
Variability, Functions and Interactions of Plant Virus Movement Proteins: What Do We Know So Far?


Abstract
:1. Introduction
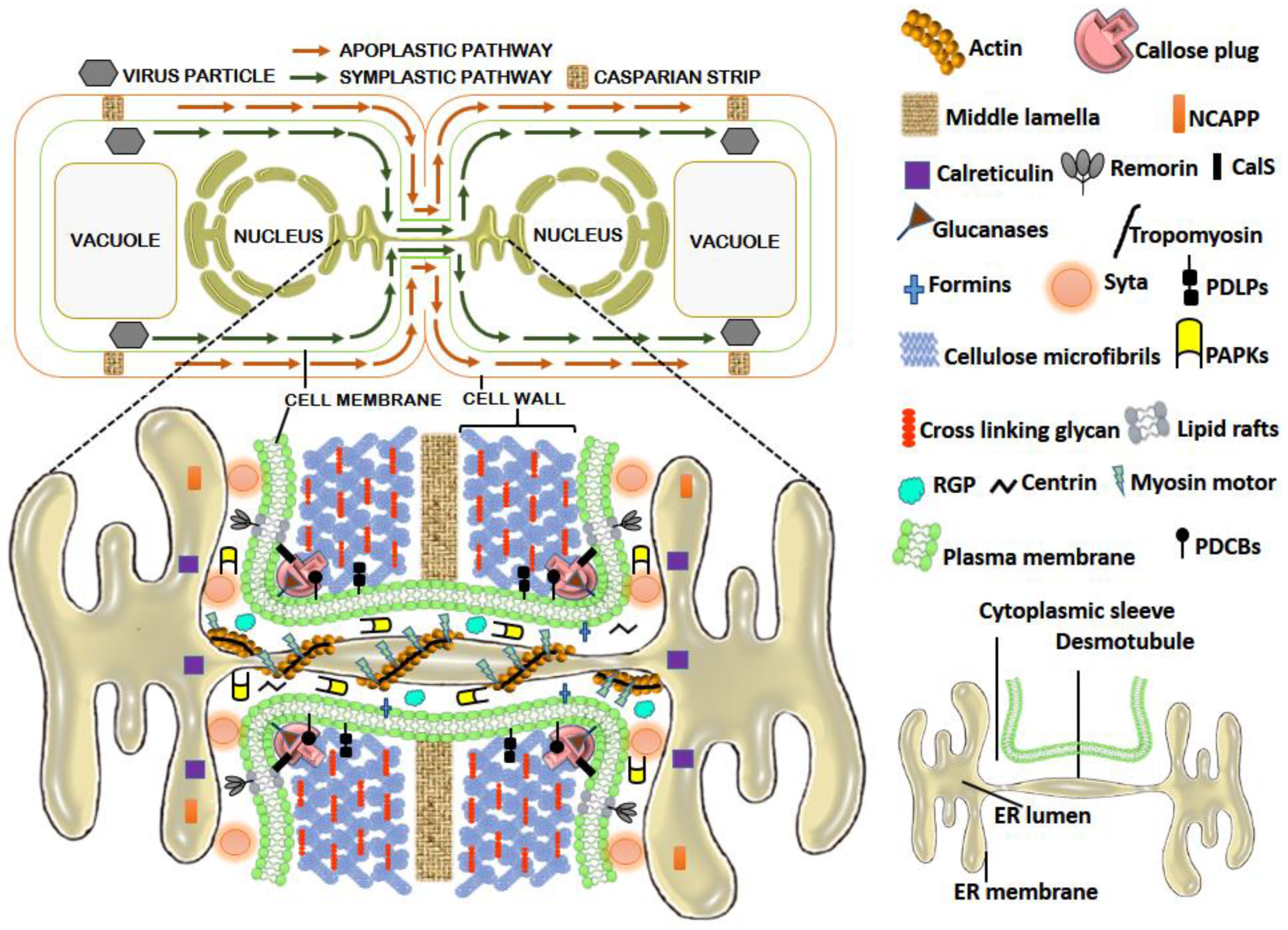
2. PD and PD Associated Proteins (PDAPs): The Facilitators of Virus Movement
3. Types of MPs
3.1. Viral Proteins (MPs) That Increase PD SEL without Any Structural Modifications in PD
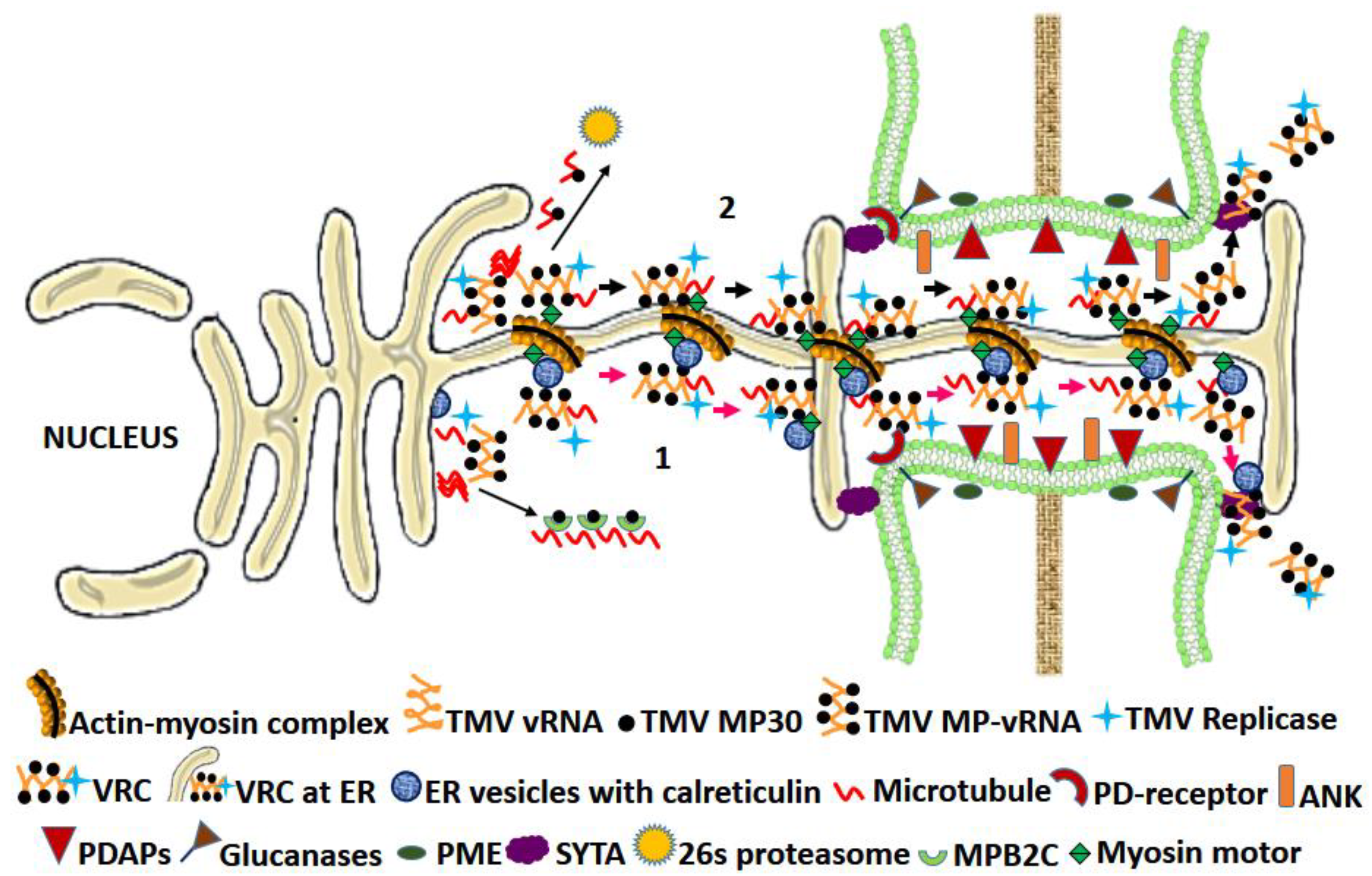
3.1.1. Characteristics of the TMV MP-The “30 K” Superfamily MPs
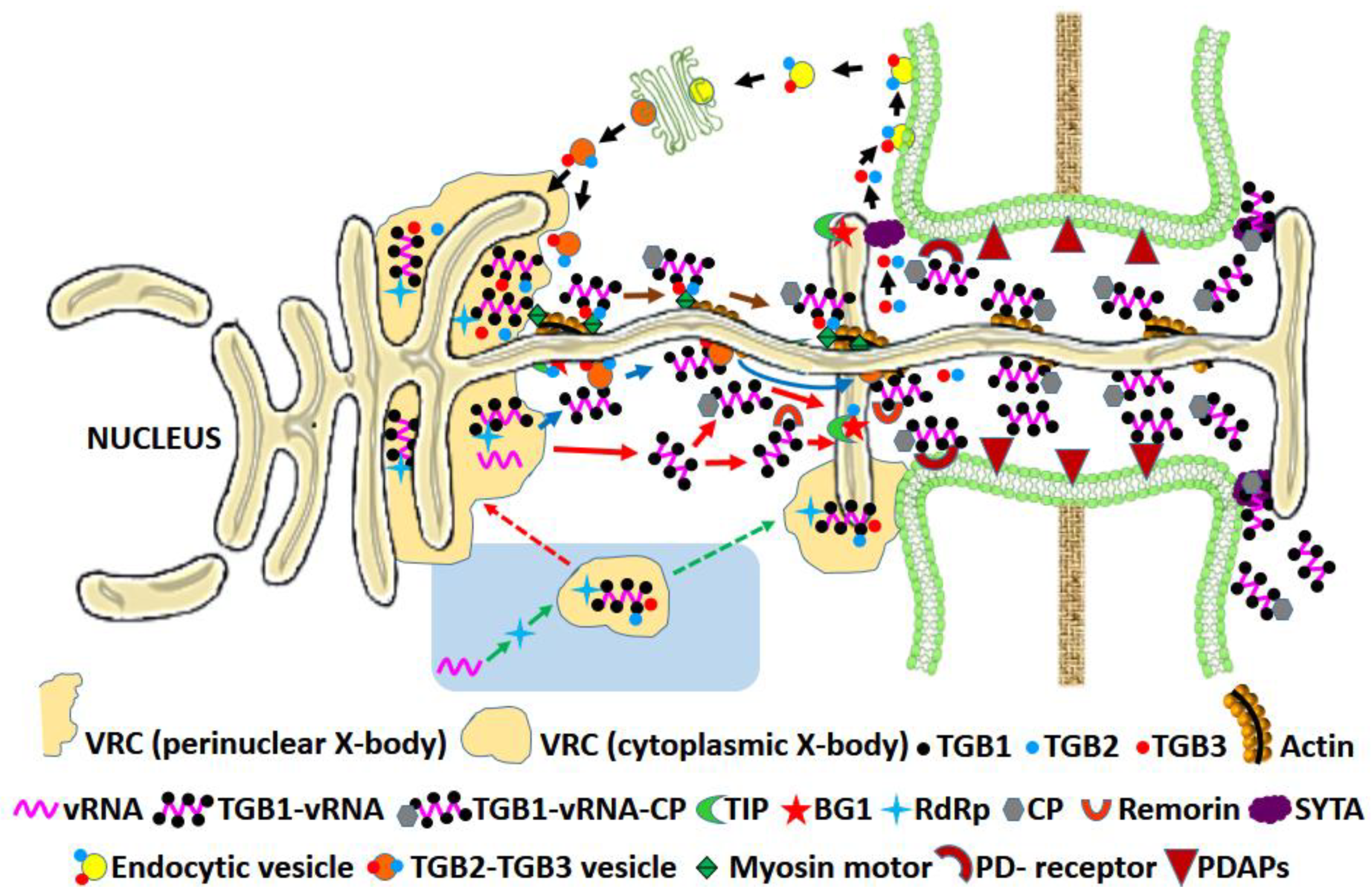
3.1.2. The Triple Gene Block (TGB) Proteins
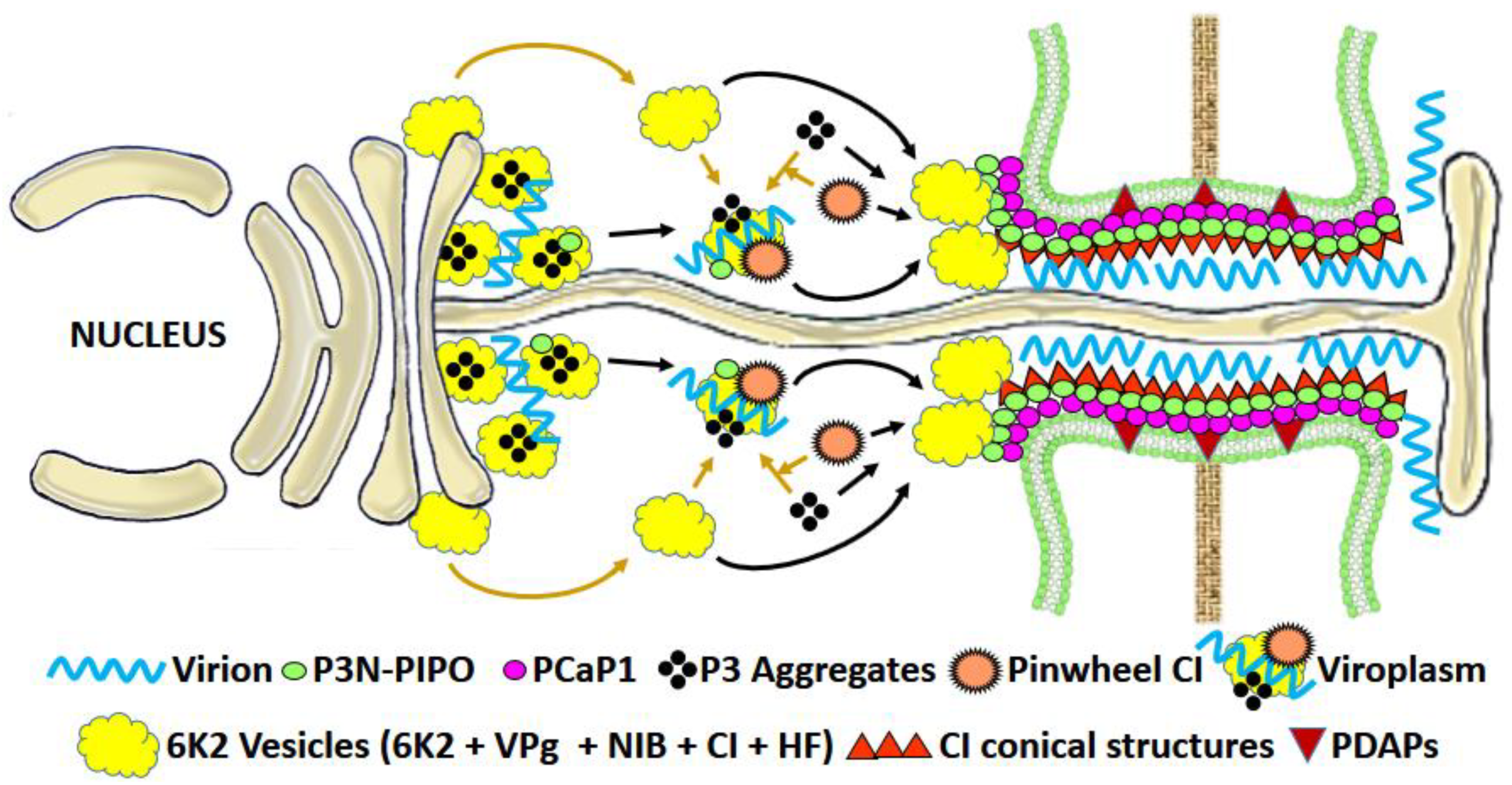
3.1.3. Potyviral MPs
3.2. Viral Proteins (MPs) That Increase PD SEL by Structural Modifications Caused by Tubular Aggregates
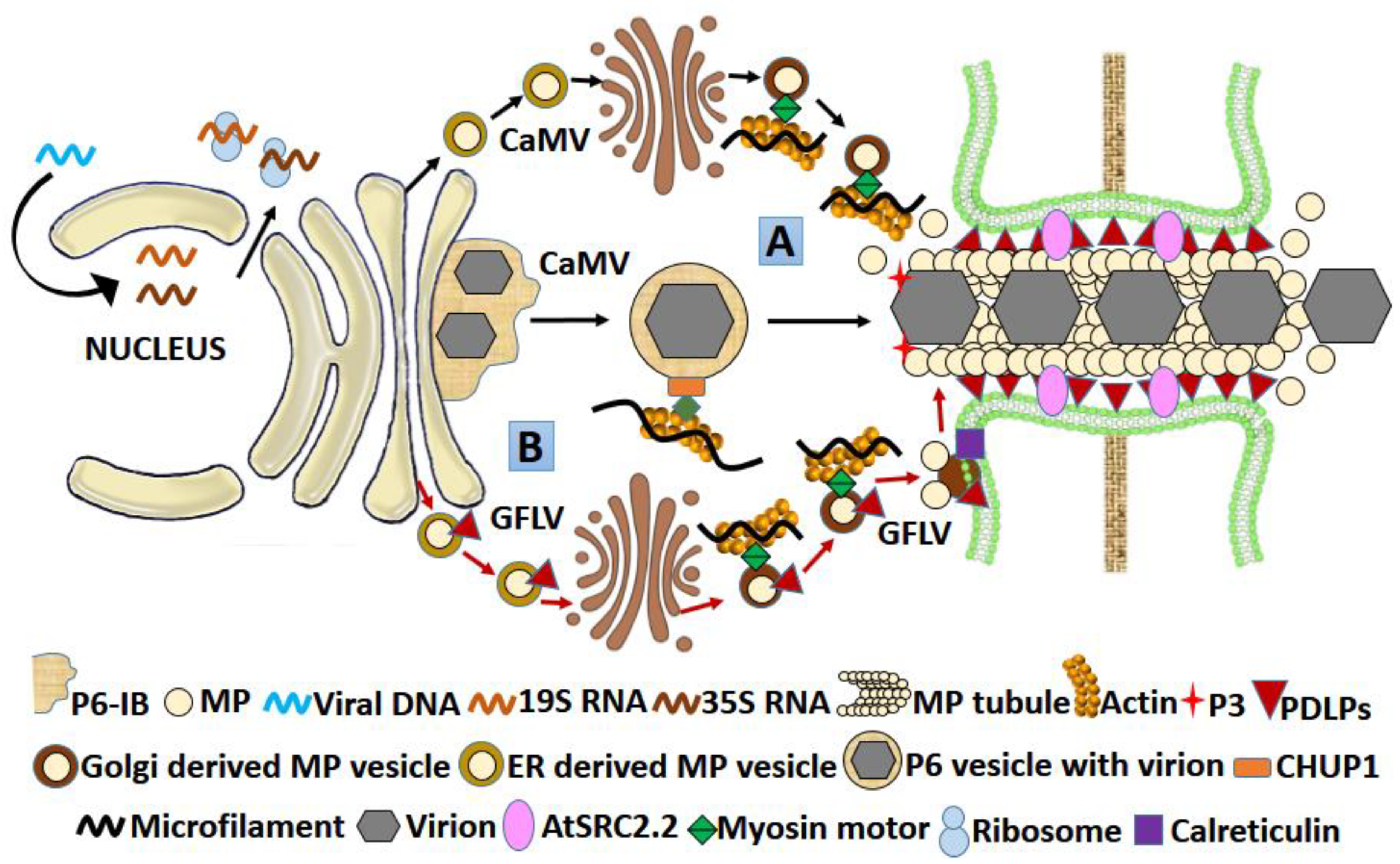
3.2.1. Caulimoviral MP
3.2.2. Tospoviral MP
3.2.3. Umbraviral MP
3.2.4. MPs of Comoviruses, Fabaviruses, and Nepoviruses
3.3. MPs That Increase PD SEL with or without Any Structural Modifications across PD
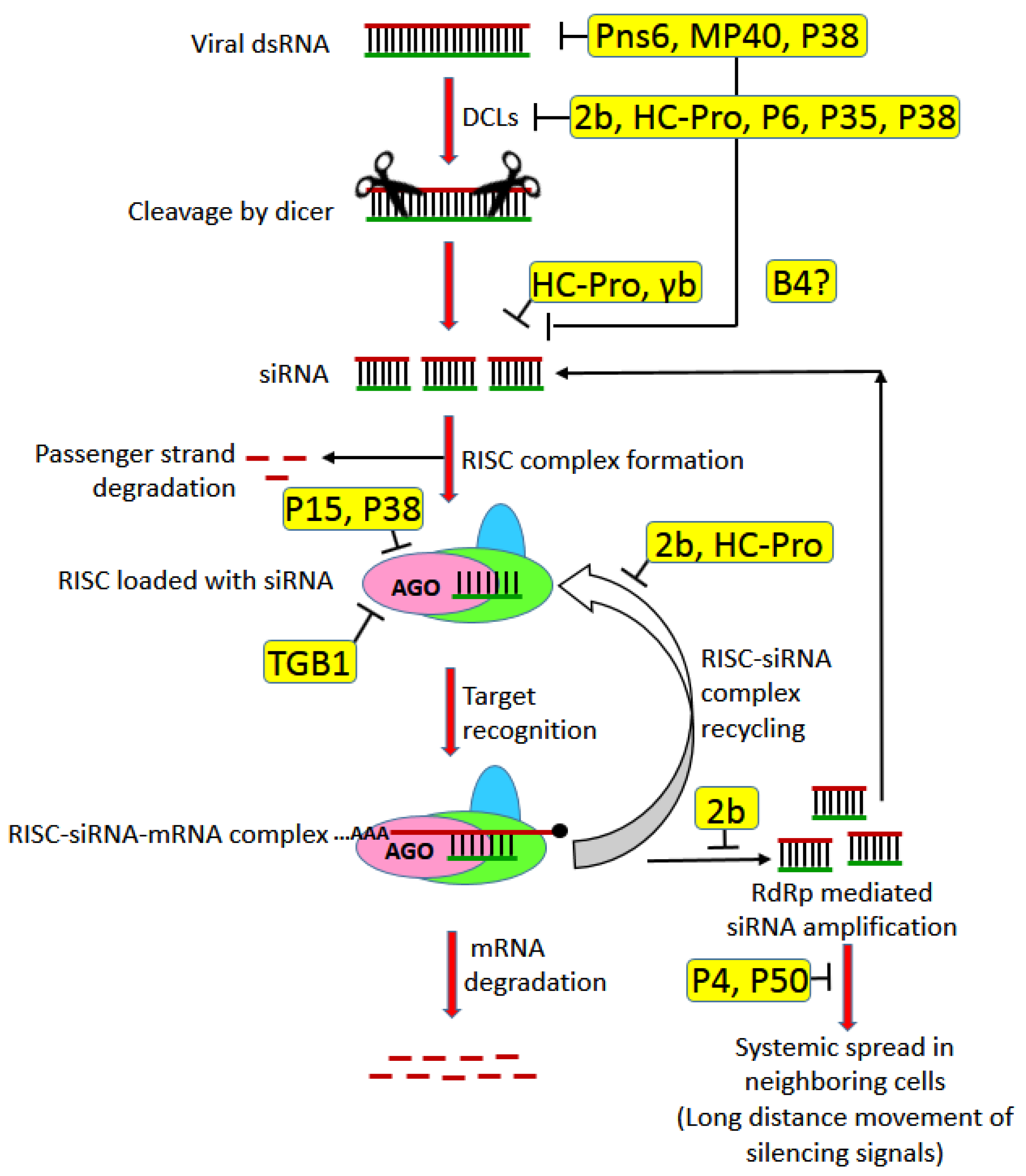
3.4. MPs as RNA Silencing Suppressors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navarro, J.A.; Sanchez-Navarro, J.A.; Pallas, V. Key checkpoints in the movement of plant viruses through the host. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 104, pp. 1–64. [Google Scholar]
- Kappagantu, M.; Collum, T.D.; Dardick, C.; Culver, J.N. Viral Hacks of the Plant Vasculature: The Role of Phloem Alterations in Systemic Virus Infection. Annu. Rev. Virol. 2020, 7, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Terry, B.R.; Robards, A.W. Hydrodynamic radius alone governs the mobility of molecules through plasmodesmata. Planta 1987, 171, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Liarzi, O.; Epel, B.L. Development of a quantitative tool for measuring changes in the coefficient of conductivity of plasmodesmata induced by developmental, biotic, and abiotic signals. Protoplasma 2005, 225, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Lucas, W.J. Plant viral movement proteins: Agents for cell-to-cell trafficking of viral genomes. Virology 2006, 344, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.-S.; Ju, H.-J. The plant cellular systems for plant virus movement. Plant Pathol. J. 2017, 33, 213. [Google Scholar] [CrossRef]
- Schoelz, J.E.; Angel, C.A.; Nelson, R.S.; Leisner, S.M. A model for intracellular movement of Cauliflower mosaic virus: The concept of the mobile virion factory. J. Exp. Bot. 2016, 67, 2039–2048. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Kumar, R.; Hyun, T.K.; Kim, J.-Y. Cell-to-cell movement of viruses via plasmodesmata. J. Plant Res. 2015, 128, 37–47. [Google Scholar] [CrossRef]
- Heinlein, M. Plant virus replication and movement. Virology 2015, 479, 657–671. [Google Scholar] [CrossRef] [Green Version]
- Pitzalis, N.; Heinlein, M. The roles of membranes and associated cytoskeleton in plant virus replication and cell-to-cell movement. J. Exp. Bot. 2018, 69, 117–132. [Google Scholar] [CrossRef]
- Reagan, B.C.; Burch-Smith, T.M. Viruses reveal the secrets of plasmodesmal cell biology. Mol. Plant-Microbe Interact. 2020, 33, 26–39. [Google Scholar] [CrossRef]
- Morozov, S.Y.; Solovyev, A.G. Small hydrophobic viral proteins involved in intercellular movement of diverse plant virus genomes. Aims Microbiol. 2020, 6, 305. [Google Scholar] [CrossRef]
- Levy, A.; Tilsner, J. Creating contacts between replication and movement at plasmodesmata–a role for membrane contact sites in plant virus infections? Front. Plant Sci. 2020, 11, 862. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cheng, X. Intercellular movement of plant RNA viruses: Targeting replication complexes to the plasmodesma for both accuracy and efficiency. Traffic 2020, 21, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Deom, C.M.; Lapidot, M.; Beachy, R.N. Plant virus movement proteins. Cell 1992, 69, 221–224. [Google Scholar] [CrossRef]
- Kellmann, J.-W. Identification of plant virus movement-host protein interactions. Zeitschrift Für Naturforschung C 2001, 56, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Folimonova, S.Y.; Tilsner, J. Hitchhikers, highway tolls and roadworks: The interactions of plant viruses with the phloem. Curr. Opin. Plant Biol. 2018, 43, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Verchot-Lubicz, J.; Torrance, L.; Solovyev, A.G.; Morozov, S.Y.; Jackson, A.O.; Gilmer, D. Varied movement strategies employed by triple gene block–encoding viruses. Mol. Plant-Microbe Interact. 2010, 23, 1231–1247. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, W.J.; Grison, M.S.; Bayer, E.M. Shaping intercellular channels of plasmodesmata: The structure-to-function missing link. J. Exp. Bot. 2018, 69, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Dorokhov, Y.L.; Ershova, N.M.; Sheshukova, E.V.; Komarova, T.V. Plasmodesmata Conductivity Regulation: A Mechanistic Model. Plants 2019, 8, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radford, J.E.; White, R.G. Localization of a myosin-like protein to plasmodesmata. Plant J. 1998, 14, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Zavaliev, R.; Levy, A.; Gera, A.; Epel, B.L. Subcellular dynamics and role of Arabidopsis β-1, 3-glucanases in cell-to-cell movement of tobamoviruses. Mol. Plant-Microbe Interact. 2013, 26, 1016–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.-H.; Tian, G.-W.; Gafni, Y.; Citovsky, V. Effects of calreticulin on viral cell-to-cell movement. Plant Physiol. 2005, 138, 1866–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baluška, F.; Šamaj, J.; Napier, R.; Volkmann, D. Maize calreticulin localizes preferentially to plasmodesmata in root apex. Plant J. 1999, 19, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Diao, M.; Ren, S.; Wang, Q.; Qian, L.; Shen, J.; Liu, Y.; Huang, S. Arabidopsis formin 2 regulates cell-to-cell trafficking by capping and stabilizing actin filaments at plasmodesmata. Elife 2018, 7, e36316. [Google Scholar] [CrossRef]
