
Genetic but No Phenotypic Associations between Biocide Tolerance and Antibiotic Resistance in Escherichia coli from German Broiler Fattening Farms

 , , , and
, , , and 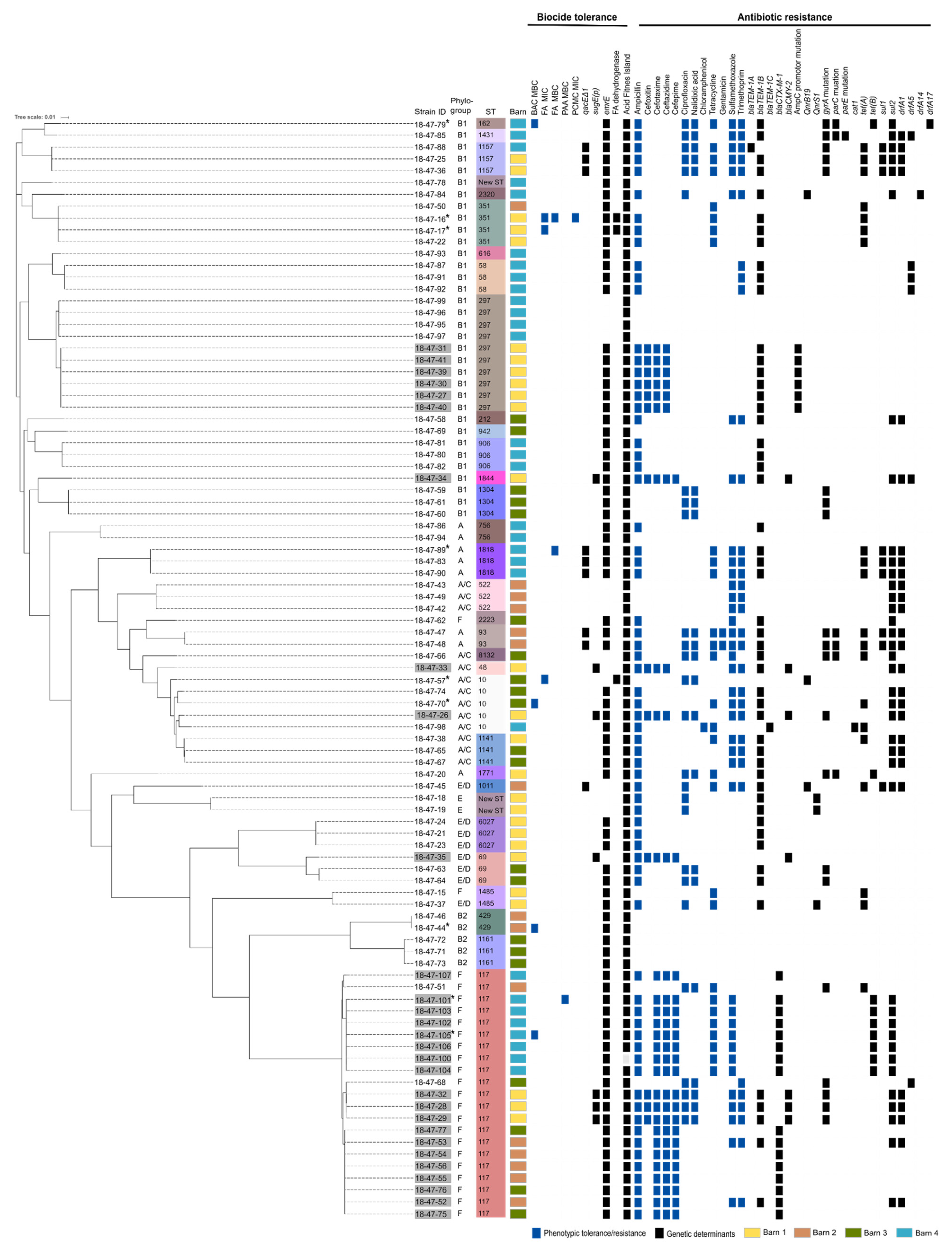{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. E. coli Isolates
2.2. Whole Genome Sequencing
2.3. Phylogenetic Analysis
2.4. Biocide Susceptibility Testing
2.4.1. Biocides
2.4.2. Minimum Inhibitory Concentration (MIC)
2.4.3. Minimum Bactericidal Concentration (MBC)
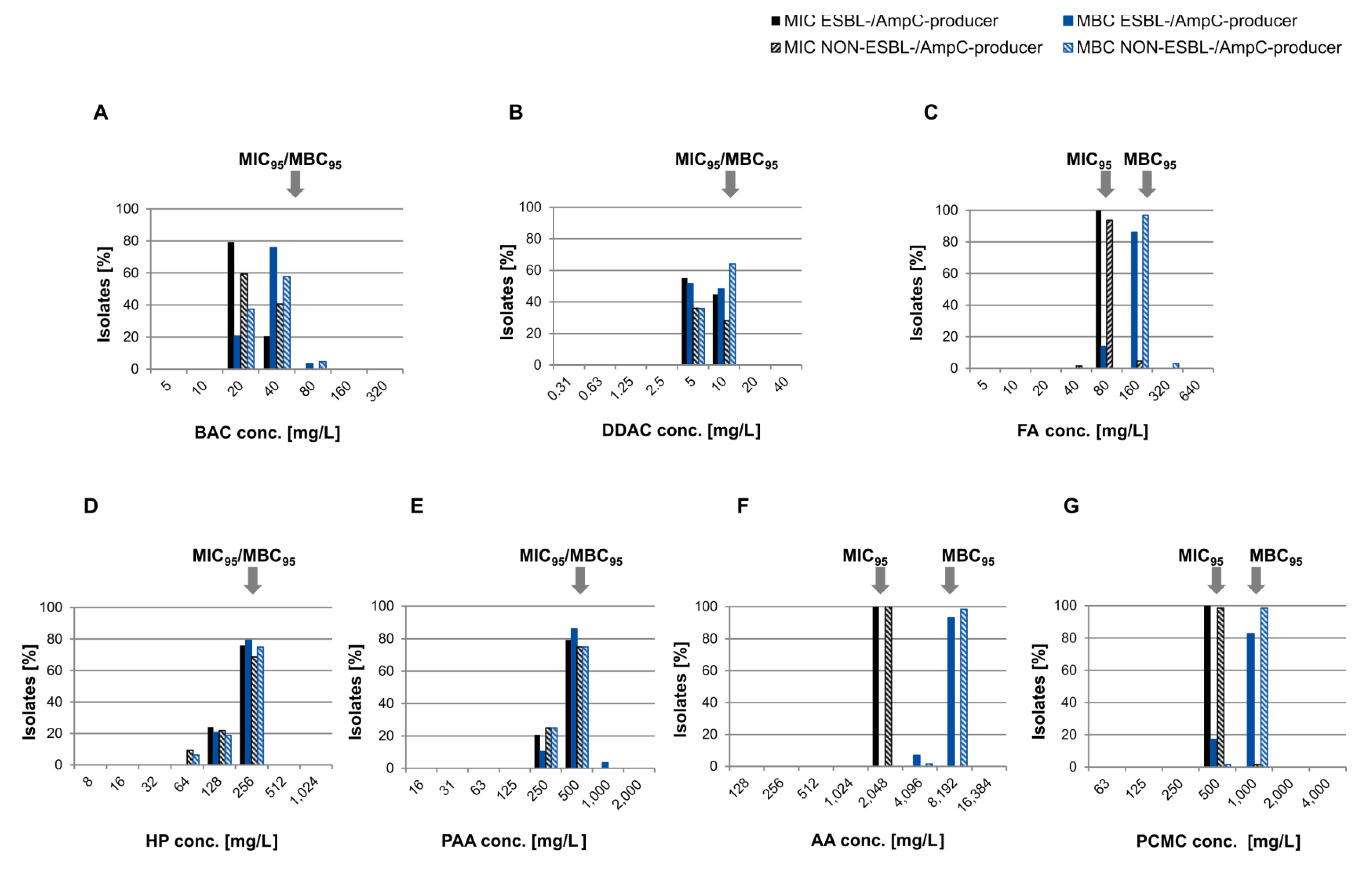
2.4.4. Determination of MIC95/MBC95
2.5. Antibiotic Susceptibility Testing
2.6. Statistical Analysis
2.7. In Silico Screening for Biocide and Metal Tolerance Determinants at Protein Level
2.8. Detection of Biocide Tolerance and Virulence Determinants at Nucleotide Level
2.9. Identification of Antibiotic Resistance Genes
2.10. Accession Numbers of Whole-Genome Sequences
3. Results
3.1. Phylogenetic Diversity and Virulence-Associated Genes
3.2. Susceptibility to Biocides
3.3. Susceptibility to Antibiotics
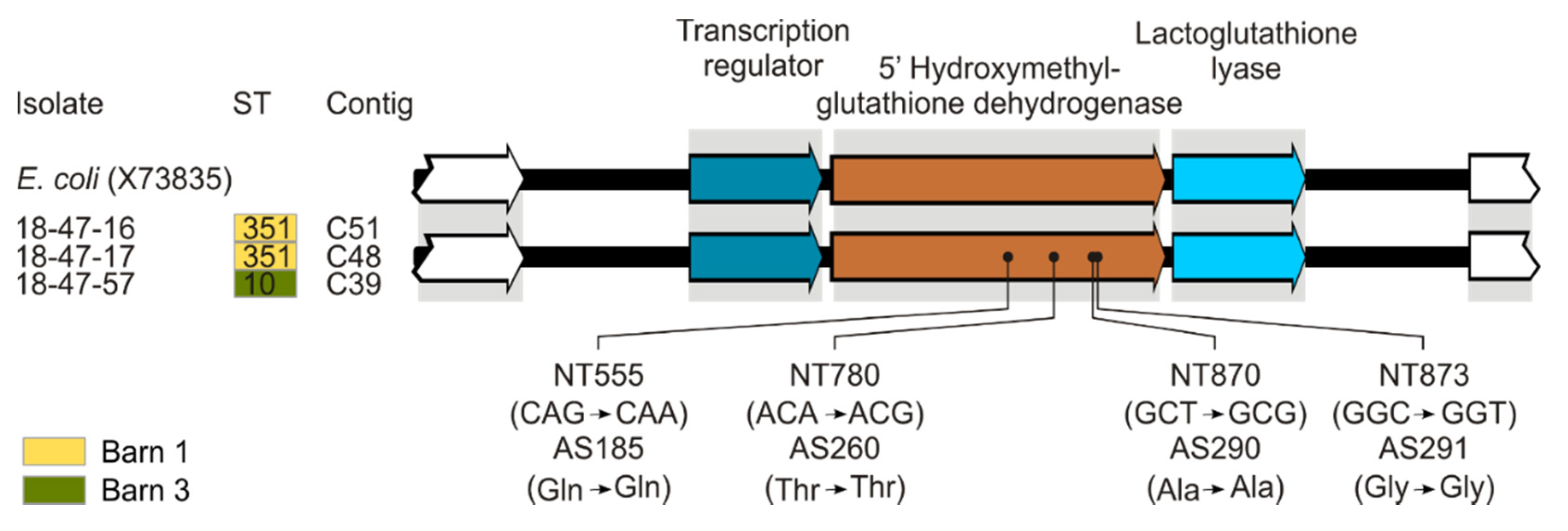
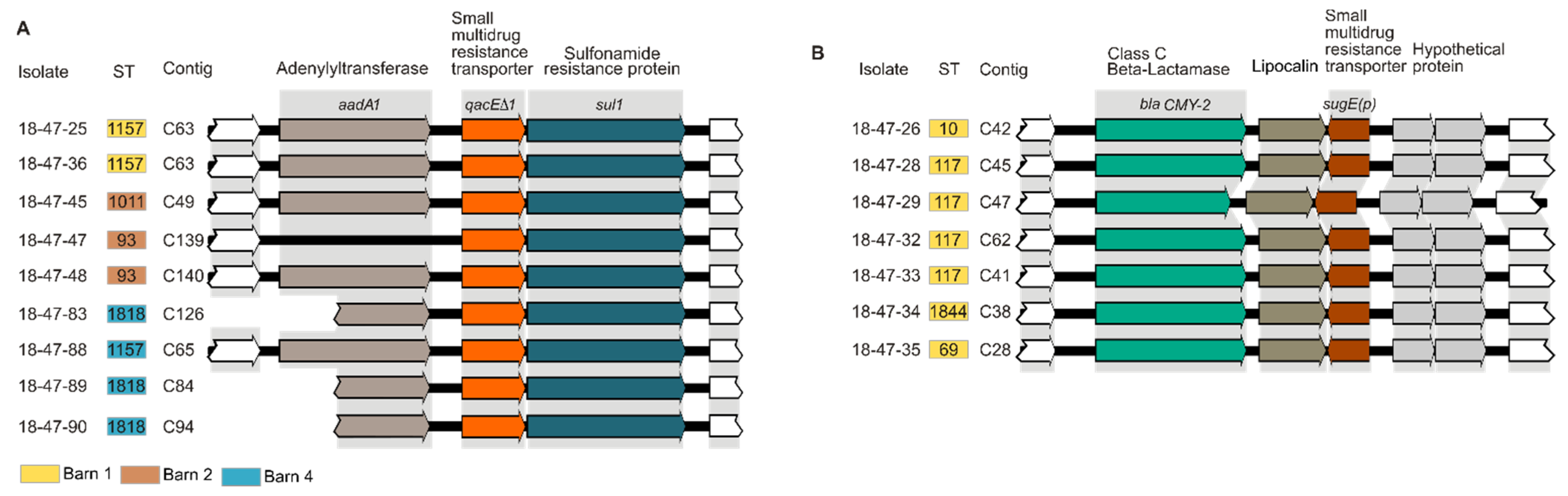
3.4. In Silico Analysis of Determinants Conferring Biocide and Metal Tolerance
3.5. In Silico Analysis of Antibiotic Resistance Gene Profiles
3.6. Association Between Reduced Biocide Susceptibility and Antibiotic Resistance and Co-occurrence of Antimicrobial Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef]
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W. Extraintestinal pathogenic Escherichia coli. Foodborne Pathog. Dis. 2007, 4, 134–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, J.; Saez-Lopez, E.; Johnson, J.R.; Romling, U.; Dobrindt, U.; Canton, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martinez-Medina, M.; et al. Escherichia coli: An old friend with new tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 2011, 9, 2322. [Google Scholar] [CrossRef] [Green Version]
