicentre investigation of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in Bulgarian hospitals—Interregional spread of ST11 NDM-1-producing K. pneumoniae. Infect. Genet. Evol. 2019, 69, 61–67. [Google Scholar] [CrossRef]
- Dsouza, R.; Pinto, N.A.; Hwang, I.; Younjee, H.; Cho, Y.; Kim, H.; Yong, D.; Choi, J.; Lee, K.; Chong, Y. Molecular epidemiology and resistome analysis of multidrug-resistant ST11 Klebsiella pneumoniae strain containing multiple copies of extended-spectrum β-lactamase genes using whole-genome sequencing. New Microbiol. 2017, 40, 38–44. [Google Scholar] [PubMed]
- Sonnevend, A.; Al Baloushi, A.; Ghazawi, A.; Hashmey, R.; Girgis, S.; Hamadeh, M.B.; Al Haj, M.; Pál, T. Emergence and spread of NDM-1 producer Enterobacteriaceae with contribution of IncX3 plasmids in the United Arab Emirates. J. Med. Microbiol. 2013, 62, 1044–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solgi, H.; Badmasti, F.; Giske, C.G.; Aghamohammad, S.; Shahcheraghi, F. Molecular epidemiology of NDM-1- and OXA-48-producing Klebsiella pneumoniae in an Iranian hospital: Clonal dissemination of ST11 and ST893. J. Antimicrob. Chemother. 2018, 73, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, S.; Crawford, M.A.; Lascols, C.; Timme, R.E.; Anderson, K.; Hodge, D.R.; Fisher, D.J.; Pillai, S.P.; Morse, S.A.; Khan, E.; et al. Resistome of carbapenem- and colistin-resistant Klebsiella pneumoniae clinical isolates. PLoS ONE 2018, 13, e0198526. [Google Scholar] [CrossRef]
- Baloch, Z.; Lv, L.; Yi, L.; Wan, M.; Aslam, B.; Yang, J.; Liu, J.H. Emergence of Almost Identical F36:A-:B32 Plasmids Carrying bla (NDM-5) and qepA in Escherichia coli from Both Pakistan and Canada. Infect. Drug Resist. 2019, 12, 3981–3985. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.-L.; Li, Z.; Lo, W.-U.; Cheung, Y.-Y.; Lin, C.-H.; Sham, P.-C.; Cheng, V.C.-C.; Ng, T.-K.; Que, T.-L.; Chow, K.-H. Identification and characterization of a novel incompatibility group X3 plasmid carrying blaNDM-1 in Enterobacteriaceae isolates with epidemiological links to multiple geographical areas in China. Emerg. Microbes Infect. 2012, 1, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Vázquez, M.; Sola Campoy, P.J.; Ortega, A.; Bautista, V.; Monzón, S.; Ruiz-Carrascoso, G.; Mingorance, J.; González-Barberá, E.M.; Gimeno, C.; Aracil, B.; et al. Emergence of NDM-producing Klebsiella pneumoniae and Escherichia coli in Spain: Phylogeny, resistome, virulence and plasmids encoding blaNDM-like genes as determined by WGS. J. Antimicrob. Chemother. 2019, 74, 3489–3496. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, Y.; Wang, G.; Li, C.; Chang, Y.F.; Chen, W.; Nian, S.; Mao, Y.; Zhang, J.; Zhong, F.; et al. bla (NDM-5) carried by a hypervirulent Klebsiella pneumoniae with sequence type 29. Antimicrob. Resist. Infect. Control. 2019, 8, 140. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbapenem Resistance Genes | N# (%) | ESBLs Encoding Genes | N * (%) |
|---|---|---|---|
| blaNDM-1 | 23 (18.4%) | blaCTXM-15 | 17 (13.6%) |
| blaTEM-1 | 1 (0.8%) | ||
| blaSHV-75 | 4 (3.2%) | ||
| blaIMP-1 | 9 (7.2%) | blaCTXM-15 | 3 (2.4%) |
| blaOXA-1 | 4 (3.2%) | ||
| blaTEM-1 | 2 (1.6%) | ||
| blaVIM-1 | 4 (3.2%) | blaCTXM-15 | 2 (1.6%) |
| blaOXA-48 | 3 (2.4%) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilal, H.; Zhang, G.; Rehman, T.; Han, J.; Khan, S.; Shafiq, M.; Yang, X.; Yan, Z.; Yang, X. First Report of blaNDM-1 Bearing IncX3 Plasmid in Clinically Isolated ST11 Klebsiella pneumoniae from Pakistan. Microorganisms 2021, 9, 951. https://doi.org/10.3390/microorganisms9050951
Bilal H, Zhang G, Rehman T, Han J, Khan S, Shafiq M, Yang X, Yan Z, Yang X. First Report of blaNDM-1 Bearing IncX3 Plasmid in Clinically Isolated ST11 Klebsiella pneumoniae from Pakistan. Microorganisms. 2021; 9(5):951. https://doi.org/10.3390/microorganisms9050951
Chicago/Turabian StyleBilal, Hazrat, Gaojian Zhang, Tayyab Rehman, Jianxion Han, Sabir Khan, Muhammad Shafiq, Xuegang Yang, Zhongkang Yan, and Xingyuan Yang. 2021. "First Report of blaNDM-1 Bearing IncX3 Plasmid in Clinically Isolated ST11 Klebsiella pneumoniae from Pakistan" Microorganisms 9, no. 5: 951. https://doi.org/10.3390/microorganisms9050951