- Iswanto, A.B.B.; Kim, J.-Y. Lipid raft, regulator of plasmodesmal callose homeostasis. Plants 2017, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Yoo, B.-C.; Rojas, M.R.; Gomez-Ospina, N.; Staehelin, L.A.; Lucas, W.J. Selective trafficking of non-cell-autonomous proteins mediated by NtNCAPP1. Science 2003, 299, 392–396. [Google Scholar] [CrossRef]
- Simpson, C.; Thomas, C.; Findlay, K.; Bayer, E.; Maule, A.J. An Arabidopsis GPI-anchor plasmodesmal neck protein with callose binding activity and potential to regulate cell-to-cell trafficking. Plant Cell 2009, 21, 581–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, G.-H.; Shine, M.B.; de Lorenzo, L.; Yu, K.; Cui, W.; Navarre, D.; Hunt, A.G.; Lee, J.-Y.; Kachroo, A.; Kachroo, P. Plasmodesmata localizing proteins regulate transport and signaling during systemic acquired immunity in plants. Cell Host Microbe 2016, 19, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Raffaele, S.; Bayer, E.; Lafarge, D.; Cluzet, S.; Retana, S.G.; Boubekeur, T.; Leborgne-Castel, N.; Carde, J.-P.; Lherminier, J.; Noirot, E. Remorin, a solanaceae protein resident in membrane rafts and plasmodesmata, impairs potato virus X movement. Plant Cell 2009, 21, 1541–1555. [Google Scholar] [CrossRef] [Green Version]
- Burch-Smith, T.M.; Cui, Y.; Zambryski, P.C. Reduced levels of class 1 reversibly glycosylated polypeptide increase intercellular transport via plasmodesmata. Plant Signal. Behav. 2012, 7, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Lazarowitz, S.G.; Citovsky, V. The plasmodesmal localization signal of TMV MP is recognized by plant synaptotagmin SYTA. MBio 2018, 9, e01314-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, M.; Huang, S. An Update on the Role of the Actin Cytoskeleton in Plasmodesmata: A Focus on Formins. Front. Plant Sci. 2021, 12, 191. [Google Scholar] [CrossRef] [PubMed]
- Nishiguchi, M.; Motoyoshi, F.; Oshima, N. Behaviour of a temperature sensitive strain of tobacco mosaic virus in tomato leaves and protoplasts. J. Gen. Virol. 1978, 39, 53–61. [Google Scholar] [CrossRef]
- Wolf, S.; Lucas, W.J.; Deom, C.M.; Beachy, R.N. Movement protein of tobacco mosaic virus modifies plasmodesmatal size exclusion limit. Science 1989, 246, 377–379. [Google Scholar] [CrossRef]
- Ueki, S.; Spektor, R.; Natale, D.M.; Citovsky, V. ANK, a host cytoplasmic receptor for the Tobacco mosaic virus cell-to-cell movement protein, facilitates intercellular transport through plasmodesmata. PLoS Pathog. 2010, 6, e1001201. [Google Scholar] [CrossRef] [PubMed]
- Melcher, U. The ‘30K’superfamily of viral movement proteins. J. Gen. Virol. 2000, 81, 257–266. [Google Scholar] [PubMed]
- Moore, P.J.; Fenczik, C.A.; Deom, C.M.; Beachy, R.N. Developmental changes in plasmodesmata in transgenic tobacco expressing the movement protein of tobacco mosaic virus. Protoplasma 1992, 170, 115–127. [Google Scholar] [CrossRef]
- Beachy, R.N.; Heinlein, M. Role of P30 in replication and spread of TMV. Traffic 2000, 1, 540–544. [Google Scholar] [CrossRef]
- Peiró, A.; Martínez-Gil, L.; Tamborero, S.; Pallás, V.; Sánchez-Navarro, J.A.; Mingarro, I. The Tobacco mosaic virus movement protein associates with but does not integrate into biological membranes. J. Virol. 2014, 88, 3016–3026. [Google Scholar] [CrossRef] [Green Version]
- Guenoune-Gelbart, D.; Elbaum, M.; Sagi, G.; Levy, A.; Epel, B.L. Tobacco mosaic virus (TMV) replicase and movement protein function synergistically in facilitating TMV spread by lateral diffusion in the plasmodesmal desmotubule of Nicotiana benthamiana. Mol. Plant-Microbe Interact. 2008, 21, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, S.; Watanabe, Y.; Beachy, R.N. Tobacco mosaic virus infection spreads cell to cell as intact replication complexes. Proc. Natl. Acad. Sci. USA 2004, 101, 6291–6296. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, Y.; Hanazawa, K.; Yoshioka, K.; Oguchi, T.; Kawakami, S.; Watanabe, Y.; Nishiguchi, M.; Nyunoya, H. In vitro phosphorylation of the movement protein of tomato mosaic tobamovirus by a cellular kinase. J. Gen. Virol. 2000, 81, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, Y.; Ohshima, M.; Yoshioka, K.; Nishiguchi, M.; Nyunoya, H. The catalytic subunit of protein kinase CK2 phosphorylates in vitro the movement protein of Tomato mosaic virus. J. Gen. Virol. 2003, 84, 497–505. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Taoka, K.; Yoo, B.-C.; Ben-Nissan, G.; Kim, D.-J.; Lucas, W.J. Plasmodesmal-associated protein kinase in tobacco and Arabidopsis recognizes a subset of non-cell-autonomous proteins. Plant Cell 2005, 17, 2817–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trutnyeva, K.; Bachmaier, R.; Waigmann, E. Mimicking carboxyterminal phosphorylation differentially effects subcellular distribution and cell-to-cell movement of Tobacco mosaic virus movement protein. Virology 2005, 332, 563–577. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Lazarowitz, S.G.; Citovsky, V. Identification of a functional plasmodesmal localization signal in a plant viral cell-to-cell-movement protein. MBio 2016, 7, e02052-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Huang, C.; Zeng, J.; Yu, H.; Li, Y.; Yuan, C. Identification of two additional plasmodesmata localization domains in the tobacco mosaic virus cell-to-cell-movement protein. Biochem. Biophys. Res. Commun. 2020, 521, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Zheng, J.Y.; Lazarowitz, S.G. Synaptotagmin SYTA forms ER-plasma membrane junctions that are recruited to plasmodesmata for plant virus movement. Curr. Biol. 2015, 25, 2018–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, K.; Tamura, K.; Fukao, Y.; Shimada, T. Structural and functional relationships between plasmodesmata and plant endoplasmic reticulum–plasma membrane contact sites consisting of three synaptotagmins. New Phytol. 2020, 226, 798–808. [Google Scholar] [CrossRef]