- Kaesbohrer, A.; Bakran-Lebl, K.; Irrgang, A.; Fischer, J.; Kämpf, P.; Schiffmann, A.; Werckenthin, C.; Busch, M.; Kreienbrock, L.; Hille, K. Diversity in prevalence and characteristics of ESBL/pAmpC producing E. coli in food in Germany. Vet. Microbiol. 2019, 233, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Projahn, M.; Daehre, K.; Semmler, T.; Guenther, S.; Roesler, U.; Friese, A. Environmental adaptation and vertical dissemination of ESBL-/pAmpC-producing Escherichia coli in an integrated broiler production chain in the absence of an antibiotic treatment. Microb. Biotechnol. 2018, 11, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Dorado-Garcia, A.; van Duijkeren, E.; van den Bunt, G.; Dierikx, C.M.; Bonten, M.J.M.; Bootsma, M.C.J.; Schmitt, H.; Hald, T.; Evers, E.G.; et al. Attributable sources of community-acquired carriage of Escherichia coli containing beta-lactam antibiotic resistance genes: A population-based modelling study. Lancet Planet. Health 2019, 3, e357–e369. [Google Scholar] [CrossRef] [Green Version]
- Friese, A.; Schulz, J.; Laube, H.; von Salviati, C.; Hartung, J.; Roesler, U. Faecal occurrence and emissions of livestock-associated methicillin-resistant Staphylococcus aureus (laMRSA) and ESBL/AmpC-producing E. coli from animal farms in Germany. Berl. Muench. Tieraerztl. Wochenschr. 2013, 126, 175–180. [Google Scholar]
- Huijbers, P.M.; Graat, E.A.; Haenen, A.P.; van Santen, M.G.; van Essen-Zandbergen, A.; Mevius, D.J.; van Duijkeren, E.; van Hoek, A.H. Extended-spectrum and AmpC beta-lactamase-producing Escherichia coli in broilers and people living and/or working on broiler farms: Prevalence, risk factors and molecular characteristics. J. Antimicrob. Chemother. 2014, 69, 2669–2675. [Google Scholar] [CrossRef] [PubMed]
- Laube, H.; Friese, A.; von Salviati, C.; Guerra, B.; Käsbohrer, A.; Kreienbrock, L.; Roesler, U. Longitudinal monitoring of extended-spectrum-beta-lactamase/AmpC-producing Escherichia coli at German broiler chicken fattening farms. Appl. Environ. Microbiol. 2013, 79, 4815–4820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daehre, K.; Projahn, M.; Semmler, T.; Roesler, U.; Friese, A. Extended-Spectrum Beta-Lactamase-/AmpC Beta-Lactamase-Producing Enterobacteriaceae in Broiler Farms: Transmission Dynamics at Farm Level. Microb. Drug Resist. 2018, 24, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Laube, H.; Friese, A.; von Salviati, C.; Guerra, B.; Rosler, U. Transmission of ESBL/AmpC-producing Escherichia coli from broiler chicken farms to surrounding areas. Vet. Microbiol. 2014, 172, 519–527. [Google Scholar] [CrossRef]
