
Low-Level Tolerance to Fluoroquinolone Antibiotic Ciprofloxacin in QAC-Adapted Subpopulations of Listeria monocytogenes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Listeria monocytogenes Strains and Growth Conditions
2.2. Preparation of QAC Solutions
2.3. L. monocytogenes Adaptation to Sublethal QAC
2.4. Preparation of QAC-Adapted Subpopulation 1 in Broth Model (QAC-P1)
2.5. Preparation of QAC-Adapted Subpopulation 2 in Broth Model (QAC-P2)
2.6. Preparation of QAC-Adapted Subpopulation 3 in Water Model (QAC-P3)
2.7. Preparation of Ciprofloxacin Solutions
2.8. Determination of Short-Range MIC of Ciprofloxacin for QAC-Adapted Subpopulations and Non-Adapted Control of L. monocytogenes Strains
2.9. Determination of Growth Kinetics of QAC-Adapted Subpopulations and Non-Adapted Control of L. monocytogenes Strains in Ciprofloxacin-Containing Broth
2.10. Determination of Survival of QAC-Adapted Subpopulations and Non-Adapted Control of L. monocytogenes Strains in Ciprofloxacin-Containing Agar
2.11. Statistical Analysis
3. Results
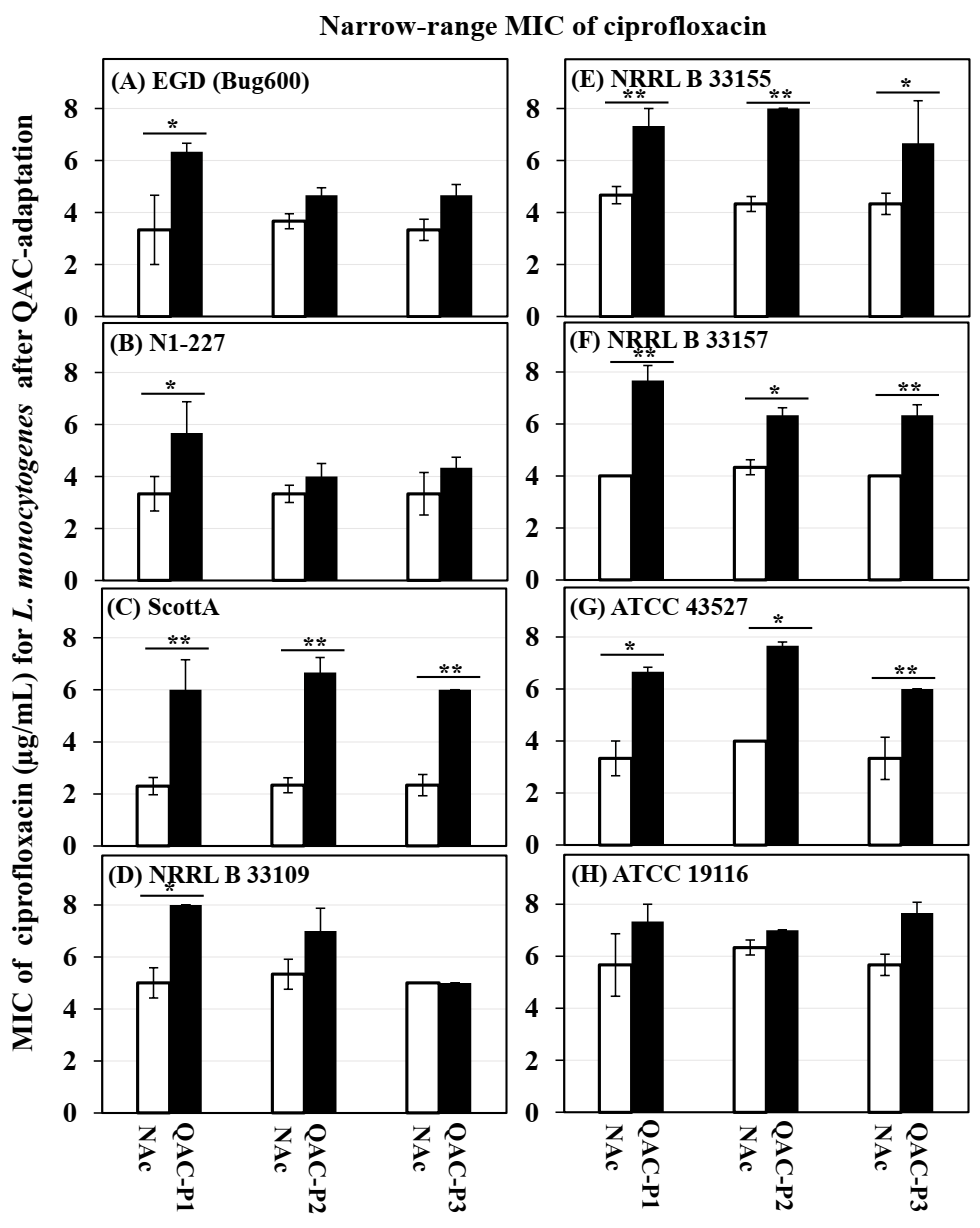
3.1. Changes in Short-Range MIC of Ciprofloxacin for QAC-Adapted Subpopulations of L. monocytogenes
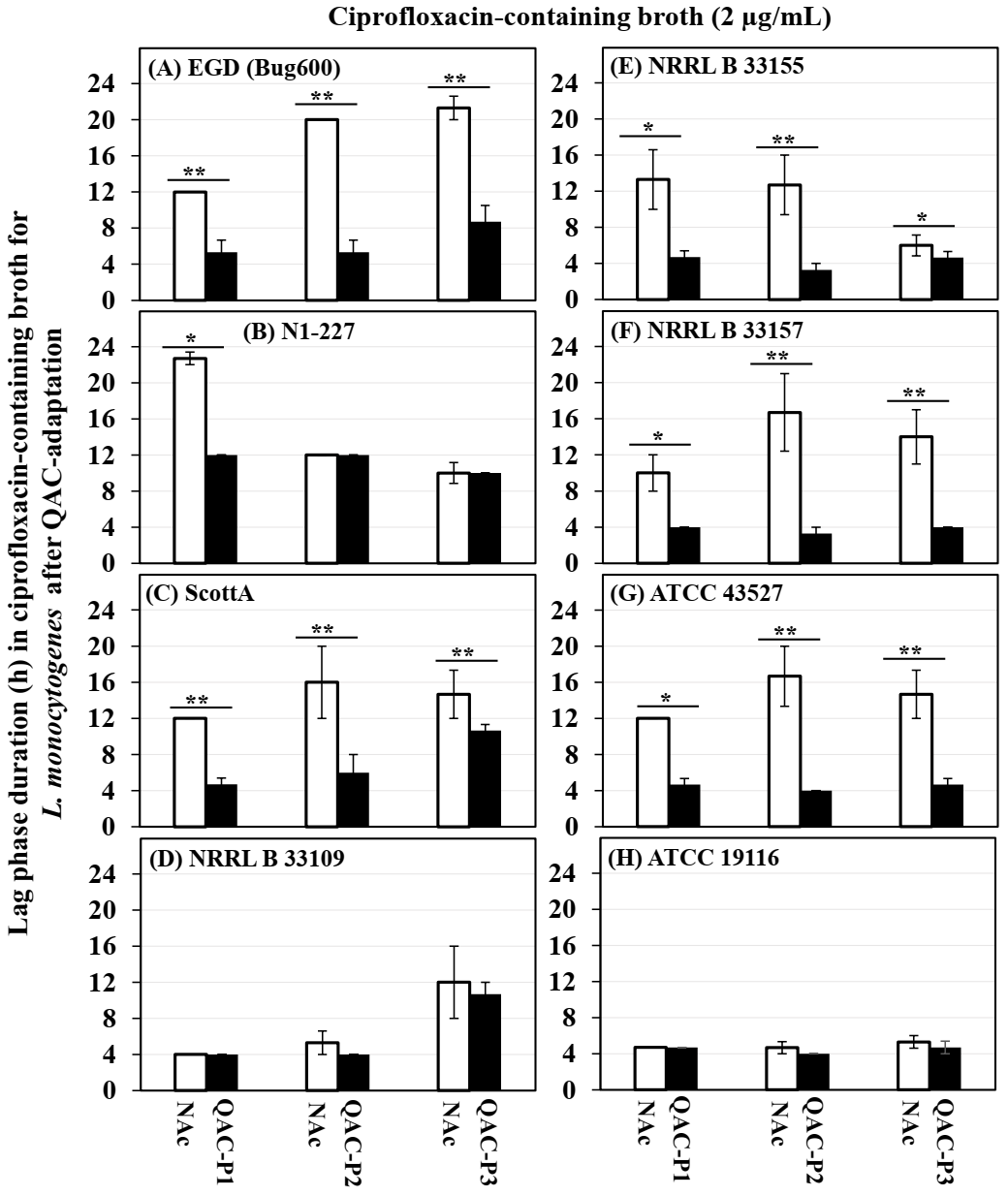
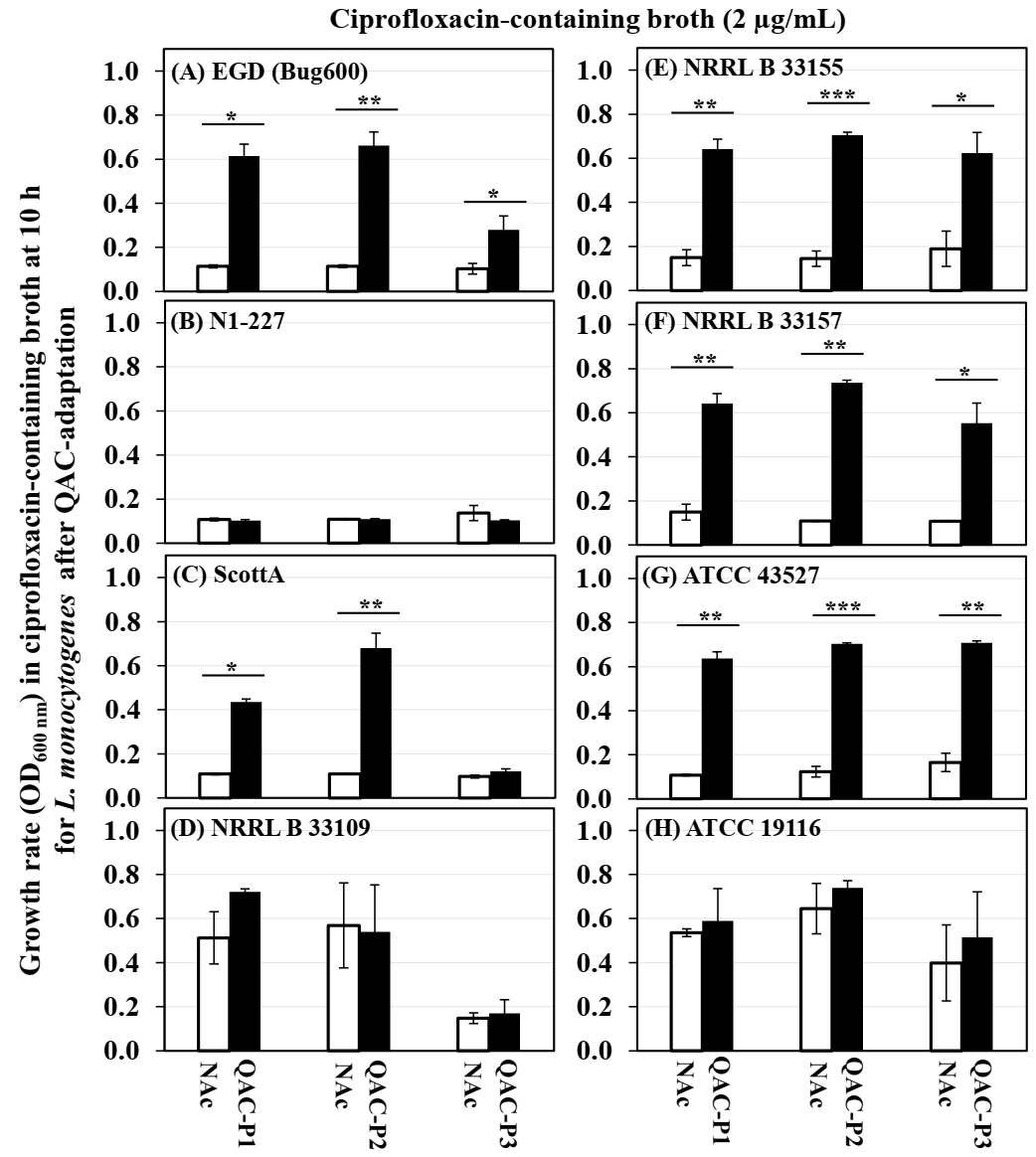
3.2. Changes in the Growth Rate of QAC-Adapted Subpopulations of L. monocytogenes in Ciprofloxacin-Containing Broth
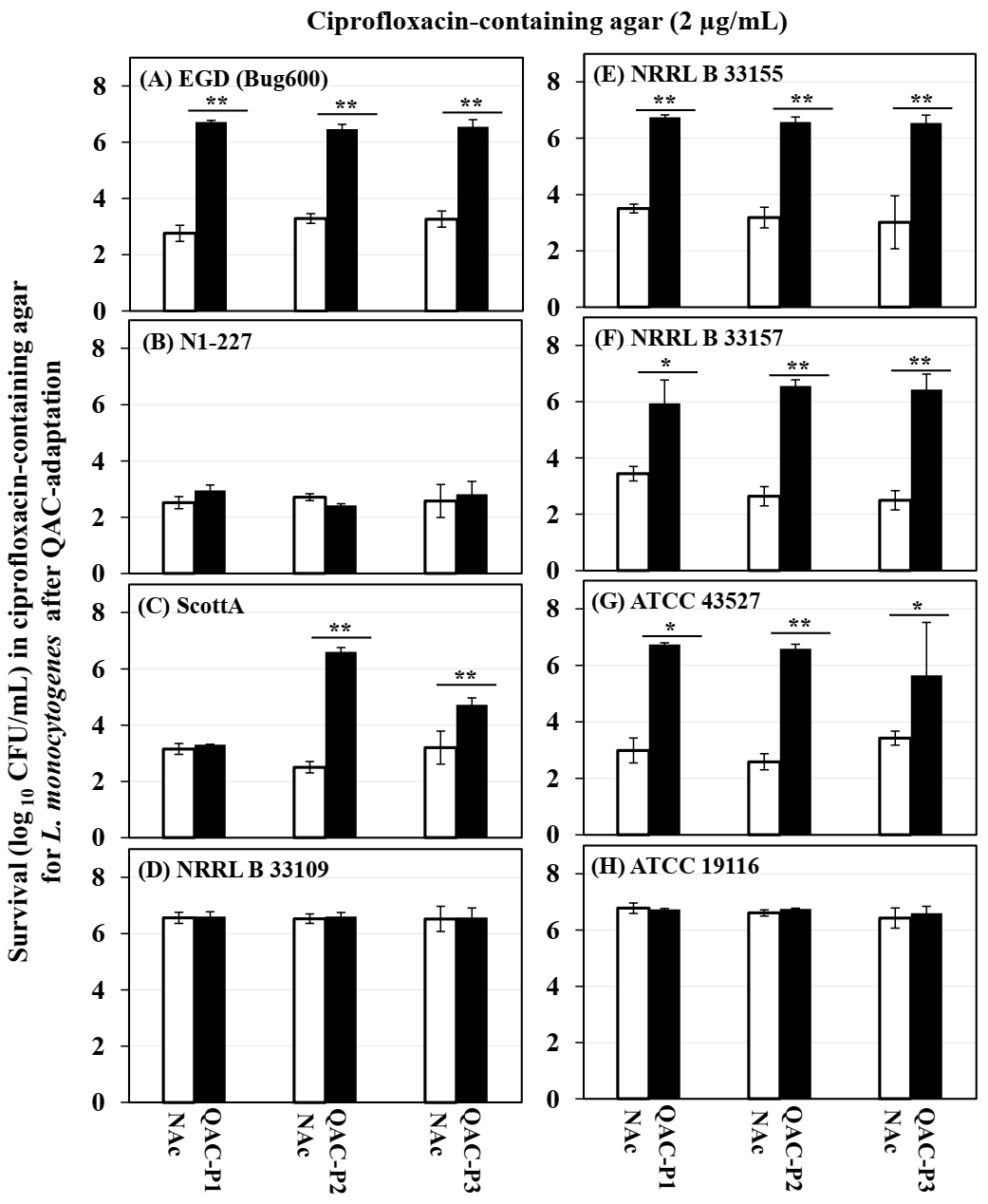
3.3. Changes in the Survival of QAC-Adapted Subpopulations of L. monocytogenes in Ciprofloxacin-Containing Agar
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poyart-Salmeron, C.; Carlier, C.; Trieu-Cuot, P.; Courvalin, P.; Courtieu, A.-L. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef]
- Zhang, Y.; Yeh, E.; Hall, G.; Cripe, J.; Bhagwat, A.A.; Meng, J. Characterization of Listeria monocytogenes isolated from retail foods. Int. J. Food Microbiol. 2007, 113, 47–53. [Google Scholar] [CrossRef]