- Harries, P.A.; Park, J.-W.; Sasaki, N.; Ballard, K.D.; Maule, A.J.; Nelson, R.S. Differing requirements for actin and myosin by plant viruses for sustained intercellular movement. Proc. Natl. Acad. Sci. USA 2009, 106, 17594–17599. [Google Scholar] [CrossRef] [Green Version]
- Amsbury, S.; Kirk, P.; Benitez-Alfonso, Y. Emerging models on the regulation of intercellular transport by plasmodesmata-associated callose. J. Exp. Bot. 2018, 69, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Sheng, J.; Hind, G.; Handa, A.K.; Citovsky, V. Interaction between the tobacco mosaic virus movement protein and host cell pectin methylesterases is required for viral cell-to-cell movement. Embo J. 2000, 19, 913–920. [Google Scholar] [CrossRef]
- Chen, M.; Citovsky, V. Systemic movement of a tobamovirus requires host cell pectin methylesterase. Plant J. 2003, 35, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Oparka, K.J.; Prior, D.A.M.; Cruz, S.S.; Padgett, H.S.; Beachy, R.N. Gating of epidermal plasmodesmata is restricted to the leading edge of expanding infection sites of tobacco mosaic virus (TMV). Plant J. 1997, 12, 781–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komarova, T.V.; Skulachev, M.V.; Ivanov, P.A.; Klyushin, A.G.; Dorokhov, Y.L.; Atabekov, J.G. Internal ribosome entry site from crucifer tobamovirus promotes initiation of translation in Escherichia coli. Dokl. Biochem. Biophys. 2003, 389, 118–121. [Google Scholar] [CrossRef]
- Zvereva, S.D.; Ivanov, P.A.; Skulachev, M.V.; Klyushin, A.G.; Dorokhov, Y.L.; Atabekov, J.G. Evidence for contribution of an internal ribosome entry site to intercellular transport of a tobamovirus. J. Gen. Virol. 2004, 85, 1739–1744. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Ivanov, P.A.; Komarova, T.V.; Skulachev, M.V.; Atabekov, J.G. An internal ribosome entry site located upstream of the crucifer-infecting tobamovirus coat protein (CP) gene can be used for CP synthesis in vivo. J. Gen. Virol. 2006, 87, 2693–2697. [Google Scholar] [CrossRef] [PubMed]
- Vogler, H.; Kwon, M.-O.; Dang, V.; Sambade, A.; Fasler, M.; Ashby, J.; Heinlein, M. Tobacco mosaic virus movement protein enhances the spread of RNA silencing. PLoS Pathog. 2008, 4, e1000038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.; Kim, K.H.; Giesman-Cookmeyer, D.; Lommel, S.A. The roles of the red clover necrotic mosaic virus capsid and cell-to-cell movement proteins in systemic infection. Virology 1993, 192, 27–32. [Google Scholar] [CrossRef]
- Tremblay, D.; Vaewhongs, A.A.; Turner, K.A.; Sit, T.L.; Lommel, S.A. Cell wall localization of Red clover necrotic mosaic virus movement protein is required for cell-to-cell movement. Virology 2005, 333, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, J.G.; Sit, T.L.; Heinsohn, C.; George, C.G.; Kim, K.-H.; Lommel, S.A. The Red clover necrotic mosaic virus RNA-2 encoded movement protein is a second suppressor of RNA silencing. Virology 2008, 381, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Kaido, M.; Tsuno, Y.; Mise, K.; Okuno, T. Endoplasmic reticulum targeting of the Red clover necrotic mosaic virus movement protein is associated with the replication of viral RNA1 but not that of RNA2. Virology 2009, 395, 232–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaido, M.; Funatsu, N.; Tsuno, Y.; Mise, K.; Okuno, T. Viral cell-to-cell movement requires formation of cortical punctate structures containing Red clover necrotic mosaic virus movement protein. Virology 2011, 413, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Ye, X.; Willie, K.; Lin, J.; Zhang, X.; Redinbaugh, M.G.; Simon, A.E.; Morris, T.J.; Qu, F. The capsid protein of Turnip crinkle virus overcomes two separate defense barriers to facilitate systemic movement of the virus in Arabidopsis. J. Virol. 2010, 84, 7793–7802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Gil, L.; Johnson, A.E.; Mingarro, I. Membrane insertion and biogenesis of the Turnip crinkle virus p9 movement protein. J. Virol. 2010, 84, 5520–5527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilar, M.; Saurí, A.; Marcos, J.F.; Mingarro, I.; Pérez-Payá, E. Transient structural ordering of the RNA-binding domain of carnation mottle virus p7 movement protein modulates nucleic acid binding. Chembiochem 2005, 6, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Turiño, S.; Hernández, C. A membrane-associated movement protein of Pelargonium flower break virus shows RNA-binding activity and contains a biologically relevant leucine zipper-like motif. Virology 2011, 413, 310–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genovés, A.; Navarro, J.A.; Pallás, V. The intra-and intercellular movement of Melon necrotic spot virus (MNSV) depends on an active secretory pathway. Mol. Plant-Microbe Interact. 2010, 23, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Serra-Soriano, M.; Antonio Navarro, J.; Pallás, V. Dissecting the multifunctional role of the N-terminal domain of the Melon necrotic spot virus coat protein in RNA packaging, viral movement and interference with antiviral plant defence. Mol. Plant Pathol. 2017, 18, 837–849. [Google Scholar] [CrossRef]
- Lazareva, E.A.; Lezzhov, A.A.; Komarova, T.V.; Morozov, S.Y.; Heinlein, M.; Solovyev, A.G. A novel block of plant virus movement genes. Mol. Plant Pathol. 2017, 18, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Lazareva, E.A.; Lezzhov, A.A.; Chergintsev, D.A.; Golyshev, S.A.; Dolja, V.V.; Morozov, S.Y.; Heinlein, M.; Solovyev, A.G. Reticulon-like properties of a plant virus-encoded movement protein. New Phytol. 2021, 229, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Park, J.-W.; Scholthof, H.B. Separate regions on the tomato bushy stunt virus p22 protein mediate cell-to-cell movement versus elicitation of effective resistance responses. Mol. Plant-Microbe Interact. 1999, 12, 285–292. [Google Scholar] [CrossRef]
- Agranovsky, A.A. Closteroviruses: Molecular biology, evolution and interactions with cells. In Plant Viruses: Evolution and Management; Springer: Singapore, 2016; pp. 231–252. [Google Scholar]
- Qiao, W.; Medina, V.; Kuo, Y.-W.; Falk, B.W. A distinct, non-virion plant virus movement protein encoded by a crinivirus essential for systemic infection. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Yaghmaiean, H.; Wu, G.; Wu, X.; Chen, X.; Thorn, G.; Wang, A. The C-terminal region of the Turnip mosaic virus P3 protein is essential for viral infection via targeting P3 to the viral replication complex. Virology 2017, 510, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; He, R.; Bernards, M.A.; Wang, A. The cis-expression of the coat protein of turnip mosaic virus is essential for viral intercellular movement in plants. Mol. Plant Pathol. 2020, 21, 1194–1211. [Google Scholar] [CrossRef]
- Rojas, M.R.; Zerbini, F.M.; Allison, R.F.; Gilbertson, R.L.; Lucas, W.J. Capsid protein and helper component-proteinase function as potyvirus cell-to-cell movement proteins. Virology 1997, 237, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Yang, Z.; Zhang, H.; Zhang, J.; Xu, J. Remorin interacting with PCaP1 impairs Turnip mosaic virus intercellular movement but is antagonised by VPg. New Phytol. 2020, 225, 2122–2139. [Google Scholar] [CrossRef] [PubMed]
- Chai, M.; Wu, X.; Liu, J.; Fang, Y.; Luan, Y.; Cui, X.; Zhou, X.; Wang, A.; Cheng, X. P3N-PIPO interacts with P3 via the shared N-terminal domain to recruit viral replication vesicles for cell-to-cell movement. J. Virol. 2020, 94, e01898-19. [Google Scholar] [CrossRef]
- Lee, L.; Palukaitis, P.; Gray, S.M. Host-dependent requirement for the Potato leafroll virus 17-kda protein in virus movement. Mol. Plant-Microbe Interact. 2002, 15, 1086–1094. [Google Scholar] [CrossRef]
- DeBlasio, S.L.; Xu, Y.; Johnson, R.S.; Rebelo, A.R.; MacCoss, M.J.; Gray, S.M.; Heck, M. The interaction dynamics of two potato leafroll virus movement proteins affects their localization to the outer membranes of mitochondria and plastids. Viruses 2018, 10, 585. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Su, X.; Luo, X.; Zhang, Y.; Zhang, D.; Du, J.; Zhang, Z.; OuYang, X.; Zhang, S.; Liu, Y. First evidence showing that Pepper vein yellows virus P4 protein is a movement protein. BMC Microbiol. 2020, 20, 1–6. [Google Scholar] [CrossRef]
- Fusaro, A.F.; Barton, D.A.; Nakasugi, K.; Jackson, C.; Kalischuk, M.L.; Kawchuk, L.M.; Vaslin, M.F.S.; Correa, R.L.; Waterhouse, P.M. The luteovirus P4 movement protein is a suppressor of systemic RNA silencing. Viruses 2017, 9, 294. [Google Scholar] [CrossRef] [Green Version]
- Kleinow, T.; Happle, A.; Kober, S.; Linzmeier, L.; Rehm, T.M.; Fritze, J.; Buchholz, P.C.F.; Kepp, G.; Jeske, H.; Wege, C. Phosphorylations of the Abutilon Mosaic Virus Movement Protein Affect Its Self-Interaction, Symptom Development, Viral DNA Accumulation, and Host Range. Front. Plant Sci. 2020, 11, 1155. [Google Scholar] [CrossRef]
- Lee, C.; Zheng, Y.; Chan, C.; Ku, H.; Chang, C.; Jan, F. A single amino acid substitution in the movement protein enables the mechanical transmission of a geminivirus. Mol. Plant Pathol. 2020, 21, 571–588. [Google Scholar] [CrossRef] [Green Version]
- Mann, K.S.; Bejerman, N.; Johnson, K.N.; Dietzgen, R.G. Cytorhabdovirus P3 genes encode 30K-like cell-to-cell movement proteins. Virology 2016, 489, 20–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Lin, W.; Sun, K.; Wang, S.; Zhou, X.; Jackson, A.O.; Li, Z. Specificity of plant rhabdovirus cell-to-cell movement. J. Virol. 2019, 93, e00296-19. [Google Scholar] [CrossRef] [Green Version]
- Solovyev, A.; Kalinina, N.; Morozov, S. Recent advances in research of plant virus movement mediated by triple gene block. Front. Plant Sci. 2012, 3, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.Y.; Yan, L.; Gorter, F.A.; Kim, B.Y.T.; Cui, Y.; Hu, Y.; Yuan, C.; Grindheim, J.; Ganesan, U.; Liu, Z. Brachypodium distachyon line Bd3-1 resistance is elicited by the barley stripe mosaic virus triple gene block 1 movement protein. J. Gen. Virol. 2012, 93, 2729–2739. [Google Scholar] [CrossRef] [PubMed]
- Perraki, A.; Binaghi, M.; Mecchia, M.A.; Gronnier, J.; German-Retana, S.; Mongrand, S.; Bayer, E.; Zelada, A.M.; Germain, V. StRemorin1. 3 hampers Potato virus X TGBp1 ability to increase plasmodesmata permeability, but does not interfere with its silencing suppressor activity. FEBS Lett. 2014, 588, 1699–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathioudakis, M.M.; Veiga, R.S.L.; Canto, T.; Medina, V.; Mossialos, D.; Makris, A.M.; Livieratos, I. P epino mosaic virus triple gene block protein 1 (TGBp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol. Plant Pathol. 2013, 14, 589–601. [Google Scholar] [CrossRef]
- Wu, X.; Liu, J.; Chai, M.; Wang, J.; Li, D.; Wang, A.; Cheng, X. The Potato virus X TGBp2 protein plays dual functional roles in viral replication and movement. J. Virol. 2019, 93, e01635-18. [Google Scholar] [CrossRef] [Green Version]
- Mann, K.; Meng, B. The triple gene block movement proteins of a grape virus in the genus Foveavirus confer limited cell-to-cell spread of a mutant Potato virus X. Virus Genes 2013, 47, 93–104. [Google Scholar] [CrossRef] [PubMed]
- May, J.P.; Johnson, P.Z.; Ilyas, M.; Gao, F.; Simon, A.E. The Multifunctional Long-Distance Movement Protein of Pea Enation Mosaic Virus 2 Protects Viral and Host Transcripts from Nonsense-Mediated Decay. Mbio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Kasteel, D.T.J.; Perbal, M.-C.; Boyer, J.-C.; Wellink, J.; Goldbach, R.W.; Maule, A.J.; Van Lent, J.W.M. The movement proteins of cowpea mosaic virus and cauliflower mosaic virus induce tubular structures in plant and insect cells. J. Gen. Virol. 1996, 77, 2857–2864. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.L.; Perbal, C.; Maule, A.J. A mutation of cauliflower mosaic virus gene I interferes with virus movement but not virus replication. Virology 1993, 192, 415–421. [Google Scholar] [CrossRef]
- Amari, K.; Boutant, E.; Hofmann, C.; Schmitt-Keichinger, C.; Fernandez-Calvino, L.; Didier, P.; Lerich, A.; Mutterer, J.; Thomas, C.L.; Heinlein, M. A family of plasmodesmal proteins with receptor-like properties for plant viral movement proteins. PLoS Pathog. 2010, 6, e1001119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, E.W.; Lauritis, J.A. Plant virions in plasmodesmata. Virology 1969, 37, 681–685. [Google Scholar] [CrossRef]
- Ritzenthaler, C.; Schmit, A.C.; Michler, P.; Stussi-Garaud, C.; Pinck, L. Grapevine fanleaf nepovirus P38 putative movement protein is located on tubules in vivo. Mol. Plant-Microbe Interact. 1995, 8, 379–387. [Google Scholar] [CrossRef]
- Laporte, C.; Vetter, G.; Loudes, A.-M.; Robinson, D.G.; Hillmer, S.; Stussi-Garaud, C.; Ritzenthaler, C. Involvement of the secretory pathway and the cytoskeleton in intracellular targeting and tubule assembly of Grapevine fanleaf virus movement protein in tobacco BY-2 cells. Plant Cell 2003, 15, 2058–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belin, C.; Schmitt, C.; Gaire, F.; Walter, B.; Demangeat, G.; Pinck, L. The nine C-terminal residues of the grapevine fanleaf nepovirus movement protein are critical for systemic virus spread. J. Gen. Virol. 1999, 80, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Lekkerkerker, A.; Wellink, J.; Yuan, P.; Van Lent, J.; Goldbach, R.; Van Kammen, A.B. Distinct functional domains in the cowpea mosaic virus movement protein. J. Virol. 1996, 70, 5658–5661. [Google Scholar] [CrossRef] [Green Version]
- Bertens, P.; Wellink, J.; Goldbach, R.; van Kammen, A. Mutational analysis of the cowpea mosaic virus movement protein. Virology 2000, 267, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Bertens, P.; Heijne, W.; Van der Wel, N.; Wellink, J.; Van Kammen, A. Studies on the C-terminus of the Cowpea mosaic virus movement protein. Arch. Virol. 2003, 148, 265–279. [Google Scholar] [CrossRef]
- Pouwels, J.; Kornet, N.; van Bers, N.; Guighelaar, T.; van Lent, J.; Bisseling, T.; Wellink, J. Identification of distinct steps during tubule formation by the movement protein of Cowpea mosaic virus. J. Gen. Virol. 2003, 84, 3485–3494. [Google Scholar] [CrossRef]
- Pouwels, J.; van der Velden, T.; Willemse, J.; Borst, J.W.; van Lent, J.; Bisseling, T.; Wellink, J. Studies on the origin and structure of tubules made by the movement protein of Cowpea mosaic virus. J. Gen. Virol. 2004, 85, 3787–3796. [Google Scholar] [CrossRef]
- Storms, M.M.H.; Kromelink, R.; Peters, D.; Van Lent, J.W.M.; Goldbach, R.O.B.W. The nonstructural NSm protein of tomato spotted wilt virus induces tubular structures in plant and insect cells. Virology 1995, 214, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ye, L.; Lang, G.; Zhang, C.; Hong, J.; Zhou, X. The VP37 protein of Broad bean wilt virus 2 induces tubule-like structures in both plant and insect cells. Virus Res. 2011, 155, 42–47. [Google Scholar] [CrossRef]
- Wijkamp, I.; van Lent, J.; Kormelink, R.; Goldbach, R.; Peters, D. Multiplication of tomato spotted wilt virus in its insect vector, Frankliniella occidentalis. J. Gen. Virol. 1993, 74, 341–349. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Oparka, K.J.; Santa Cruz, S.; Robinson, D.J.; Taliansky, M.E. Intracellular location of two groundnut rosette umbravirus proteins delivered by PVX and TMV vectors. Virology 1998, 242, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Nurkiyanova, K.M.; Ryabov, E.V.; Kalinina, N.O.; Fan, Y.; Andreev, I.; Fitzgerald, A.G.; Palukaitis, P.; Taliansky, M. Umbravirus-encoded movement protein induces tubule formation on the surface of protoplasts and binds RNA incompletely and non-cooperatively. J. Gen. Virol. 2001, 82, 2579–2588. [Google Scholar] [CrossRef]
- Tenllado, F.; Bol, J.F. Genetic dissection of the multiple functions of alfalfa mosaic virus coat protein in viral RNA replication, encapsidation, and movement. Virology 2000, 268, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Kasteel, D.T.; Van der Wel, N.N.; Jansen, K.A.; Goldbach, R.W.; Van Lent, J.W. Tubule-forming capacity of the movement proteins of alfalfa mosaic virus and brome mosaic virus. J. Gen. Virol. 1997, 78, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Kaido, M.; Okuno, T.; Mise, K. The C terminus of the movement protein of Brome mosaic virus controls the requirement for coat protein in cell-to-cell movement and plays a role in long-distance movement. J. Gen. Virol. 2004, 85, 1751–1761. [Google Scholar] [CrossRef] [PubMed]
- Kaido, M.; Inoue, Y.; Takeda, Y.; Sugiyama, K.; Takeda, A.; Mori, M.; Tamai, A.; Meshi, T.; Okuno, T.; Mise, K. Downregulation of the NbNACa1 gene encoding a movement-protein-interacting protein reduces cell-to-cell movement of Brome mosaic virus in Nicotiana benthamiana. Mol. Plant-Microbe Interact. 2007, 20, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Canto, T.; Palukaitis, P. Are tubules generated by the 3a protein necessary for cucumber mosaic virus movement? Mol. Plant-Microbe Interact. 1999, 12, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Palukaitis, P.; García-Arenal, F. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar] [PubMed]
- Nagano, H.; Mise, K.; Furusawa, I.; Okuno, T. Conversion in the requirement of coat protein in cell-to-cell movement mediated by the cucumber mosaic virus movement protein. J. Virol. 2001, 75, 8045–8053. [Google Scholar] [CrossRef] [Green Version]
- Andreev, I.A.; Kim, S.H.; Kalinina, N.O.; Rakitina, D.V.; Fitzgerald, A.G.; Palukaitis, P.; Taliansky, M.E. Molecular interactions between a plant virus movement protein and RNA: Force spectroscopy investigation. J. Mol. Biol. 2004, 339, 1041–1047. [Google Scholar] [CrossRef]
- Herranz, M.C.; Pallas, V. RNA-binding properties and mapping of the RNA-binding domain from the movement protein of Prunus necrotic ringspot virus. J. Gen. Virol. 2004, 85, 761–768. [Google Scholar] [CrossRef]
- Waigmann, E.; Lucas, W.J.; Citovsky, V.; Zambryski, P. Direct functional assay for tobacco mosaic virus cell-to-cell movement protein and identification of a domain involved in increasing plasmodesmal permeability. Proc. Natl. Acad. Sci. USA 1994, 91, 1433–1437. [Google Scholar] [CrossRef] [Green Version]
- Epel, B.L. Plant viruses spread by diffusion on ER-associated movement-protein-rafts through plasmodesmata gated by viral induced host beta-1,3-glucanases. Semin Cell Dev. Biol. 2009, 20. [Google Scholar] [CrossRef]
- Zavaliev, R.; Sagi, G.; Gera, A.; Epel, B.L. The constitutive expression of Arabidopsis plasmodesmal-associated class 1 reversibly glycosylated polypeptide impairs plant development and virus spread. J. Exp. Bot. 2010, 61, 131–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkar-Purushothama, C.R.; Brosseau, C.; Giguère, T.; Sano, T.; Moffett, P.; Perreault, J.-P. Small RNA derived from the virulence modulating region of the potato spindle tuber viroid silences callose synthase genes of tomato plants. Plant Cell 2015, 27, 2178–2194. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Lee, J.-Y. Arabidopsis callose synthases CalS1/8 regulate plasmodesmal permeability during stress. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef]
- Yan, D.; Yadav, S.R.; Paterlini, A.; Nicolas, W.J.; Petit, J.D.; Brocard, L.; Belevich, I.; Grison, M.S.; Vaten, A.; Karami, L. Sphingolipid biosynthesis modulates plasmodesmal ultrastructure and phloem unloading. Nat. Plants 2019, 5, 604–615. [Google Scholar] [CrossRef]
- Adams, M.J.; Adkins, S.; Bragard, C.; Gilmer, D.; Li, D.; MacFarlane, S.A.; Wong, S.-M.; Melcher, U.; Ratti, C.; Ryu, K.H. ICTV virus taxonomy profile: Virgaviridae. J. Gen. Virol. 2017, 98, 1999. [Google Scholar] [CrossRef]
- Rojas, M.R.; Maliano, M.R.; de Souza, J.O.; Vasquez-Mayorga, M.; de Macedo, M.A.; Ham, B.-K.; Gilbertson, R.L. Cell-to-cell movement of plant viruses: A diversity of mechanisms and strategies. In Current Research Topics in Plant Virology; Springer: Cham, Switzerland, 2016; pp. 113–152. [Google Scholar]
- Sheshukova, E.V.; Ershova, N.M.; Kamarova, K.A.; Dorokhov, Y.L.; Komarova, T.V. The Tobamoviral Movement Protein: A “Conditioner” to Create a Favorable Environment for Intercellular Spread of Infection. Front. Plant Sci. 2020, 11, 959. [Google Scholar] [CrossRef]
- Curin, M.; Ojangu, E.-L.; Trutnyeva, K.; Ilau, B.; Truve, E.; Waigmann, E. MPB2C, a microtubule-associated plant factor, is required for microtubular accumulation of tobacco mosaic virus movement protein in plants. Plant Physiol. 2007, 143, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Kleinow, T.; Tanwir, F.; Kocher, C.; Krenz, B.; Wege, C.; Jeske, H. Expression dynamics and ultrastructural localization of epitope-tagged Abutilon mosaic virus nuclear shuttle and movement proteins in Nicotiana benthamiana cells. Virology 2009, 391, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.; Tzfira, T. Bean dwarf mosaic virus: A model system for the study of viral movement. Mol. Plant Pathol. 2010, 11, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.V. Plant antiviral immunity against geminiviruses and viral counter-defense for survival. Front. Microbiol. 2019, 10, 1460. [Google Scholar] [CrossRef] [Green Version]
- Lazarowitz, S.G.; Beachy, R.N. Viral movement proteins as probes for intracellular and intercellular trafficking in plants. Plant Cell 1999, 11, 535–548. [Google Scholar] [CrossRef] [Green Version]
- van Lent, J.; Storms, M.; van der Meer, F.; Wellink, J.; Goldbach, R. Tubular structures involved in movement of cowpea mosaic virus are also formed in infected cowpea protoplasts. J. Gen. Virol. 1991, 72, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Carluccio, A.V.; Zicca, S.; Stavolone, L. Hitching a ride on vesicles: Cauliflower mosaic virus movement protein trafficking in the endomembrane system. Plant Physiol. 2014, 164, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Taliansky, M.E.; Robinson, D.J. Molecular biology of umbraviruses: Phantom warriors. J. Gen. Virol. 2003, 84, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Ritzenthaler, C.; Hofmann, C. Tubule-Guided Movement of Plant Viruses. In Viral Transport in Plants; Waigmann, E., Heinlein, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 7. [Google Scholar] [CrossRef]
- Carvalho, C.M.; Wellink, J.; Ribeiro, S.G.; Goldbach, R.W.; Van Lent, J.W.M. The C-terminal region of the movement protein of Cowpea mosaic virus is involved in binding to the large but not to the small coat protein. J. Gen. Virol. 2003, 84, 2271–2277. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Meng, C.; Xie, L.; Hong, J.; Zhou, X. Cell-to-cell trafficking, subcellular distribution, and binding to coat protein of Broad bean wilt virus 2 VP37 protein. Virus Res. 2009, 143, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wang, G.; Zhang, L. Alfalfa mosaic virus movement protein induces tubules in plant protoplasts. Mol. Plant-Microbe Interact. 1997, 10, 1010–1014. [Google Scholar] [CrossRef]
- Callaway, A.; Giesman-Cookmeyer, D.; Gillock, E.T.; Sit, T.L.; Lommel, S.A. The multifunctional capsid proteins of plant RNA viruses. Annu. Rev. Phytopathol. 2001, 39, 419–460. [Google Scholar] [CrossRef]
- Rao, A.L.N.; Cooper, B. Capsid protein gene and the type of host plant differentially modulate cell-to-cell movement of cowpea chlorotic mottle virus. Virus Genes 2006, 32, 219–227. [Google Scholar] [CrossRef]
- Sasaki, N.; Arimoto, M.; Nagano, H.; Mori, M.; Kaido, M.; Mise, K.; Okuno, T. The movement protein gene is involved in the virus-specific requirement of the coat protein in cell-to-cell movement of bromoviruses. Arch. Virol. 2003, 148, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Zhang, F.; Zhang, Y.; Liang, Y. RNA interference: A natural immune system of plants to counteract biotic stressors. Cells 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Chen, J.; Janda, M.; Sullivan, M.; den Boon, J.; Ahlquist, P. A positive-strand RNA virus replication complex parallels form and function of retrovirus capsids. Mol. Cell 2002, 9, 505–514. [Google Scholar] [CrossRef]
- Díaz-Pendón, J.A.; Ding, S.W. Direct and indirect roles of viral suppressors of RNA silencing in pathogenesis. Annu. Rev. Phytopathol 2008, 46, 303–326. [Google Scholar] [CrossRef]
- Chiu, M.; Chen, I.; Baulcombe, D.C.; Tsai, C. The silencing suppressor P25 of Potato virus X interacts with Argonaute1 and mediates its degradation through the proteasome pathway. Mol. Plant Pathol. 2010, 11, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Du, Z.; Wang, C.; Cai, L.; Hu, M.; Lin, Q.; Wu, Z.; Li, Y.; Xie, L. Identification of Pns6, a putative movement protein of RRSV, as a silencing suppressor. Virol. J. 2010, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaegashi, H.; Takahashi, T.; Isogai, M.; Kobori, T.; Ohki, S.; Yoshikawa, N. Apple chlorotic leaf spot virus 50 kDa movement protein acts as a suppressor of systemic silencing without interfering with local silencing in Nicotiana benthamiana. J. Gen. Virol. 2007, 88, 316–324. [Google Scholar] [CrossRef]
- Renovell, A.; Vives, M.C.; Ruiz-Ruiz, S.; Navarro, L.; Moreno, P.; Guerri, J. The Citrus leaf blotch virus movement protein acts as silencing suppressor. Virus Genes 2012, 44, 131–140. [Google Scholar] [CrossRef]
- Zvereva, A.S.; Golyaev, V.; Turco, S.; Gubaeva, E.G.; Rajeswaran, R.; Schepetilnikov, M.V.; Srour, O.; Ryabova, L.A.; Boller, T.; Pooggin, M.M. Viral protein suppresses oxidative burst and salicylic acid-dependent autophagy and facilitates bacterial growth on virus-infected plants. New Phytol. 2016, 211, 1020–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mérai, Z.; Kerényi, Z.; Kertész, S.; Magna, M.; Lakatos, L.; Silhavy, D. Double-stranded RNA binding may be a general plant RNA viral strategy to suppress RNA silencing. J. Virol. 2006, 80, 5747–5756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, S.; Wang, B.; Guo, X.; Yu, J.; Wang, X.; Xu, K.; Zhai, Y.; Wang, J.; Liu, Z. Identification of two RNA silencing suppressors from banana bunchy top virus. Arch. Virol. 2009, 154, 1775. [Google Scholar] [CrossRef] [PubMed]
- Kasschau, K.D.; Carrington, J.C. Long-distance movement and replication maintenance functions correlate with silencing suppression activity of potyviral HC-Pro. Virology 2001, 285, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.S.; Liu, J.; Cheng, N.-H.; Folimonov, A.; Hou, Y.-M.; Bao, Y.; Katagi, C.; Carter, S.A.; Nelson, R.S. The Tobacco mosaic virus 126-kDa protein associated with virus replication and movement suppresses RNA silencing. Mol. Plant-Microbe Interact. 2004, 17, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Bayne, E.H.; Rakitina, D.V.; Morozov, S.Y.; Baulcombe, D.C. Cell-to-cell movement of potato potexvirus X is dependent on suppression of RNA silencing. Plant J. 2005, 44, 471–482. [Google Scholar] [CrossRef]
- Liu, J.Z.; Blancaflor, E.B.; Nelson, R.S. The tobacco mosaic virus 126-kilodalton protein, a constituent of the virus replication complex, alone or within the complex aligns with and traffics along microfilaments. Plant Physiol. 2005, 138, 1853–1865. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family/Genus | Species | MP | MP—Properties | MP—Mode of Virus Intercellular Movement | Assisting Viral Proteins and Their Putative Function | Interacting Host Proteins and Their Putative Function | References |
|---|---|---|---|---|---|---|---|
| Virgaviridae/Tobamovirus | TMV | 30 kDa (P30) |
|
|
|
| [1,11,15,22,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59] |
| Tombusviridae/Dianthovirus | RCNMV | 35 kDa (P35) |
| PD-gating (increases SEL) |
|
| [60,61,62,63,64] |
| Tombusviridae/Carmovirus MPs-(often termed double gene block proteins) | TCV | P8 and P9 |
| PD-gating (increase SEL) |
|
| [65,66] |
| CarMV | P7 and P9 |
| PD-gating (increase SEL) | NK | NK | [67] | |
| PFBV | P7 and P12 |
| PD-gating (increase SEL) | NK | NK | [68] | |
| MNSV | P7A and P7B |
|
| CP—R2-subdomain, also a VSR | Movement is energy-dependent on unknown host protein (s) | [69,70] | |
| Kitaviridae/Higrivirus | HGSV | BMB1 and BMB2 |
|
| NK | NK | [71,72] |
| Tombusviridae/Tombusvirus | TBSV | P22 |
| PD-gating (increases SEL) | P19—assist systemic movement through silencing suppression | HFI22—leucine zipper homeodomain protein interacts with P22 for delivery of P22/RNA complexes through PD for intercellular movement | [73] |
| Closteroviridae/Closterovirus | BYV CTV | P6, P64, CP, CPm and HSP70h |
| PD-gating (increase SEL) | P20—interacts with HSP70h for long-distance systemic movement | Class VII myosins—motility and targeting of HSP70h to PD | [74] |
| Closteroviridae/Crinivirus | LIYV | P6, P64, CP, CPm and HSP70h P26 (essential for virus systemic infection) |
| PD-gating (increases SEL) | P9—unknown function | NK | [75] |
| Potyviridae/Potyvirus | TEV, TuMV, SMV, BCNMV, LMV | P3N-PIPO, CP (TEV/TuMV/SMV) CP and HC-Pro (BCNMV/LMV) |
| PD-gating (increase SEL) |
| pCAP1—binds to P3N-PIPO aids its localization to the PD and intercellular movement | [76,77,78,79,80] |
| Luteoviridae/Polerovirus | PLRV, TuYV, PeVYV | 17 kDa (P17) 175 kDa) P4 (PeVYV) |
| PD-gating (increase SEL) | P3a—localization to the outer membrane of mitochondria and plastid |
| [81,82,83] |
| Luteoviridae/Luteovirus | BYDV | 17 kDa (P4) |
| PD-gating (increases SEL) | P3a—localization to the outer membrane of mitochondria and plastid | NK | [84] |
| Geminiviridae/Begomovirus | BDMV AbMV, ToLCNDV | BC1 (BL1) |
| PD-gating (increases SEL) |
|
| [85,86] |
| Rhabdoviridae/Nucleorhadbdovirus | RYSV, SYNV, MMV, MFSV, PYDV, EMDV, CaLCuV | P3 (RYSV and MMV) P3 or sc4 (SYNV) P4 (MFSV) Y (PYDV and EMDV) |
| PD-gating (increase SEL) |
|
| [87,88] |
| Rhabdoviridae/Cytorhabdovirus | LNYV, ADV, TYMaV | 4b (LNVY) P3 (ADV and TYMaV) |
| PD-gating (increase SEL) | P (phospho) protein— formation of movement complex (ADV) | P3 (LNVY) —interacts with MT-associated VOZ1-like transcriptional activator—for anchoring the virus movement complexes to the MT network for intracellular trafficking and aiding in the intercellular movement | [87,88] |
| Alphaflexiviridae/Potexvirus and Betaflexiviridae/Foveavirusa | PVX GRSPaV | TGB1 TGB2 TGB3 TGB1 TGB2 TGB3 Some members have TMV 30 K-like MP |
|
|
| TGB1-
| [12,18,37,89,90,91,92,93,94] |
| Virgaviridae/Hordeivirus | BSMV | TGB1 TGB2 TGB3 |
|
|
|
| [18,89,90,91,92,93] |
| Bunyaviridae/Benyviruses | BNYVV | TGB1 TGB2 TGB3 |
|
| NK | NK | [12] |
| Tombusviridae/ Umbravirus | PEMV-2 | P26 and P27 |
| Tubule formation | NK | NK | [95] |
| Caulimoviridae /Caulimovirus | CaMV | 38 kDa MP (P1) | Binds RNA | Tubule formation |
|
| [96,97,98,99] |
| Secoviridae/Nepovirus | GFLV | 38 kDa MP | Binds RNA | Tubule formation | CP |
| [100,101,102] |
| Comoviridae/ Comovirus | CPMV | 48 kDa MP | Has large CP, GTP, ssRNA, ssDNA-binding regions | Tubule formation | Large CP (CPL)-37 kDa | PDLPs-recruitment of MP to PD | [103,104,105,106,107] |
| Bunyaviridae/ Tospovirus | TSWV | NSm protein |
| Tubule formation | TSWV N—protein-recognition of nucleocapsid structures |
| [96,97,98,99,108,109,110] |
| Tombusviridae/ Umbravirus | GRV | 28 kDa, MP (P4) |
| Tubule formation | ORF3 protein assist for vRNP formation for cell-to-cell movement | Fibrillarin—vRNP complex formation for transport, interacts with virus ORF3 protein | [111,112] |
| Bromoviridae/Alfamovirus | AMV | 32 kDa MP (P3) |
|
| CP |
| [113] |
| Bromoviridae/ Bromovirus | BMV | 32 kDa MP (3a) |
|
| CP (BMV-MI strain) | NbNACa1—PD localization | [114,115,116] |
| Bromoviridae/ Cucumovirus | CMV | 32 to 36 kDa MP (3a) 2b-for long-distance movement |
|
| CP (for tubule formation?) |
| [117,118,119,120] |
| Bromoviridae/Ilarvirus | PNRSV | 32 to 36 kDa MP (3a) |
|
| CP | NK | [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, G.; Dasgupta, I. Variability, Functions and Interactions of Plant Virus Movement Proteins: What Do We Know So Far? Microorganisms 2021, 9, 695. https://doi.org/10.3390/microorganisms9040695
Kumar G, Dasgupta I. Variability, Functions and Interactions of Plant Virus Movement Proteins: What Do We Know So Far? Microorganisms. 2021; 9(4):695. https://doi.org/10.3390/microorganisms9040695
Chicago/Turabian StyleKumar, Gaurav, and Indranil Dasgupta. 2021. "Variability, Functions and Interactions of Plant Virus Movement Proteins: What Do We Know So Far?" Microorganisms 9, no. 4: 695. https://doi.org/10.3390/microorganisms9040695