- Luyckx, K.; Dewulf, J.; Van Weyenberg, S.; Herman, L.; Zoons, J.; Vervaet, E.; Heyndrickx, M.; De Reu, K. Comparison of sampling procedures and microbiological and non-microbiological parameters to evaluate cleaning and disinfection in broiler houses. Poult. Sci. 2015, 94, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, K.Y.; Van Weyenberg, S.; Dewulf, J.; Herman, L.; Zoons, J.; Vervaet, E.; Heyndrickx, M.; De Reu, K. On-farm comparisons of different cleaning protocols in broiler houses. Poult. Sci. 2015, 94, 1986–1993. [Google Scholar] [CrossRef]
- Gilbert, P.; McBain, A.J. Potential impact of increased use of biocides in consumer products on prevalence of antibiotic resistance. Clin. Microbiol. Rev. 2003, 16, 189–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SCENIHR Opinion on the Assessment of the Antibiotic Resistance Effects of Biocides. Available online: https://ec.europa.eu/health/archive/ph_risk/committees/04_scenihr/docs/scenihr_o_021.pdf (accessed on 21 January 2021).
- Kanjee, U.; Houry, W.A. Mechanisms of acid resistance in Escherichia coli. Annu. Rev. Microbiol. 2013, 67, 65–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kummerle, N.; Feucht, H.H.; Kaulfers, P.M. Plasmid-mediated formaldehyde resistance in Escherichia coli: Characterization of resistance gene. Antimicrob. Agents Chemother. 1996, 40, 2276–2279. [Google Scholar] [CrossRef] [Green Version]
- Bas, S.; Kramer, M.; Stopar, D. Biofilm Surface Density Determines Biocide Effectiveness. Front. Microbiol. 2017, 8, 2443. [Google Scholar] [CrossRef]
- Denyer, S.P.; Maillard, J.Y. Cellular impermeability and uptake of biocides and antibiotics in Gram-negative bacteria. J. Appl. Microbiol. 2002, 92, 35S–45S. [Google Scholar] [CrossRef]
- Levy, S.B. Active efflux, a common mechanism for biocide and antibiotic resistance. J. Appl. Microbiol. 2002, 92, 65S–71S. [Google Scholar] [CrossRef] [PubMed]
