
Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Bacterial Treatment and Nematode Inoculum
2.2. Pine Seedling Assay
2.3. Pine Rhizosphere and Soil Metagenomic DNA Extraction, Sequencing, and Bioinformatics Analysis
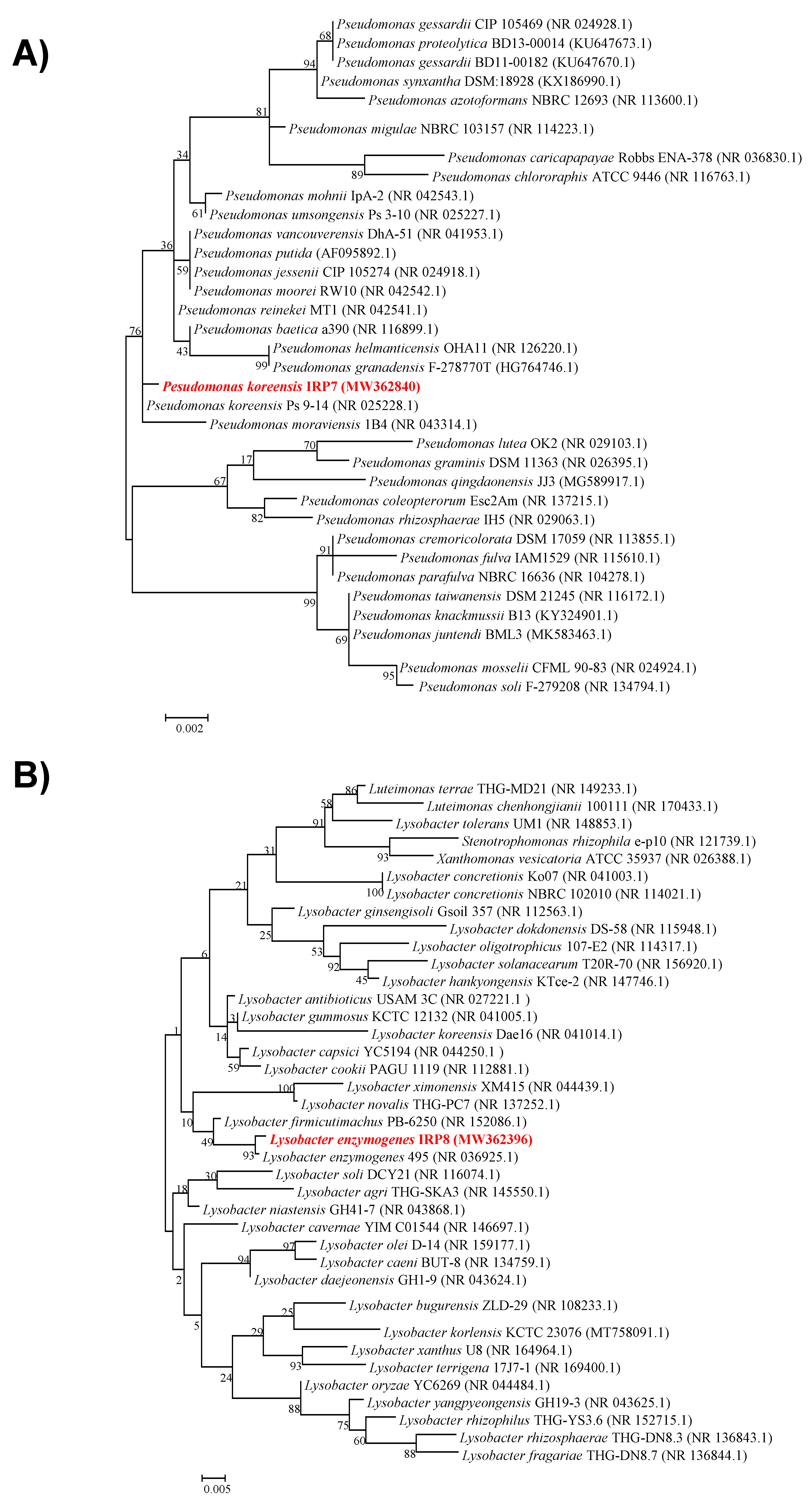
2.4. Taxonomic Identification of Resistance-Inducing Bacterial Strains IRP7 and IRP8
2.5. Statistical Analysis
3. Results
3.1. Foliar Treatment with Selected Bacterial Strains Reduced Pine Wilt Disease (PWD) Severity in Inoculated Seedlings
3.2. Sequencing Results and Sequence Analysis
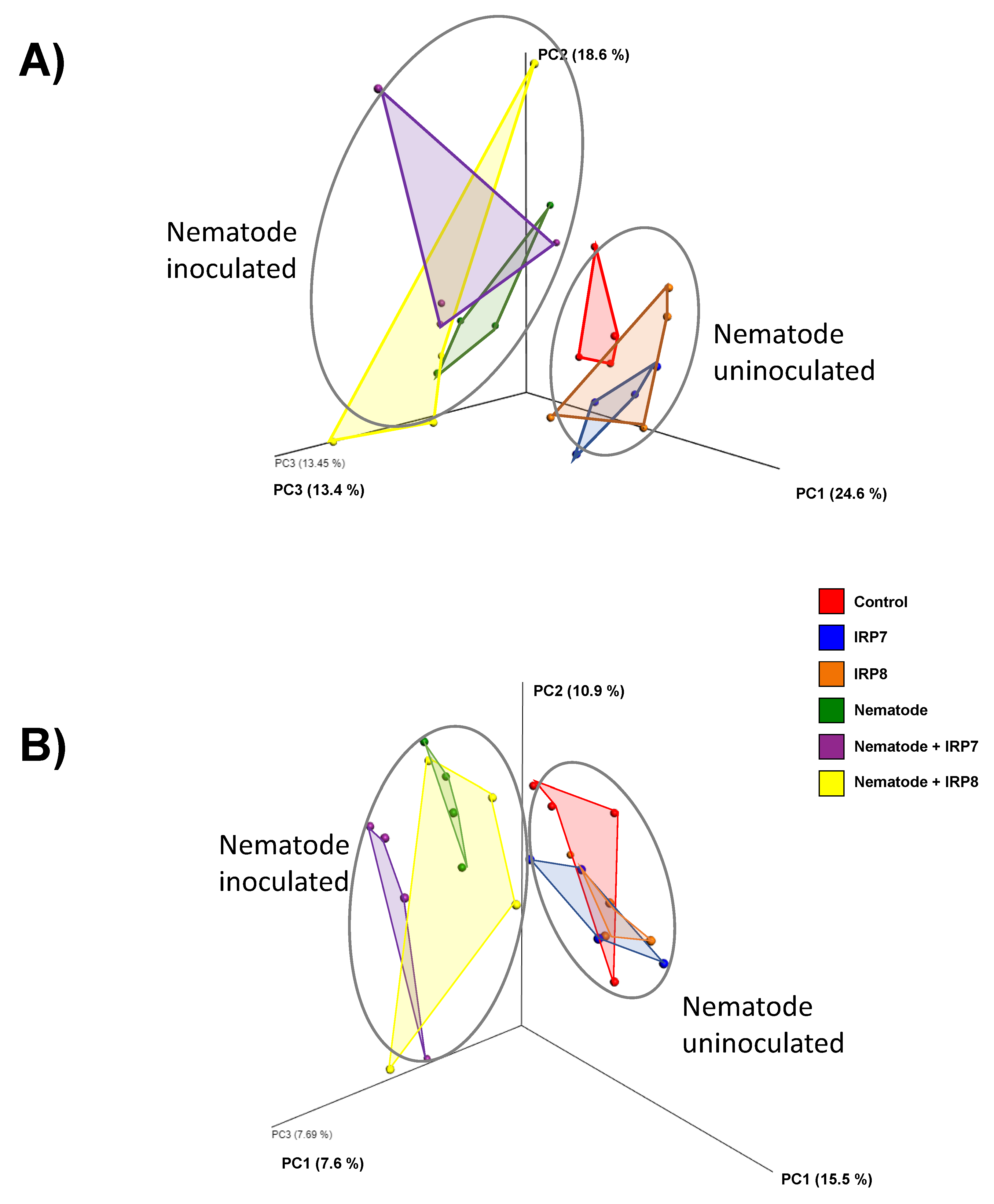
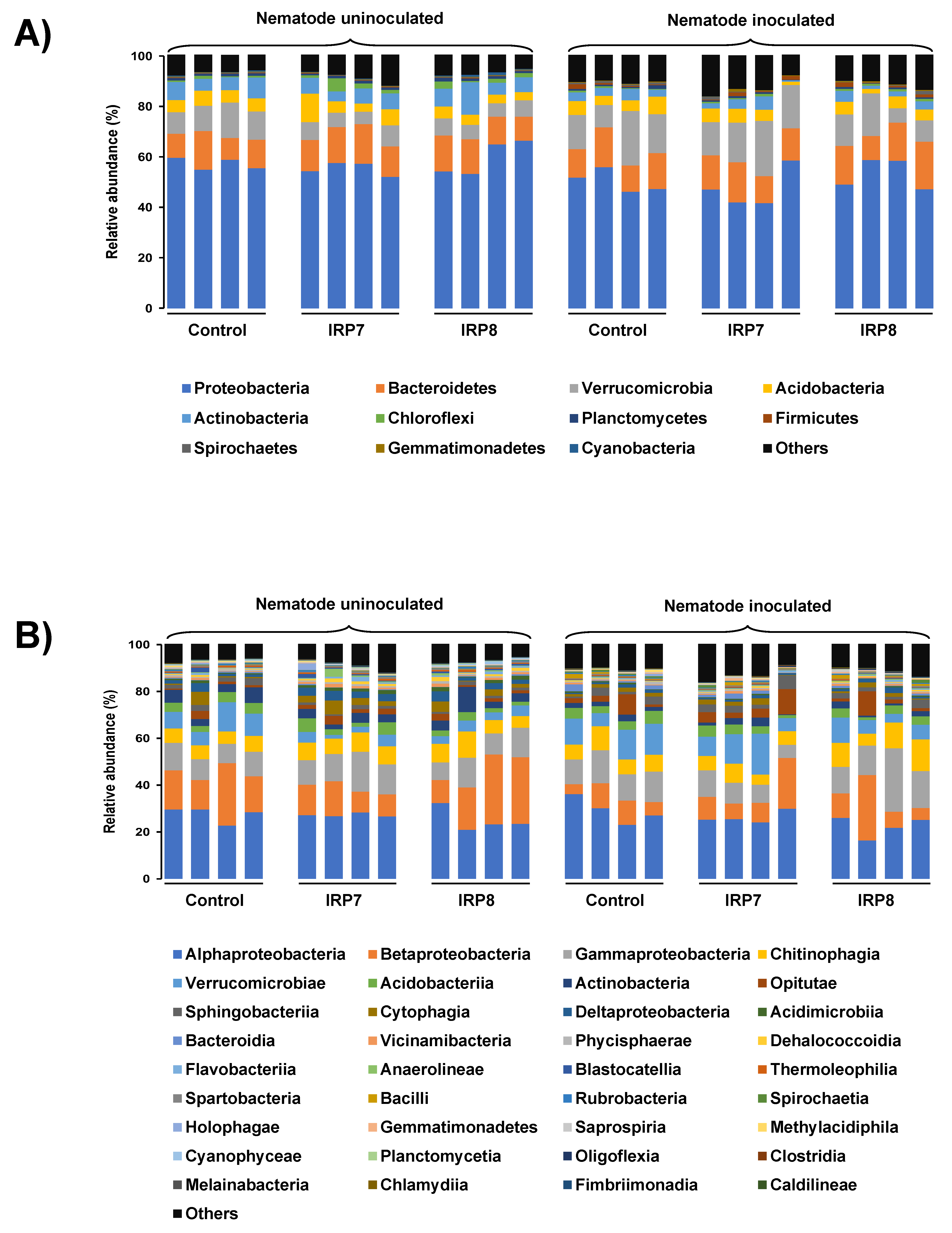
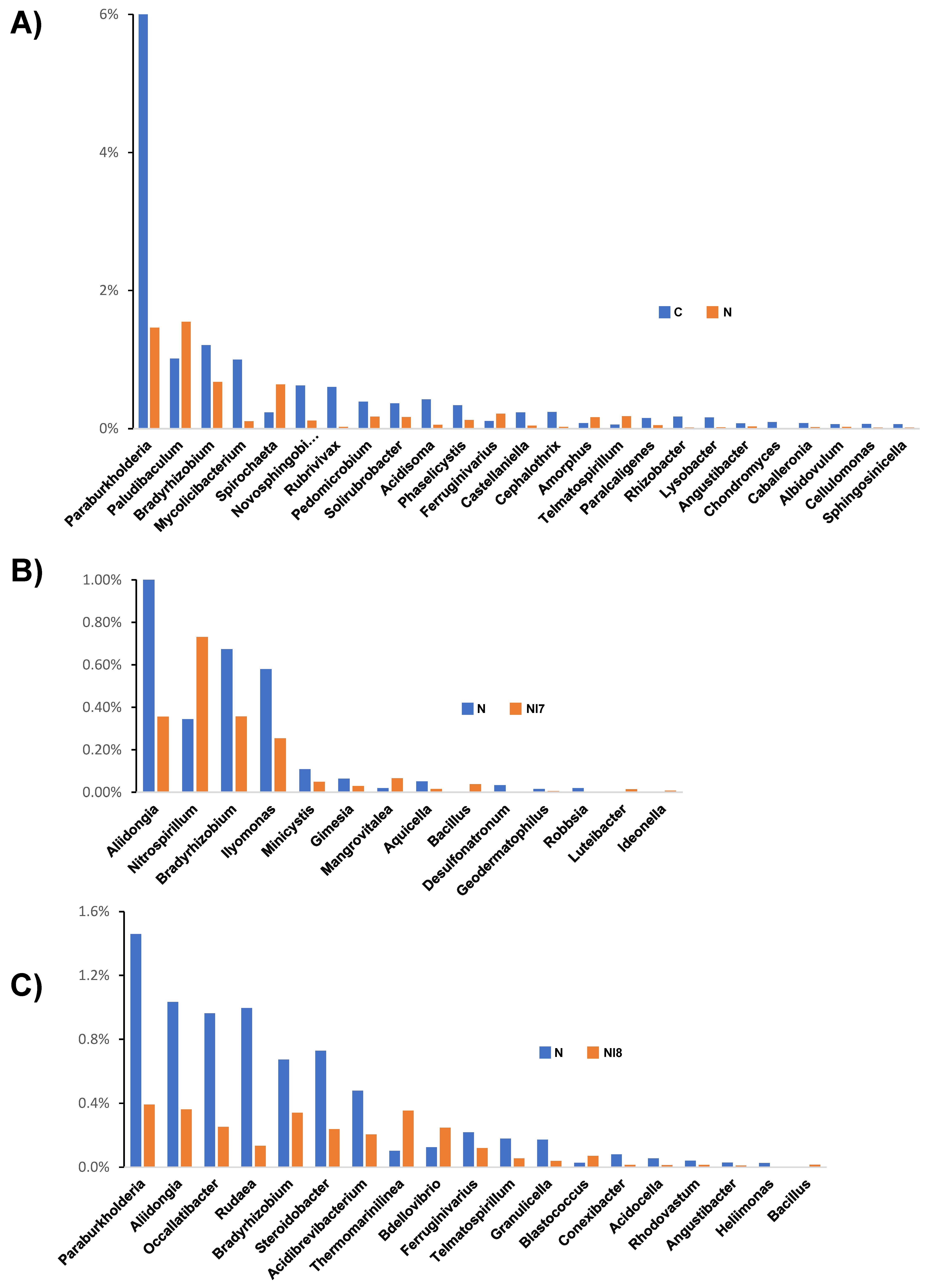
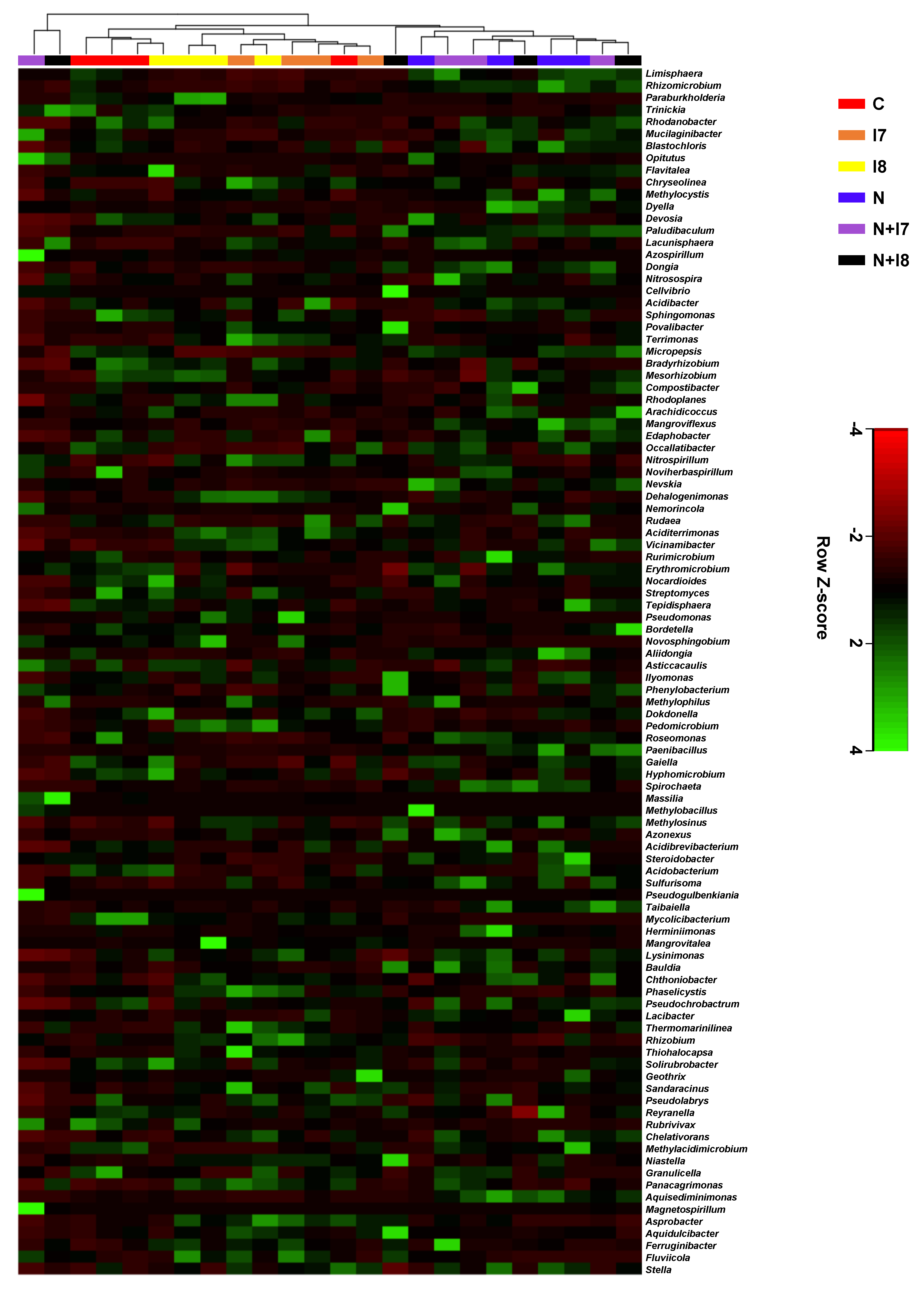
3.3. Pine Rhizosphere Bacterial Composition Structure Affected by Nematode Inoculation and Bacterial Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tóth, Á. Bursaphelenchus xylophilus, the pinewood nematode: Its significance and a historical review. Acta Biol. Szeged. 2011, 55, 213–217. [Google Scholar]
- Kiyohara, T.; Tokushige, Y. Inoculation experiments of a nematode, Bursaphelenchus sp., onto pine trees. J. Jpn. For. Soc. 1971, 53, 210–218. [Google Scholar]
- Burgermeister, W.; Braasch, H.; Sousa, E.; Penas, A.C.; Mota, M.; Metge, K.; Bravo, M.A. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, causal agent of pine wilt disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef] [PubMed]
- Mannaa, M.; Seo, Y.S. Plants under the Attack of Allies: Moving towards the Plant Pathobiome Paradigm. Plants 2021, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Le Dang, Q.; Choi, G.J.; Kim, J.C.; Cheon, H.M.; Jang, K.S.; Son, S.W.; Choi, Y.H.; Lim, C.H. Pyochelin isolated from Burkholderia arboris KRICT1 carried by pine wood nematodes exhibits phytotoxicity in pine callus. Nematology 2011, 13, 521–528. [Google Scholar] [CrossRef]
- Guo, Q.; Guo, D.; Zhao, B.; Xu, J.; Li, R. Two Cyclic Dipeptides from Pseudomonas fluorescens GcM5-1A Carried by the Pine Wood Nematode and Their Toxicities to Japanese Black Pine Suspension Cells and Seedlings in vitro. J. Nematol. 2007, 39, 243–247. [Google Scholar] [PubMed]
- Kawazu, K.; Zhang, H.; Yamashita, H.; Kanzaki, H. Relationship between the pathogenicity of the pine wood nematode, Bursaphelenchus xylophilus, and phenylacetic acid production. Biosci. Biotechnol. Biochem. 1996, 60, 1413–1415. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.G.; Li, R.G. The Role of Bacteria Associated with the Pine Wood Nematode in Pathogenicity and Toxin-Production Related to Pine Wilt. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 250–259. [Google Scholar]
- Han, Z.M.; Hong, Y.D.; Zhao, B.G. A study on pathogenicity of bacteria carried by pine wood nematodes. J. Phytopathol. 2003, 151, 683–689. [Google Scholar] [CrossRef]
- Proença, D.N.; Francisco, R.; Santos, C.V.; Lopes, A.; Fonseca, L.; Abrantes, I.M.; Morais, P.V. Diversity of bacteria associated with Bursaphelenchus xylophilus and other nematodes isolated from Pinus pinaster trees with pine wilt disease. PLoS ONE 2010, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Stud. China 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Kwon, T.S. Change of abundance of arthropods in pine forests caused by aerial insecticide spray. Arch. Environ. Contam. Toxicol. 2008, 54, 92–106. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, D.S.; Lee, S.G.; Park, N.C.; Lee, D.W. Selection of trunk injection pesticides for preventive of pine wilt disease, Bursaphelenchus xylophilus on Japanese black pine (Pinus thunbergii). Korean J. Pestic. Sci. 2009, 13, 267–274. [Google Scholar]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine wilt disease: A threat to European forestry. Eur. J. Plant Pathol. 2012, 133, 89–99. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wang, Z.; Fang, Z.M.; Zhang, D.L.; Gu, L.J.; Liu, L.; Sung, C.K. Attraction of pinewood nematode to endoparasitic nematophagous fungus Esteya vermicola. Curr. Microbiol. 2010, 60, 387–392. [Google Scholar]
- Wang, C.Y.; Yin, C.; Fang, Z.M.; Wang, Z.; Wang, Y.B.; Xue, J.J.; Gu, L.J.; Sung, C.K. Using the nematophagous fungus Esteya vermicola to control the disastrous pine wilt disease. Biocontrol. Sci. Technol. 2018, 28, 268–277. [Google Scholar]
- Chu, W.H.; Dou, Q.; Chu, H.L.; Wang, H.H.; Sung, C.K.; Wang, C.Y. Research advance on Esteya vermicola, a high potential biocontrol agent of pine wilt disease. Mycol. Prog. 2015, 14, 1–9. [Google Scholar]
- Nascimento, F.X.; Vicente, C.S.; Barbosa, P.; Espada, M.; Glick, B.R.; Mota, M.; Oliveira, S. Evidence for the involvement of ACC deaminase from Pseudomonas putida UW4 in the biocontrol of pine wilt disease caused by Bursaphelenchus xylophilus. BioControl 2013, 58, 427–433. [Google Scholar] [CrossRef]
- Xu, F.; Xu, K.; Xie, C.; Zhang, P.; Shin, S.; Cheong, Y. Studies on Scleroderma guani to control the pine sawyer beetle, Monochamus alternatus. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 379–388. [Google Scholar]
- Kosaka, H.; Aikawa, T.; Ogura, N.; Tabata, K.; Kiyohara, T. Pine wilt disease caused by the pine wood nematode: The induced resistance of pine trees by the avirulent isolates of nematode. Eur. J. Plant Pathol. 2001, 107, 667–675. [Google Scholar] [CrossRef]
- Kim, N.; Jeon, H.W.; Mannaa, M.; Jeong, S.I.; Kim, J.; Kim, J.; Lee, C.; Park, A.R.; Kim, J.C.; Seo, Y.S. Induction of resistance against pine wilt disease caused by Bursaphelenchus xylophilus using selected pine endophytic bacteria. Plant Pathol. 2019, 68, 434–444. [Google Scholar] [CrossRef]
- Mannaa, M.; Han, G.; Jeon, H.W.; Kim, J.; Kim, N.; Park, A.R.; Kim, J.C.; Seo, Y.S. Influence of resistance-inducing chemical elicitors against pine wilt disease on the rhizosphere microbiome. Microorganisms 2020, 8, 884. [Google Scholar] [CrossRef]
- Viglierchio, D.R.; Schmitt, R.V. On the methodology of nematode extraction from field samples: Baermann funnel modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar] [PubMed]
- Kwon, H.R.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Sung, N.D.; Kang, M.S.; Moon, Y.; Lee, S.K.; Kim, J.C. Suppression of pine wilt disease by an antibacterial agent, oxolinic acid. Pest Manag. Sci. 2010, 66, 634–639. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Li, W.; Chang, Y. CD-HIT-OTU-MiSeq, an improved approach for clustering and analyzing paired end MiSeq 16S rRNA sequences. bioRxiv 2017, 153783. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, D.A.; Ryan, P.D. Palaeontological statistics sofware package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Xiang, N.; Lawrence, K.S.; Donald, P.A. Biological control potential of plant growth-promoting rhizobacteria suppression of Meloidogyne incognita on cotton and Heterodera glycines on soybean: A review. J. Phytopathol. 2018, 166, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Adam, M.; Heuer, H.; Hallmann, J. Bacterial antagonists of fungal pathogens also control root-knot nematodes by induced systemic resistance of tomato plants. PLoS ONE 2014, 9, e90402. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: Feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Shcherbakova, E.N.; Shcherbakov, A.V.; Andronov, E.E.; Gonchar, L.N.; Kalenskaya, S.M.; Chebotar, V.K. Combined pre-seed treatment with microbial inoculants and Mo nanoparticles changes composition of root exudates and rhizosphere microbiome structure of chickpea (Cicer arietinum L.) plants. Symbiosis 2017, 73, 57–69. [Google Scholar] [CrossRef]
- Hu, L.; Robert, C.A.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; Van Der Heijden, M.G.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 1–13. [Google Scholar]
- Xie, Y.; Wright, S.; Shen, Y.; Du, L. Bioactive natural products from Lysobacter. Nat. Prod. Rep. 2012, 29, 1277–1287. [Google Scholar]
- Antoun, H.; Beauchamp, C.J.; Goussard, N.; Chabot, R.; Lalande, R. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effect on radishes (Raphanus sativus L.). In Molecular Microbial Ecology of the Soil; Hardarson, G., Broughton, W.J., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 57–67. [Google Scholar]
- Mannaa, M.; Park, I.; Seo, Y.S. Genomic features and insights into the taxonomy, virulence, and benevolence of plant-associated Burkholderia species. Int. J. Mol. Sci. 2019, 20, 121. [Google Scholar] [CrossRef] [Green Version]
- Schwab, S.; Terra, L.A.; Baldani, J.I. Genomic characterization of Nitrospirillum amazonense strain CBAmC, a nitrogen-fixing bacterium isolated from surface-sterilized sugarcane stems. Mol. Genet. Genom. 2018, 293, 997–1016. [Google Scholar] [CrossRef]
- Tiwari, S.; Prasad, V.; Lata, C. Bacillus: Plant growth promoting bacteria for sustainable agriculture and environment. In New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, J.S., Singh, D.P., Eds.; Elsevier: Amsterdam, Switzerland, 2019; pp. 43–55. [Google Scholar]
- Xiong, Y.W.; Li, X.W.; Wang, T.T.; Gong, Y.; Zhang, C.M.; Xing, K.; Qin, S. Root exudates-driven rhizosphere recruitment of the plant growth-promoting rhizobacterium Bacillus flexus KLBMP 4941 and its growth-promoting effect on the coastal halophyte Limonium sinense under salt stress. Ecotoxicol. Environ. Saf. 2020, 194, 110374. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Hernández, A.G.; Glick, B.R.; Rossi, M.J. Plant growth-promoting activities and genomic analysis of the stress-resistant Bacillus megaterium STB1, a bacterium of agricultural and biotechnological interest. Biotechnol. Rep. 2020, 25, e00406. [Google Scholar] [CrossRef]
- Probanza, A.; Garcıa, J.L.; Palomino, M.R.; Ramos, B.; Mañero, F.G. Pinus pinea L. seedling growth and bacterial rhizosphere structure after inoculation with PGPR Bacillus (B. licheniformis CECT 5106 and B. pumilus CECT 5105). Appl. Soil Ecol. 2002, 20, 75–84. [Google Scholar]
- Guglielmetti, S.; Basilico, R.; Taverniti, V.; Arioli, S.; Piagnani, C.; Bernacchi, A. Luteibacter rhizovicinus MIMR1 promotes root development in barley (Hordeum vulgare L.) under laboratory conditions. World J. Microbiol. Biotechnol. 2013, 29, 2025–2032. [Google Scholar] [CrossRef]
- Scherff, R.H. Control of bacterial blight of soybean by Bdellovibrio bacteriovorus. Can. J. Microbiol. 1973, 328, 400–402. [Google Scholar] [CrossRef]
- Saxon, E.B.; Jackson, R.W.; Bhumbra, S.; Smith, T.; Sockett, R.E. Bdellovibrio bacteriovorus HD100 guards against Pseudomonas tolaasii brown-blotch lesions on the surface of post-harvest Agaricus bisporus supermarket mushrooms. BMC Microbiol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Youdkes, D.; Helman, Y.; Burdman, S.; Matan, O.; Jurkevitch, E. Potential control of potato soft rot disease by the obligate predators Bdellovibrio and like organisms. Appl. Environ. Microbiol. 2020, 86, e02543-19. [Google Scholar] [CrossRef] [PubMed]
- Rafikova, G.F.; Korshunova, T.Y.; Minnebaev, L.F.; Chetverikov, S.P.; Loginov, O.N. A new bacterial strain, Pseudomonas koreensis IB-4, as a promising agent for plant pathogen biological control. Microbiology 2016, 85, 333–341. [Google Scholar] [CrossRef]
- Hultberg, M.; Alsberg, T.; Khalil, S.; Alsanius, B. Suppression of disease in tomato infected by Pythium ultimum with a biosurfactant produced by Pseudomonas koreensis. BioControl 2010, 55, 435–444. [Google Scholar] [CrossRef]
- Hultberg, M.; Bengtsson, T.; Liljeroth, E. Late blight on potato is suppressed by the biosurfactant-producing strain Pseudomonas koreensis 2.74, and its biosurfactant. BioControl 2010, 55, 543–550. [Google Scholar] [CrossRef]
- Li, S.; Jochum, C.C.; Yu, F.; Zaleta-Rivera, K.; Du, L.; Harris, S.D.; Yuen, G.Y. An antibiotic complex from Lysobacter enzymogenes strain C3: Antimicrobial activity and role in plant disease control. Phytopathology 2008, 98, 695–701. [Google Scholar]
- Qian, G.L.; Hu, B.S.; Jiang, Y.H.; Liu, F.Q. Identification and characterization of Lysobacter enzymogenes as a biological control agent against some fungal pathogens. Agric. Sci. China 2009, 8, 68–75. [Google Scholar]
- Chen, J.; Moore, W.H.; Yuen, G.Y.; Kobayashi, D.; Caswell-Chen, E.P. Influence of Lysobacter enzymogenes strain C3 on nematodes. J. Nematol. 2006, 38, 233–239. [Google Scholar]
- Kilic-Ekici, O.; Yuen, G.Y. Induced resistance as a mechanism of biological control by Lysobacter enzymogenes strain C3. Phytopathology 2003, 93, 1103–1110. [Google Scholar] [CrossRef] [Green Version]
- Jochum, C.C.; Osborne, L.E.; Yuen, G.Y. Fusarium head blight biological control with Lysobacter enzymogenes strain C3. Biol. Control 2006, 39, 336–344. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | OTUs | Chao1 | Shannon | Inverse Simpson |
|---|---|---|---|---|
| C | 1145 ± 98.29 | 1416.73 ± 122.95 | 7.96 ± 0.62 | 0.99 ± 0.01 |
| I7 | 1229 ± 25.91 | 1438.03 ± 38.25 | 8.42 ± 0.06 | 0.99 ± 0.00 |
| I8 | 1191 ± 159.7 | 1420.94 ± 177.89 | 7.79 ± 0.56 | 0.97 ± 0.02 * |
| N | 1125 ± 57.6 | 1350.52 ± 82.64 | 7.82 ± 0.07 | 0.99 ± 0.00 |
| N+I7 | 1112 ± 193 | 1322.57 ± 211.73 | 7.68 ± 0.73 * | 0.98 ± 0.01 |
| N+I8 | 1081 ± 98.98 | 1314.23 ± 109.29 | 7.48 ± 0.21 * | 0.97 ± 0.01 |
| Nematode-inoculated | 1106 ± 118.81 | 1329.11 ± 132.70 | 7.66 ± 0.43 | 0.98 ± 0.01 |
| Nematode-uninoculated | 1183 ± 105.13 * | 1425.23 ± 115.09 * | 8.06 ± 0.52 * | 0.98 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, G.; Mannaa, M.; Kim, N.; Jeon, H.W.; Jung, H.; Lee, H.-H.; Kim, J.; Park, J.; Park, A.R.; Kim, J.-C.; et al. Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease. Microorganisms 2021, 9, 688. https://doi.org/10.3390/microorganisms9040688
Han G, Mannaa M, Kim N, Jeon HW, Jung H, Lee H-H, Kim J, Park J, Park AR, Kim J-C, et al. Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease. Microorganisms. 2021; 9(4):688. https://doi.org/10.3390/microorganisms9040688
Chicago/Turabian StyleHan, Gil, Mohamed Mannaa, Namgyu Kim, Hee Won Jeon, Hyejung Jung, Hyun-Hee Lee, Junheon Kim, Jungwook Park, Ae Ran Park, Jin-Cheol Kim, and et al. 2021. "Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease" Microorganisms 9, no. 4: 688. https://doi.org/10.3390/microorganisms9040688