- Conter, M.; Paludi, D.; Zanardi, E.; Ghidini, S.; Vergara, A.; Ianieri, A. Characterization of antimicrobial resistance of foodborne Listeria monocytogenes. Int. J. Food Microbiol. 2009, 128, 497–500. [Google Scholar] [CrossRef]
- Alonso-Hernando, A.; Prieto, M.; García-Fernández, C.; Alonso-Calleja, C.; Capita, R. Increase over time in the prevalence of multiple antibiotic resistance among isolates of Listeria monocytogenes from poultry in Spain. Food Control 2012, 23, 37–41. [Google Scholar] [CrossRef]
- Chen, M.; Cheng, J.; Zhang, J.; Chen, Y.; Zeng, H.; Xue, L.; Lei, T.; Pang, R.; Wu, S.; Wu, H.; et al. Isolation, potential virulence, and population diversity of Listeria monocytogenes from meat and meat products in China. Front. Microbiol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gerba, C.P. Quaternary ammonium biocides: Efficacy in application. Appl. Environ. Microbiol. 2015, 81, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Bashor, M.P.; Curtis, P.A.; Keener, K.M.; Sheldon, B.W.; Kathariou, S.; Osborne, J.A. Effects of Carcass Washers on Campylobacter Contamination in Large Broiler Processing Plants. Poult. Sci. 2004, 83, 1232–1239. [Google Scholar] [CrossRef]
- Iñiguez-Moreno, M.; Avila-Novoa, M.G.; Gutiérrez-Lomelí, M. Resistance of pathogenic and spoilage microorganisms to disinfectants in the presence of organic matter and their residual effect on stainless steel and polypropylene. J. Glob. Antimicrob. Resist. 2018, 14, 197–201. [Google Scholar] [CrossRef]
- Tezel, U.; Pavlostathis, S.G. Quaternary ammonium disinfectants: Microbial adaptation, degradation and ecology. Curr. Opin. Biotechnol. 2015, 33, 296–304. [Google Scholar] [CrossRef]
- Møretrø, T.; Schirmer, B.C.T.; Heir, E.; Fagerlund, A.; Hjemli, P.; Langsrud, S. Tolerance to quaternary ammonium compound disinfectants may enhance growth of Listeria monocytogenes in the food industry. Int. J. Food Microbiol. 2017, 241, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Cerf, O.; Carpentier, B.; Sanders, P. Tests for determining in-use concentrations of antibiotics and disinfectants are based on entirely different concepts: “Resistance” has different meanings. Int. J. Food Microbiol. 2010, 136, 247–254. [Google Scholar] [CrossRef]
- Aase, B.; Sundheim, G.; Langsrud, S.; Rorvik, L.M. Occurrence of and a possible mechanism for resistance to a quaternary ammonium compound in Listeria monocytogenes. Int. J. Food Microbiol. 2000, 62, 57–63. [Google Scholar] [CrossRef]
- Carpentier, B.; Cerf, O. Review—Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes Persistence in Food-Associated Environments: Epidemiology, Strain Characteristics, and Implications for Public Health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef]
- Fox, E.M.; Leonard, N.; Jordan, K. Physiological and Transcriptional Characterization of Persistent and Nonpersistent Listeria monocytogenes Isolates. Appl. Environ. Microbiol. 2011, 77, 6559–6569. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, S.; Lopez, V.; Martinez-Suarez, J.V. Control of Listeria monocytogenes contamination in an Iberian pork processing plant and selection of benzalkonium chloride-resistant strains. Food Microbiol. 2014, 39, 81–88. [Google Scholar] [CrossRef]
- Romanova, N.A.; Wolffs, P.F.G.; Brovko, L.Y.; Griffiths, M.W. Role of Efflux Pumps in Adaptation and Resistance of Listeria monocytogenes to Benzalkonium Chloride Role of Efflux Pumps in Adaptation and Resistance of Listeria monocytogenes to Benzalkonium Chloride. Appl. Environ. Microbiol. 2006, 72, 3498–3503. [Google Scholar] [CrossRef] [Green Version]
- Soumet, C.; Méheust, D.; Pissavin, C.; Le Grandois, P.; Frémaux, B.; Feurer, C.; Le Roux, A.; Denis, M.; Maris, P. Reduced susceptibilities to biocides and resistance to antibiotics in food-associated bacteria following exposure to quaternary ammonium compounds. J. Appl. Microbiol. 2016, 121, 1275–1281. [Google Scholar] [CrossRef]
- Sereno, M.J.; Viana, C.; Pegoraro, K.; da Silva, D.A.L.; Yamatogi, R.S.; Nero, L.A.; Bersot, L. dos S. Distribution, adhesion, virulence and antibiotic resistance of persistent Listeria monocytogenes in a pig slaughterhouse in Brazil. Food Microbiol. 2019, 84, 103234. [Google Scholar] [CrossRef]
- Morvan, A.; Moubareck, C.; Leclercq, A.; Hervé-Bazin, M.; Bremont, S.; Lecuit, M.; Courvalin, P.; Le Monnier, A. Antimicrobial resistance of Listeria monocytogenes strains isolated from humans in France. Antimicrob. Agents Chemother. 2010, 54, 2728–2731. [Google Scholar] [CrossRef] [Green Version]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes strains selected on ciprofloxacin or the disinfectant benzalkonium chloride exhibit reduced susceptibility to ciprofloxacin, gentamicin, benzalkonium chloride, and other toxic compounds. Appl. Environ. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Jiang, X.; Zhang, Y.; Ji, S.; Gao, W.; Shi, L. Effect of Benzalkonium Chloride adaptation on sensitivity to antimicrobial agents and tolerance to environmental stresses in Listeria monocytogenes. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Theophel, K.; Schacht, V.J.; Schlüter, M.; Schnell, S.; Stingu, C.-S.; Schaumann, R.; Bunge, M. The importance of growth kinetic analysis in determining bacterial susceptibility against antibiotics and silver nanoparticles. Front. Microbiol. 2014, 5, 544. [Google Scholar] [CrossRef]
- Wales, A.; Davies, R. Co-Selection of Resistance to Antibiotics, Biocides and Heavy Metals, and Its Relevance to Foodborne Pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef] [Green Version]
- Guérin, A.; Bridier, A.; Le Grandois, P.; Sévellec, Y.; Palma, F.; Félix, B.; Roussel, S.; Soumet, C.; Karpíšková, R.; Pomelio, F.; et al. Exposure to quaternary ammonium compounds selects resistance to ciprofloxacin in Listeria monocytogenes. Pathogens 2021, 10, 220. [Google Scholar] [CrossRef]
- Mc Cay, P.H.; Ocampo-Sosa, A.A.; Fleming, G.T.A. Effect of subinhibitory concentrations of benzalkonium chloride on the competitiveness of Pseudomonas aeruginosa grown in continuous culture. Microbiology 2010, 156, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Ching, C.; Zaman, M.H. Development and selection of low-level multi-drug resistance over an extended range of sub-inhibitory ciprofloxacin concentrations in Escherichia coli. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Goldstein, F. The potential clinical impact of low-level antibiotic resistance in Staphylococcus aureus. J. Antimicrob. Chemother. 2007, 59, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Qiu, Y.; Shi, H.; Yin, H. The importance of lag time extension in determining bacterial resistance to antibiotics. Analyst 2016, 141, 3059–3067. [Google Scholar] [CrossRef] [Green Version]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418–421. [Google Scholar] [CrossRef]
- Tamburro, M.; Ripabelli, G.; Vitullo, M.; Dallman, T.J.; Pontello, M.; Amar, C.F.L.; Sammarco, M.L. Gene expression in Listeria monocytogenes exposed to sublethal concentration of benzalkonium chloride. Comp. Immunol. Microbiol. Infect. Dis. 2015, 40, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Hernando, A.; Capita, R.; Prieto, M.; Alonso-Calleja, C. Comparison of antibiotic resistance patterns in Listeria monocytogenes and Salmonella enterica strains pre-exposed and exposed to poultry decontaminants. Food Control 2009, 20, 1108–1111. [Google Scholar] [CrossRef]
- Godreuil, S.; Galimand, M.; Gerbaud, G.; Jacquet, C.; Courvalin, P. Efflux pump Lde is associated with fluoroquinolone resistance in Listeria monocytogenes. Antimicrob. Agents Chemother. 2003, 47, 704–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Designation | Lineage | Serotype | Source | Isolation Source |
|---|---|---|---|---|---|
| L. monocytogenes | N1-227 | I | 4b | CDC, Atlanta | Food epidemic |
| L. monocytogenes | ATCC 19116 | III | 4c | University of Wisconsin | Poultry |
| L. monocytogenes | ScottA | I | 4b | FDA | Human epidemic |
| L. monocytogenes | EGD (Bug600) | II | 1/2a | Institute Pasteur | Guinea pigs |
| L. monocytogenes | NRRL B-33109 | I | 4b | USDA-ARS, NADC | Cooler condenser |
| L. monocytogenes | NRRL B-33155 | I | 4b | USDA-ARS, NADC | Sodium caesinate epidemic strain, CA, 1985 outbreak |
| L. monocytogenes | NRRL B-33157 | I | 4b | USDA-ARS, NADC | Insect debris found in cheese plant |
| L. monocytogenes | ATCC 43257 | I | 4b | CDC, Atlanta | Mexican Style cheese, CA |
| Listeria monocytogenes Strains | Fold Increase 1 in MIC of Ciprofloxacin for QAC-P1 ± SE | Fold Increase 2 in Growth (OD600) of QAC-P1 Compared to Non-Adapted Control in Ciprofloxacin-Containing Broth at Different Time Points | Percentage Increase 3 in Survivals in Ciprofloxacin-Agar for QAC-P1 ± SE | |||
|---|---|---|---|---|---|---|
| 10 h | 12 h | 20 h | 24 h | |||
| EGD (Bug600) | 1.9 ± 0.2 a | 4.5 ± 0.8 a | 5.3 ± 1.0 a | 2.4 ± 0.4 a | 1.9 ± 0.4 b | 58.3 ± 5.4 a |
| N1 227 | 1.7 ± 0.3 b | 1.0 ± 0.0 c | 0.9± 0.0 b | 2.4 ± 0.5 a | 4.0 ± 0.2 a | 5.77 ± 0.5 c |
| Scott A | 2.6 ± 0.3 a | 4.0 ± 0.2 a | 4.8 ± 0.2 a | 3.1 ± 0.9 a | 2.2 ± 1.0 a | 1.75 ± 2.7 c |
| NRRL B 33109 | 1.6 ± 0.2 b | 1.4 ± 0.4 b | 1.4 ± 0.5 b | 1.2 ± 0.3 b | 1.1 ± 0.1 d | 1.20 ± 0.3 c |
| NRRL B 33155 | 1.6 ± 0.2 b | 3.9 ± 0.7 a | 3.4 ± 1.3 a | 2.3 ± 1.1 a | 1.5 ± 0.2 c | 46.4 ± 1.7 a |
| NRRL B 33157 | 1.9 ± 0.1 a | 3.6 ± 0.4 a | 4.3 ± 0.8 a | 1.0 ± 0.1 b | 1.0 ± 0.1 d | 36.7 ± 13.6 b |
| ATCC 43257 | 2.0 ± 0.4 a | 5.0 ± 1.1 a | 5.9 ± 0.2 a | 2.5 ± 0.4 a | 2.2 ± 0.4 b | 55.6 ± 10.6 a |
| ATCC 19116 | 1.3 ± 0.1 b | 1.1 ± 0.3 c | 1.1 ± 0.2 b | 1.2 ± 0.0 b | 1.1 ± 0.1 d | 0.80 ± 0.4 c |
| Listeria monocytogenes Strains | Fold Increase 1 in MIC of Ciprofloxacin for QAC-P2 ± SE | Fold Increase 2 in Growth (OD600) of QAC-P2 Compared to Non-Adapted Control in Ciprofloxacin-Containing Broth at Different Time Points | Percentage Increase 3 in Survivals in Ciprofloxacin-Agar for QAC-P2 ± SE | |||
| 10 h | 12 h | 20 h | 24 h | |||
| EGD (Bug600) | 1.3 ± 0.1 c | 4.9 ± 0.9 a | 5.6 ± 1.0 a | 5.1 ± 0.4 a | 2.1 ± 0.4 a | 46.8 ± 4.1 b |
| N1 227 | 1.2 ± 0.3 c | 1.0 ± 0.0 b | 1.0 ± 0.0 c | 1.0 ± 0.5 b | 1.3 ± 0.2 a | −4.6 ± 2.3 d |
| Scott A | 2.9 ± 0.1 a | 5.1 ± 0.2 a | 5.6 ± 0.2 a | 3.7 ± 0.9 a | 1.6 ± 1.0 a | 60.3 ± 2.6 a |
| NRRL B 33109 | 1.3 ± 0.0 c | 0.9 ± 0.4 b | 0.8 ± 0.5 c | 0.9 ± 0.3 b | 1.0 ± 0.1 b | 0.90 ± 0.8 c |
| NRRL B 33155 | 1.8 ± 0.1 b | 3.3 ± 0.7 a | 2.6 ± 1.3 b | 2.0 ± 1.1 a | 1.2 ± 0.2 a | 49.8 ± 7.1 b |
| NRRL B 33157 | 1.5 ± 0.0 b | 6.8 ± 0.4 a | 5.7 ± 0.8 a | 2.2 ± 0.1 a | 1.2 ± 0.1 a | 58.9 ± 3.9 a |
| ATCC 43257 | 1.9 ± 0.1 b | 6.7 ± 1.1 a | 5.1 ± 0.2 a | 3.1 ± 0.4 a | 2.1 ± 0.4 a | 59.7 ± 4.2 a |
| ATCC 19116 | 1.2 ± 0.1 c | 1.1 ± 0.3 b | 1.0 ± 0.2 c | 1.0 ± 0.0 b | 1.0 ± 0.1 b | 2.85 ± 1.7 c |
| Listeria monocytogenes Strains | Fold Increase 1 in MIC of Ciprofloxacin for QAC-P3 ± SE | Fold Increase 2 in Growth (OD600) of QAC-P3 Compared to Non-Adapted Control in Ciprofloxacin-Containing Broth at Different Time Points | Percentage Increase 3 in Survivals in Ciprofloxacin-Agar for QAC-P3 ± SE | |||
| 10 h | 12 h | 20 h | 24 h | |||
| EGD (Bug600) | 1.4 ± 0.1 b | 2.1 ± 0.6 b | 3.1 ± 1.1 b | 3.4 ± 0.7 a | 2.3 ± 1.0 a | 48.1 ± 1.9 a |
| N1 227 | 1.3 ± 0.3 b | 0.8 ± 0.2 c | 1.2 ± 0.4 c | 1.3 ± 0.4 b | 1.2 ± 0.1 b | 3.3 ± 5.4 d |
| Scott A | 2.6 ± 0.3 a | 1.2 ± 0.2 c | 1.8 ± 0.6 c | 2.0 ± 0.3 a | 1.2 ± 0.3 b | 22.6 ± 6.8 c |
| NRRL B 33109 | 1.0 ± 0.0 c | 1.1 ± 0.8 c | 0.8 ± 1.7 c | 1.2 ± 1.5 b | 1.1 ± 0.4 b | 0.41 ± 1.3 d |
| NRRL B 33155 | 1.5 ± 0.3 a | 1.7 ± 1.0 c | 2.2 ± 1.5 b | 1.6 ± 1.4 a | 1.2 ± 0.1 b | 52.1 ± 5.0 a |
| NRRL B 33157 | 1.6 ± 0.1 b | 5.1 ± 0.7 a | 5.2 ± 0.7 a | 1.3 ± 1.4 b | 1.0 ± 0.0 b | 57.3 ± 7.5 a |
| ATCC 43257 | 1.8 ± 0.5 a | 6.4 ± 0.7 a | 6.6 ± 0.6 a | 3.5 ± 1.0 a | 1.9 ± 0.2 a | 33.1 ± 17.6 b |
| ATCC 19116 | 1.4 ± 0.1 b | 1.3 ± 2.5 c | 1.1 ± 2.5 c | 0.9 ± 0.2 b | 0.9 ± 0.1 b | 3.24 ± 1.1 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kode, D.; Nannapaneni, R.; Bansal, M.; Chang, S.; Cheng, W.-H.; Sharma, C.S.; Kiess, A. Low-Level Tolerance to Fluoroquinolone Antibiotic Ciprofloxacin in QAC-Adapted Subpopulations of Listeria monocytogenes. Microorganisms 2021, 9, 1052. https://doi.org/10.3390/microorganisms9051052
Kode D, Nannapaneni R, Bansal M, Chang S, Cheng W-H, Sharma CS, Kiess A. Low-Level Tolerance to Fluoroquinolone Antibiotic Ciprofloxacin in QAC-Adapted Subpopulations of Listeria monocytogenes. Microorganisms. 2021; 9(5):1052. https://doi.org/10.3390/microorganisms9051052
Chicago/Turabian StyleKode, Divya, Ramakrishna Nannapaneni, Mohit Bansal, Sam Chang, Wen-Hsing Cheng, Chander S. Sharma, and Aaron Kiess. 2021. "Low-Level Tolerance to Fluoroquinolone Antibiotic Ciprofloxacin in QAC-Adapted Subpopulations of Listeria monocytogenes" Microorganisms 9, no. 5: 1052. https://doi.org/10.3390/microorganisms9051052