- Bay, D.C.; Rommens, K.L.; Turner, R.J. Small multidrug resistance proteins: A multidrug transporter family that continues to grow. Biochim. Biophys. Acta 2008, 1778, 1814–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, D.C.; Turner, R.J. Diversity and evolution of the small multidrug resistance protein family. BMC Evol. Biol. 2009, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.; Bibi, E. MdfA, an Escherichia coli multidrug resistance protein with an extraordinarily broad spectrum of drug recognition. J. Bacteriol. 1997, 179, 2274–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.X.; Zhang, C.; Crow, R.R.; Thorpe, C.; Chen, H.; Kumar, S.; Tsuchiya, T.; Varela, M.F. SugE, a new member of the SMR family of transporters, contributes to antimicrobial resistance in Enterobacter cloacae. Antimicrob. Agents Chemother. 2011, 55, 3954–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fessler, A.T.; Schug, A.R.; Geber, F.; Scholtzek, A.D.; Merle, R.; Brombach, J.; Hensel, V.; Meurer, M.; Michael, G.B.; Reinhardt, M.; et al. Development and evaluation of a broth macrodilution method to determine the biocide susceptibility of bacteria. Vet. Microbiol. 2018, 223, 59–64. [Google Scholar] [CrossRef]
- Knapp, L.; Amezquita, A.; McClure, P.; Stewart, S.; Maillard, J.Y. Development of a protocol for predicting bacterial resistance to microbicides. Appl. Environ. Microbiol. 2015, 81, 2652–2659. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, I.; Oggioni, M.R.; Knight, D.; Curiao, T.; Coque, T.; Kalkanci, A.; Martinez, J.L. Evaluation of epidemiological cut-off values indicates that biocide resistant subpopulations are uncommon in natural isolates of clinically-relevant microorganisms. PLoS ONE 2014, 9, e86669. [Google Scholar] [CrossRef] [Green Version]
- Projahn, M.; Daehre, K.; Roesler, U.; Friese, A. Extended-Spectrum-Beta-Lactamase- and Plasmid-Encoded Cephamycinase-Producing Enterobacteria in the Broiler Hatchery as a Potential Mode of Pseudo-Vertical Transmission. Appl. Environ. Microbiol. 2017, 83, e02364-16. [Google Scholar] [CrossRef] [Green Version]
- Roschanski, N.; Fischer, J.; Guerra, B.; Roesler, U. Development of a multiplex real-time PCR for the rapid detection of the predominant beta-lactamase genes CTX-M, SHV, TEM and CIT-type AmpCs in Enterobacteriaceae. PLoS ONE 2014, 9, e100956. [Google Scholar] [CrossRef]
- Thomsen, M.C.; Ahrenfeldt, J.; Cisneros, J.L.; Jurtz, V.; Larsen, M.V.; Hasman, H.; Aarestrup, F.M.; Lund, O. A Bacterial Analysis Platform: An Integrated System for Analysing Bacterial Whole Genome Sequencing Data for Clinical Diagnostics and Surveillance. PLoS ONE 2016, 11, e0157718. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Knapp, L.; Rushton, L.; Stapleton, H.; Sass, A.; Stewart, S.; Amezquita, A.; McClure, P.; Mahenthiralingam, E.; Maillard, J.Y. The effect of cationic microbicide exposure against Burkholderia cepacia complex (Bcc); the use of Burkholderia lata strain 383 as a model bacterium. J. Appl. Microbiol. 2013, 115, 1117–1126. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roedel, A.; Dieckmann, R.; Brendebach, H.; Hammerl, J.A.; Kleta, S.; Noll, M.; Al Dahouk, S.; Vincze, S. Biocide tolerant Listeria monocytogenes isolates from German food production plants do not show cross-resistance to clinically relevant antibiotics. Appl. Environ. Microbiol. 2019, 85, e01253-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology 2005, 151, 2097–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronco, T.; Stegger, M.; Olsen, R.H.; Sekse, C.; Nordstoga, A.B.; Pohjanvirta, T.; Lilje, B.; Lyhs, U.; Andersen, P.S.; Pedersen, K. Spread of avian pathogenic Escherichia coli ST117 O78:H4 in Nordic broiler production. BMC Genom. 2017, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Kohler, C.D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef]
- Bourne, J.A.; Chong, W.L.; Gordon, D.M. Genetic structure, antimicrobial resistance and frequency of human associated Escherichia coli sequence types among faecal isolates from healthy dogs and cats living in Canberra, Australia. PLoS ONE 2019, 14, e0212867. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Wakeham, D.; Brouwers, H.J.; Cobbold, R.N.; Abraham, S.; Mollinger, J.L.; Johnson, J.R.; Chapman, T.A.; Gordon, D.M.; Barrs, V.R.; et al. Human-associated fluoroquinolone-resistant Escherichia coli clonal lineages, including ST354, isolated from canine feces and extraintestinal infections in Australia. Microbes Infect. 2015, 17, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Vangchhia, B.; Abraham, S.; Bell, J.M.; Collignon, P.; Gibson, J.S.; Ingram, P.R.; Johnson, J.R.; Kennedy, K.; Trott, D.J.; Turnidge, J.D.; et al. Phylogenetic diversity, antimicrobial susceptibility and virulence characteristics of phylogroup F Escherichia coli in Australia. Microbiology 2016, 162, 1904–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.; Meng, T.; Guastalli, E.A.; Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef]
- Cordoni, G.; Woodward, M.J.; Wu, H.; Alanazi, M.; Wallis, T.; La Ragione, R.M. Comparative genomics of European avian pathogenic E. coli (APEC). BMC Genom. 2016, 17, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dissanayake, D.R.; Octavia, S.; Lan, R. Population structure and virulence content of avian pathogenic Escherichia coli isolated from outbreaks in Sri Lanka. Vet. Microbiol. 2014, 168, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.; Lopez, C.; Herrera, A.; Viso, S.; Mamani, R.; Dhabi, G.; Alonso, M.P.; Blanco, M.; Blanco, J.E.; Blanco, J. Emerging avian pathogenic Escherichia coli strains belonging to clonal groups O111:H4-D-ST2085 and O111:H4-D-ST117 with high virulence-gene content and zoonotic potential. Vet. Microbiol. 2012, 156, 347–352. [Google Scholar] [CrossRef]
- Oteo, J.; Diestra, K.; Juan, C.; Bautista, V.; Novais, A.; Perez-Vazquez, M.; Moya, B.; Miro, E.; Coque, T.M.; Oliver, A.; et al. Extended-spectrum beta-lactamase-producing Escherichia coli in Spain belong to a large variety of multilocus sequence typing types, including ST10 complex/A, ST23 complex/A and ST131/B2. Int. J. Antimicrob. Agents 2009, 34, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Claeys, G.; Lontie, M.; Van Meensel, B.; Herman, L.; Haesebrouck, F.; et al. Characterization of extended-spectrum beta-lactamases produced by Escherichia coli isolated from hospitalized and nonhospitalized patients: Emergence of CTX-M-15-producing strains causing urinary tract infections. Microb. Drug Resist. 2010, 16, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Vincent, C.; Boerlin, P.; Daignault, D.; Dozois, C.M.; Dutil, L.; Galanakis, C.; Reid-Smith, R.J.; Tellier, P.P.; Tellis, P.A.; Ziebell, K.; et al. Food reservoir for Escherichia coli causing urinary tract infections. Emerg. Infect. Dis. 2010, 16, 88–95. [Google Scholar] [CrossRef]
- Heidemann Olsen, R.; Christensen, H.; Kabell, S.; Bisgaard, M. Characterization of prevalent bacterial pathogens associated with pododermatitis in table egg layers. Avian Pathol. 2018, 47, 281–285. [Google Scholar] [CrossRef]
- Mates, A.K.; Sayed, A.K.; Foster, J.W. Products of the Escherichia coli acid fitness island attenuate metabolite stress at extremely low pH and mediate a cell density-dependent acid resistance. J. Bacteriol. 2007, 189, 2759–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, J.W. Escherichia coli acid resistance: Tales of an amateur acidophile. Nat. Rev. Microbiol. 2004, 2, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, K.; Van Coillie, E.; Dewulf, J.; Van Weyenberg, S.; Herman, L.; Zoons, J.; Vervaet, E.; Heyndrickx, M.; De Reu, K. Identification and biocide susceptibility of dominant bacteria after cleaning and disinfection of broiler houses. Poult. Sci. 2016. [Google Scholar] [CrossRef]
- Schwaiger, K.; Bauer, J.; Holzel, C.S. Selection and persistence of antimicrobial-resistant Escherichia coli including extended-spectrum beta-lactamase producers in different poultry flocks on one chicken farm. Microb. Drug Resist. 2013, 19, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Poole, T.L.; Brichta-Harhay, D.M.; Anderson, R.C.; Bischoff, K.M.; Hernandez, C.A.; Bono, J.L.; Arthur, T.M.; Nagaraja, T.G.; Crippen, T.L.; et al. Disinfectant and antibiotic susceptibility profiles of Escherichia coli O157:H7 strains from cattle carcasses, feces, and hides and ground beef from the United States. J. Food Prot. 2013, 76, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Deus, D.; Krischek, C.; Pfeifer, Y.; Sharifi, A.R.; Fiegen, U.; Reich, F.; Klein, G.; Kehrenberg, C. Comparative analysis of the susceptibility to biocides and heavy metals of extended-spectrum beta-lactamase-producing Escherichia coli isolates of human and avian origin, Germany. Diagn. Microbiol. Infect. Dis. 2017, 88, 88–92. [Google Scholar] [CrossRef]
- Maertens, H.; De Reu, K.; Meyer, E.; Van Coillie, E.; Dewulf, J. Limited association between disinfectant use and either antibiotic or disinfectant susceptibility of Escherichia coli in both poultry and pig husbandry. BMC Vet. Res. 2019, 15, 310. [Google Scholar] [CrossRef] [Green Version]
- Oosterik, L.H.; Peeters, L.; Mutuku, I.; Goddeeris, B.M.; Butaye, P. Susceptibility of avian pathogenic Escherichia coli from laying hens in Belgium to antibiotics and disinfectants and integron prevalence. Avian Dis. 2014, 58, 271–278. [Google Scholar] [CrossRef]
- Sheridan, À.; Lenahan, M.; Duffy, G.; Fanning, S.; Burgess, C. The potential for biocide tolerance in Escherichia coli and its impact on the response to food processing stresses. Food Control 2012, 26, 98–106. [Google Scholar] [CrossRef]
- Schwaiger, K.; Harms, K.S.; Bischoff, M.; Preikschat, P.; Mölle, G.; Bauer-Unkauf, I.; Lindorfer, S.; Thalhammer, S.; Bauer, J.; Hölzel, C.S. Insusceptibility to disinfectants in bacteria from animals, food and humans-is there a link to antimicrobial resistance? Front. Microbiol. 2014, 5, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; He, X.; Meng, Y.; Guo, L.; Long, M.; Yu, H.; Li, B.; Fan, L.; Liu, S.; Wang, H.; et al. Antibiotic and Disinfectant Resistance of Escherichia coli Isolated from Retail Meats in Sichuan, China. Microb. Drug Resist. 2016, 22, 80–87. [Google Scholar] [CrossRef]
- Halstead, F.D.; Rauf, M.; Moiemen, N.S.; Bamford, A.; Wearn, C.M.; Fraise, A.P.; Lund, P.A.; Oppenheim, B.A.; Webber, M.A. The Antibacterial Activity of Acetic Acid against Biofilm-Producing Pathogens of Relevance to Burns Patients. PLoS ONE 2015, 10, e0136190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottini, C.B. Evaluation of Antimicrobials against Multi-Strain Cocktails of Salmonella, Escherichia coli O157:H7 and Listeria monocytogenes Using a Kinetic Growth Inhibition Assay. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 2012. [Google Scholar]
- Lesser, K.H. p-Chloro-m-cresol. In Cosmetic and Drug Preservation Principle and Practice; Kabara, J.J., Ed.; Marcel Dekker: New York, NY, USA, 1984; pp. 683–684. [Google Scholar]
- Paulus, W. Phenolics. In Directory of Microbicides for the Protection of Materials—A Handbook; Paulus, W., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; p. 534. [Google Scholar]
- Slipski, C.J.; Jamieson, T.R.; Lam, A.; Shing, V.L.; Bell, K.; Zhanel, G.G.; Bay, D.C. