
Mild Lactic Acid Stress Causes Strain-Dependent Reduction in SEC Protein Levels




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, and Sample Collection for sec mRNA and SEC Protein Quantification
2.2. RNA Extraction
2.3. Reverse-Transcription and Quantitative Real-Time PCR
2.4. Protein Quantification
3. Results
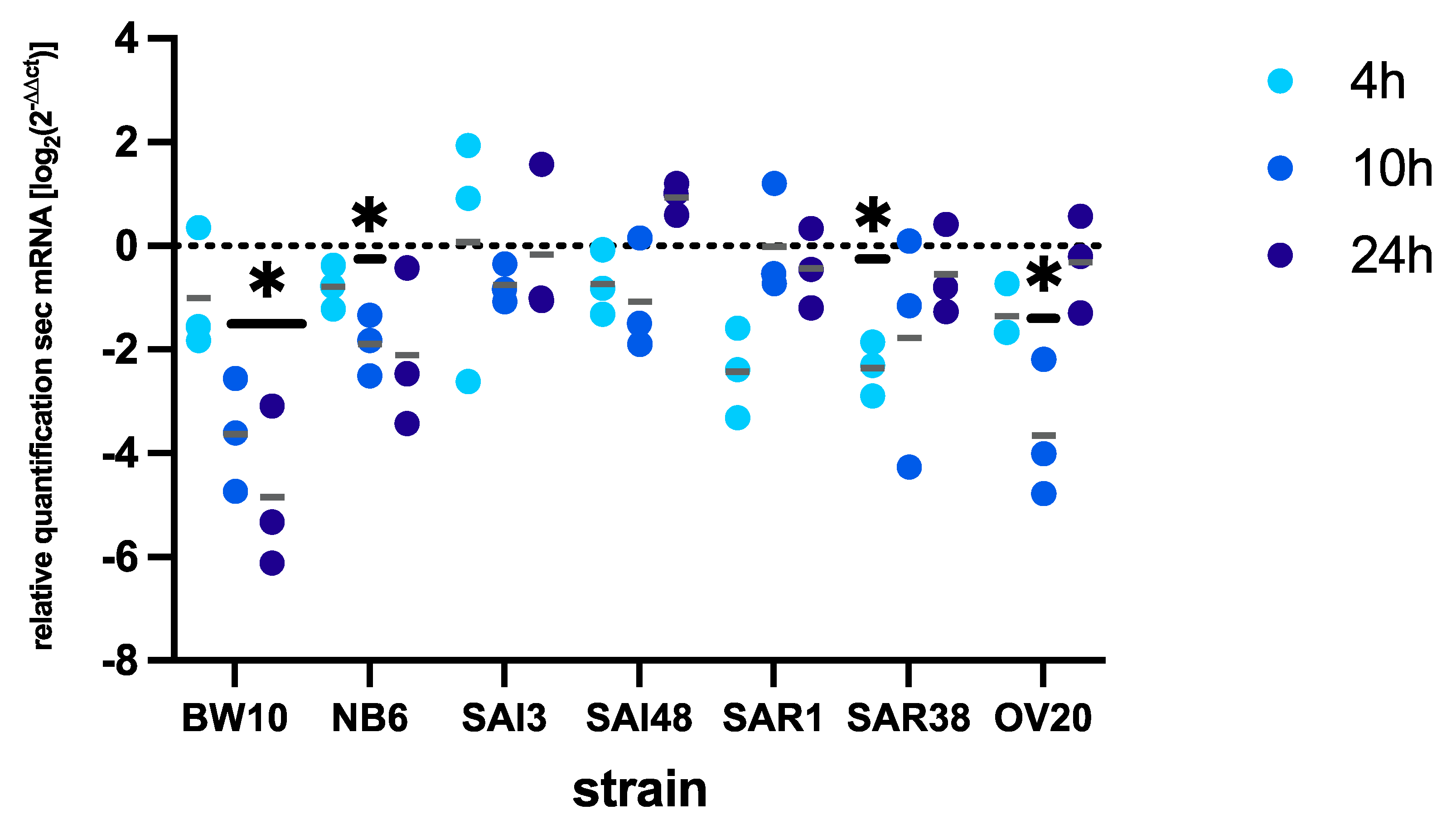
3.1. Effect of Mild Lactic Acid Stress (pH 6.0) on Bacterial Growth and sec mRNA Levels
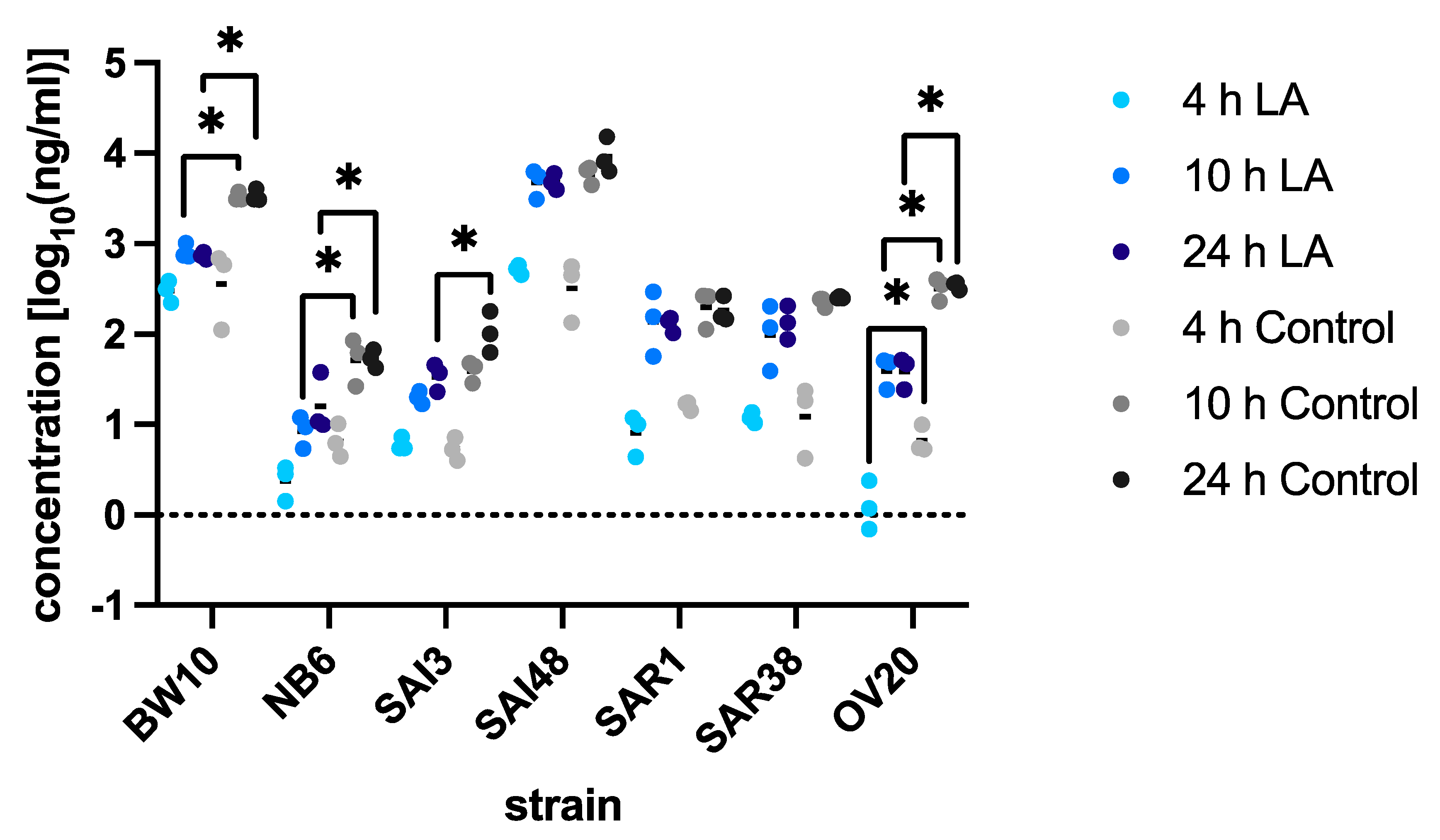
3.2. SEC Protein Levels under Lactic Acid Stress
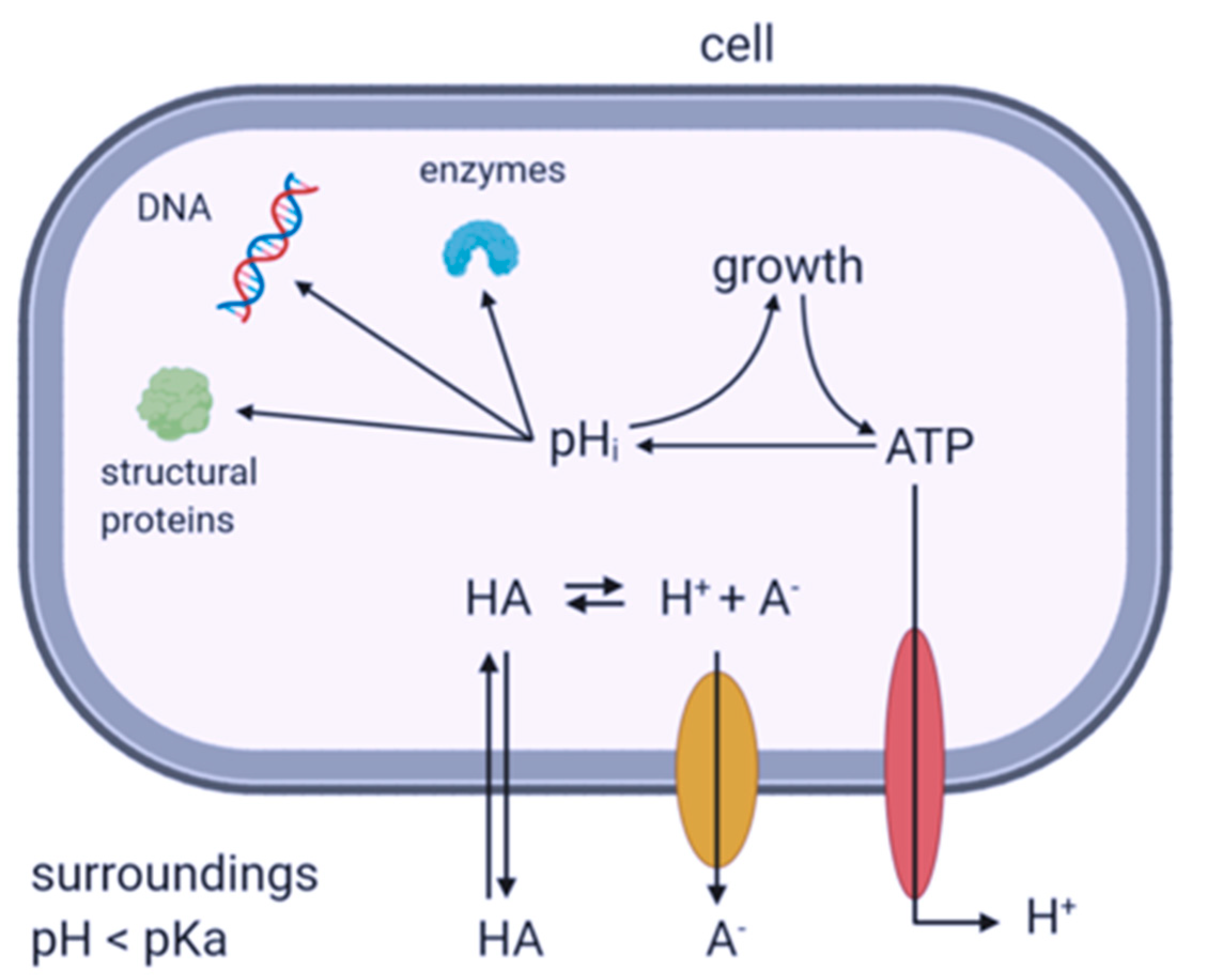
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fetsch, A.; Johler, S. Staphylococcus aureus as a foodborne pathogen. Curr. Clin. Microbiol. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Food-Borne Disease Burden Epidemiology Reference Group. WHO Estimates of the Global Burden of Foodborne Diseases; WHO Press: Geneva, Switzerland, 2015. [Google Scholar]
- Langley, R.J.; Fraser, J.D.; Proft, T. Bacterial Superantigens and Superantigen-Like Toxins; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9780128005897. [Google Scholar]
- Johnson, H.M.; Russell, J.K.; Pontzer, C.H. Staphylococcal enterotoxin superantigens. Proc. Soc. Exp. Biol. Med. 1991, 198, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar] [CrossRef]
- Nia, Y.; Mutel, I.; Assere, A.; Lombard, B.; Auvray, F.; Hennekinne, J.A. Review over a 3-year period of european union proficiency tests for detection of staphylococcal enterotoxins in food matrices. Toxins (Basel) 2016, 8, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA The Euroean Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J. 2016, 13. [CrossRef]
- Food, E.; Authority, S.; Centre, E. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13. [Google Scholar] [CrossRef]
- Gould, L.H.; Nisler, A.L.; Herman, K.M.; Cole, D.J.; Williams, I.T.; Mahon, B.E.; Griffin, P.M.; Hall, A.J. Surveillance for foodborne disease outbreaks—United States, 2008. Mmwr. Morb. Mortal. Wkly. Rep. 2011, 60, 1197–1197202. [Google Scholar]
- Etter, D.; Schelin, J.; Schuppler, M.; Johler, S. Staphylococcal Enterotoxin C—An Update on SEC Variants, Their Structure and Properties, and Their Role in Foodborne Intoxications. Toxins 2020, 12, 584. [Google Scholar] [CrossRef]
- Marr, J.C.; Lyon, J.D.; Roberson, J.R.; Lupher, M.; Davis, W.C.; Bohach, G.A. Characterization of novel type C staphylococcal enterotoxins: Biological and evolutionary implications. Infect. Immun. 1993, 61, 4254–4262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viana, D.; Blanco, J.; Tormo-Más, M.Á.; Selva, L.; Guinane, C.M.; Baselga, R.; Corpa, J.M.; Lasa, Í.; Novick, R.P.; Fitzgerald, J.R.; et al. Adaptation of Staphylococcus aureus to ruminant and equine hosts involves SaPI-carried variants of von Willebrand factor-binding protein. Mol. Microbiol. 2010, 77, 1583–1594. [Google Scholar] [CrossRef]
- Hajek, V. Identification of enterotoxigenic staphylococci from sheep and sheep cheese. Appl. Environ. Microbiol. 1978, 35, 264–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunikowski, R.; Mielke, M.E.A.; Skarabis, H.; Worm, M.; Anagnostopoulos, I.; Kolde, G.; Wahn, U.; Renz, H. Evidence for a disease-promoting effect of Staphylococcus aureus-derived exotoxins in atopic dermatitis. J. Allergy Clin. Immunol. 2000, 105, 814–819. [Google Scholar] [CrossRef]
- Benkerroum, N. Staphylococcal enterotoxins and enterotoxin-like toxins with special reference to dairy products: An overview. Crit. Rev. Food Sci. Nutr. 2018, 58, 1943–1970. [Google Scholar] [CrossRef]
- Aragon-Alegro, L.C.; Konta, E.M.; Suzuki, K.; Silva, M.G.; Júnior, A.F.; Rall, R.; Rall, V.L.M. Occurrence of coagulase-positive Staphylococcus in various food products commercialized in Botucatu, SP, Brazil and detection of toxins from food and isolated strains. Food Control 2007, 18, 630–634. [Google Scholar] [CrossRef]
- Lindqvist, R.; Sylve, S. Quantitative microbial risk assessment exemplified by Staphylococcus aureus in unripened cheese made from raw milk. Int. J. Food Microbiol. 2002, 78, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Corti, S.; Muehlherr, J.E.; Zweifel, C.; Stephan, R. Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from raw bulk-tank milk samples of goats and sheep. Vet. Microbiol. 2004, 101, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, Å.; Fabricius, A.; Guss, B.; Sylvén, S.; Lindqvist, R. Occurrence of foodborne pathogens and characterization of Staphylococcus aureus in cheese produced on farm-dairies. Int. J. Food Microbiol. 2010, 144, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Wikler, M.; Cockerill, F.; Craig, W.; Dudley, M.; Eliopoulos, G.; Hecht, D. Performance Standards for Antimicrobial Susceptibility Testing; Seventeenth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2007; Volume 27, ISBN 1562386255. [Google Scholar]
- Moser, A.; Stephan, R.; Corti, S.; Johler, S. Comparison of genomic and antimicrobial resistance features of latex agglutination test-positive and latex agglutination test-negative Staphylococcus aureus isolates causing bovine mastitis. J. Dairy Sci. 2013, 96, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgartner, A.; Niederhauser, I.; Johler, S. Virulence and resistance gene profiles of staphylococcus aureus strains isolated from ready-to-eat foods. J. Food Prot. 2014, 77, 1232–1236. [Google Scholar] [CrossRef] [PubMed]
- Stalder, U.; Stephan, R.; Corti, S.; Bludau, M.; Maeschli, A.; Klocke, P.; Johler, S. Short communication: Staphylococcus aureus isolated from colostrum of dairy heifers represent a closely related group exhibiting highly homogeneous genomic and antimicrobial resistance features. J. Dairy Sci. 2014, 97, 4997–5000. [Google Scholar] [CrossRef]
- EFSA. The European Union One Health 2018 Zoonoses Report. Sci. Rep. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- Fang, R.; Cui, J.; Cui, T.; Guo, H.; Ono, H.; Park, C.-H.; Okamura, M.; Nakane, A.; Hu, D.-L. Staphylococcal Enterotoxin C Is an Important Virulence Factor for Mastitis. Toxins 2019, 11, 141. [Google Scholar] [CrossRef] [Green Version]
- Niskanen, A.; Koranen, L.; Roine, K. Staphylococcal enterotoxin and thermonuclease production during induced bovine mastitis and the clinical reaction of enterotoxin in udders. Infect. Immun. 1978, 19, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, J.R.; Monday, S.R.; Foster, T.J.; Bohach, G.A.; Hartigan, P.J.; Meaney, W.J.; Smyth, C.J. Characterization of a putative pathogenicity island from bovine Staphylococcus aureus encoding multiple superantigens. J. Bacteriol. 2001, 183, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orden, J.A.; Goyache, J.; Hernández, J.; Doménech, A.; Suárez, G.; Gómez-Lucia, E. Detection of enterotoxins and TSST-1 secreted by Staphylococcus aureus isolated from ruminant mastitis. Comparison of ELISA and immunoblot. J. Appl. Bacteriol. 1992, 72, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, Y.C.; Wang, Y.; Li, H.; Wang, X.M.; Wu, Y.; Zhang, D.R.; Gao, S.; Qi, Z.L. Impact of yeast and lactic acid bacteria on mastitis and milk microbiota composition of dairy cows. AMB Express 2020, 10. [Google Scholar] [CrossRef]
- Diepers, A.C.; Krömker, V.; Zinke, C.; Wente, N.; Pan, L.; Paulsen, K.; Paduch, J.H. In vitro ability of lactic acid bacteria to inhibit mastitis-causing pathogens. Sustain. Chem. Pharm. 2017, 5, 84–92. [Google Scholar] [CrossRef]
- Silva, G.O.; Castro, R.D.; Oliveira, L.G.; Sant’Anna, F.M.; Barbosa, C.D.; Sandes, S.H.C.; Silva, R.S.; Resende, M.F.S.; Lana, A.M.Q.; Nunes, A.C.; et al. Viability of Staphylococcus aureus and expression of its toxins (SEC and TSST-1) in cheeses using Lactobacillus rhamnosus D1 or Weissella paramesenteroides GIR16L4 or both as starter cultures. J. Dairy Sci. 2020, 103, 4100–4108. [Google Scholar] [CrossRef]
- Zdenkova, K.; Alibayov, B.; Karamonova, L.; Purkrtova, S.; Karpiskova, R.; Demnerova, K. Transcriptomic and metabolic responses of Staphylococcus aureus in mixed culture with Lactobacillus plantarum, Streptococcus thermophilus and Enterococcus durans in milk. J. Ind. Microbiol. Biotechnol. 2016, 43, 1237–1247. [Google Scholar] [CrossRef]
- Guan, N.; Liu, L. Microbial response to acid stress: Mechanisms and applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Le Marc, Y.; Valík, L.; Medveďová, A. Modelling the effect of the starter culture on the growth of Staphylococcus aureus in milk. Int. J. Food Microbiol. 2009, 129, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Geisinger, E. Quorum Sensing in Staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- Sihto, H.-M.; Tasara, T.; Stephan, R.; Johler, S. Temporal expression of the staphylococcal enterotoxin D gene under NaCl stress conditions. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Genigeorgis, C.; Foda, M.S.; Mantis, A.; Sadler, W.W. Effect of sodium chloride and pH on enterotoxin C production. Appl. Microbiol. 1971, 21, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Sihto, H.-M.; Budi Susilo, Y.; Tasara, T.; Rådström, P.; Stephan, R.; Schelin, J.; Johler, S. Effect of sodium nitrite and regulatory mutations Δagr, ΔsarA, and ΔsigB on the mRNA and protein levels of staphylococcal enterotoxin D. Food Control 2016, 65, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Schelin, J.; Susilo, Y.; Johler, S. Expression of Staphylococcal Enterotoxins under Stress Encountered during Food Production and Preservation. Toxins (Basel) 2017, 9, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallin-Carlquist, N.; Cao, R.; Márta, D.; Da Silva, A.S.; Schelin, J.; Rådström, P. Acetic acid increases the phage-encoded enterotoxin A expression in Staphylococcus aureus. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef] [Green Version]
- Barber, L.E.; Deibel, R.H. Effect of pH and oxygen tension on staphylococcal growth and enterotoxin formation in fermented sausage. Appl. Microbiol. 1972, 24, 891–898. [Google Scholar] [CrossRef]
- Stewart, C.M. Staphylococcus aureus and Staphylococcal Enterotoxins. In Foodborne Microorganisms of Public Health Significance; Hocking, A.D., Ed.; Australian Institute of Food Science and Technology: North Ryde, Australia, 2003; pp. 359–380. [Google Scholar]
- Rode, T.M.; Møretrø, T.; Langsrud, S.; Langsrud, Ø.; Vogt, G.; Holck, A. Responses of Staphylococcus aureus exposed to HCl and organic acid stress. Can. J. Microbiol. 2010, 56, 777–792. [Google Scholar] [CrossRef]
- Rosengren, Å.; Lindblad, M.; Lindqvist, R. The effect of undissociated lactic acid on Staphylococcus aureus growth and enterotoxin A production. Int. J. Food Microbiol. 2013, 162, 159–166. [Google Scholar] [CrossRef]
- Elahi, S.; Fujikawa, H. Effects of Lactic Acid and Salt on Enterotoxin A Production and Growth of Staphylococcus aureus. J. Food Sci. 2019, 84, 3233–3240. [Google Scholar] [CrossRef]
- Sihto, H.-M.; Tasara, T.; Stephan, R.; Johler, S. Growth behavior and temporal enterotoxin D expression of Staphylococcus aureus strains under glucose and lactic acid stress. Food Control 2016, 62, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Cretenet, M.; Nouaille, S.; Thouin, J.; Rault, L.; Stenz, L.; François, P.; Hennekinne, J.A.; Piot, M.; Maillard, M.B.; Fauquant, J.; et al. Staphylococcus aureus virulence and metabolism are dramatically affected by Lactococcus lactis in cheese matrix. Environ. Microbiol. Rep. 2011, 3, 340–351. [Google Scholar] [CrossRef]
- Wattinger, L.; Stephan, R.; Layer, F.; Johler, S. Comparison of Staphylococcus aureus isolates associated with food intoxication with isolates from human nasal carriers and human infections. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 455–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johler, S.; Layer, F.; Stephan, R. Comparison of virulence and antibiotic resistance genes of food poisoning outbreak isolates of Staphylococcus aureus with isolates obtained from bovine mastitis milk and pig carcasses. J. Food Prot. 2011, 74, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Guinane, C.M.; Zakour, N.L.B.; Tormo-Mas, M.A.; Weinert, L.A.; Lowder, B.V.; Cartwright, R.A.; Smyth, D.S.; Smyth, C.J.; Lindsay, J.A.; Gould, K.A.; et al. Evolutionary genomics of Staphylococcus aureus reveals insights into the origin and molecular basis of ruminant host adaptation. Genome Biol. Evol. 2010, 2, 454–466. [Google Scholar] [CrossRef] [Green Version]
- Sihto, H.M.; Tasara, T.; Stephan, R.; Johler, S. Validation of reference genes for normalization of qPCR mRNA expression levels in Staphylococcus aureus exposed to osmotic and lactic acid stress conditions encountered during food production and preservation. FEMS Microbiol. Lett. 2014, 356, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Poli, M.A.; Rivera, V.R.; Neal, D. Sensitive and specific colorimetric ELISAs for Staphylococcus aureus enterotoxins A and B in urine and buffer. Toxicon 2002, 40, 1723–1726. [Google Scholar] [CrossRef]
- Valihrach, L.; Alibayov, B.; Zdenkova, K.; Demnerova, K. Expression and production of staphylococcal enterotoxin C is substantially reduced in milk. Food Microbiol. 2014, 44, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Valihrach, L.; Alibayov, B.; Demnerova, K. Production of staphylococcal enterotoxin C in milk. Int. Dairy J. 2013, 30, 103–107. [Google Scholar] [CrossRef]
- Zeaki, N.; Cao, R.; Skandamis, P.N.; Rådström, P.; Schelin, J. Assessment of high and low enterotoxin A producing Staphylococcus aureus strains on pork sausage. Int. J. Food Microbiol. 2014, 182–183, 44–50. [Google Scholar] [CrossRef]
- Zeaki, N.; Rådström, P.; Schelin, J. Evaluation of Potential Effects of NaCl and Sorbic Acid on Staphylococcal Enterotoxin A Formation. Microorganisms 2015, 3, 551–566. [Google Scholar] [CrossRef] [Green Version]
- Birbaum-Sihto, H.M. Stress Response in Staphylococcus aureus: Regulatory Mechanisms Influencing Enterotoxin Gene Expression under Stress Relevant to Food Production and Preservation; University of Zurich: Zurich, Switzerland, 2016. [Google Scholar]
- Guillier, L.; Bergis, H.; Guillier, F.; Noel, V.; Auvray, F.; Hennekinne, J.-A. Dose-response Modelling of Staphylococcal Enterotoxins Using Outbreak Data. Procedia Food Sci. 2016, 7, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Domenech, A.; Hernandez, F.J.; Orden, J.A.; Goyache, J.; Lopez, B.; Suarez, G.; Gomez-Lucia, E. Effect of six organic acids on staphylococcal growth and enterotoxin production. Z. Lebensm. Unters. 1992, 194, 124–128. [Google Scholar] [CrossRef]
- Blevins, J.S.; Beenken, K.E.; Elasri, M.O.; Hurlburt, B.K.; Smeltzer, M.S. Strain-dependent differences in the regulatory roles of sarA and agr in Staphylococcus aureus. Infect. Immun. 2002, 70, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassat, J.; Dunman, P.M.; Murphy, E.; Projan, S.J.; Beenken, K.E.; Palm, K.J.; Yang, S.J.; Rice, K.C.; Bayles, K.W.; Smeltzer, M.S. Transcriptional profiling of a Staphylococcus aureus clinical isolate and its isogenic agr and sarA mutants reveals global differences in comparison to the laboratory strain RN6390. Microbiology 2006, 152, 3075–3090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, V.; Smeltzer, M.S.; Elasri, M.O. Genome-scale transcriptional profiling in Staphylococcus aureus: Bringing order out of chaos. FEMS Microbiol. Lett. 2009, 295, 204–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, H.Y.; Ching, W.T.; Stewart, G.C. Regulation of rot expression in Staphylococcus aureus. J. Bacteriol. 2008, 190, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 7, 1–21. [Google Scholar] [CrossRef]
- Weinrick, B.; Dunman, P.M.; McAleese, F.; Murphy, E.; Projan, S.J.; Fang, Y.; Novick, R.P. Effect of Mild Acid on Gene Expression in Staphylococcus aureus. J. Bacteriol. 2004, 186, 8407–8423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starosta, A.L.; Lassak, J.; Jung, K.; Wilson, D.N. The bacterial translation stress response. FEMS Microbiol. Rev. 2014, 38, 1172–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, P.; Tramonti, A.; De Biase, D. Coping with low pH: Molecular strategies in neutralophilic bacteria. FEMS Microbiol. Rev. 2014, 38, 1091–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strain | Protein Variant | Promotor Variant | Origin | Clonal Complex | Reference |
|---|---|---|---|---|---|
| BW10 | SEC2 | secp v1 | SFP | CC45 | 1 |
| NB6 | SEC2 | secp v1 | SFP | CC45 | 2 |
| SAI3 | SEC1 | secp v3 (H-EMRSA-15) | Human infection | CC8 | [49] |
| SAI48 | SEC2 | secp v1 (79_S10) | Human infection | CC5 | [49] |
| SAR1 | SECbovine | secp v2 | bovine mastitis milk | CC151 | [50] |
| SAR38 | SECbovine | secp v2 | bovine mastitis milk | CC151 | [50] |
| OV20 | SECovine | secp v4 | ovine | CC133 | [51] |
| Strain | SEC Produced under pH Stress (ng/mL) | Effect of pH Stress (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 h | 10 h | 24 h | 4 h | 10 h | 24 h | Sum | |||||||
| BW10 | 307 | ± | 81 | 832 | ± | 161 | 740 | ± | 69 | −33 | −75 | −78 | −186 |
| NB6 | 3 | ± | 1 | 9 | ± | 3 | 20 | ± | 16 | −63 | −85 | −64 | −212 |
| SAI3 | 6 | ± | 1 | 20 | ± | 3 | 35 | ± | 11 | 10 | −50 | −69 | −109 |
| SAI48 | 523 | ± | 63 | 4968 | ± | 1641 | 4887 | ± | 1027 | 38 | −17 | −50 | −29 |
| SAR1 | 9 | ± | 4 | 169 | ± | 119 | 132 | ± | 25 | −46 | −21 | −30 | −97 |
| SAR38 | 12 | ± | 2 | 120 | ± | 82 | 143 | ± | 59 | −22 | −47 | −44 | −113 |
| OV20 | 1 | ± | 1 | 41 | ± | 15 | 41 | ± | 14 | −80 | −87 | −88 | −255 |
| SEC produced under control conditions (ng/mL) | |||||||||||||
| BW10 | 462 | ± | 307 | 3325 | ± | 360 | 3410 | ± | 562 | - | - | - | - |
| NB6 | 7 | ± | 3 | 58 | ± | 29 | 54 | ± | 12 | - | - | - | - |
| SAI3 | 6 | ± | 2 | 40 | ± | 10 | 114 | ± | 59 | - | - | - | - |
| SAI48 | 380 | ± | 220 | 5988 | ± | 1317 | 9868 | ± | 4688 | - | - | - | - |
| SAR1 | 16 | ± | 2 | 213 | ± | 87 | 189 | ± | 65 | - | - | - | - |
| SAR38 | 15 | ± | 10 | 228 | ± | 27 | 254 | ± | 7 | - | - | - | - |
| OV20 | 7 | ± | 3 | 328 | ± | 87 | 345 | ± | 34 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etter, D.; Jenni, C.; Tasara, T.; Johler, S. Mild Lactic Acid Stress Causes Strain-Dependent Reduction in SEC Protein Levels. Microorganisms 2021, 9, 1014. https://doi.org/10.3390/microorganisms9051014
Etter D, Jenni C, Tasara T, Johler S. Mild Lactic Acid Stress Causes Strain-Dependent Reduction in SEC Protein Levels. Microorganisms. 2021; 9(5):1014. https://doi.org/10.3390/microorganisms9051014
Chicago/Turabian StyleEtter, Danai, Céline Jenni, Taurai Tasara, and Sophia Johler. 2021. "Mild Lactic Acid Stress Causes Strain-Dependent Reduction in SEC Protein Levels" Microorganisms 9, no. 5: 1014. https://doi.org/10.3390/microorganisms9051014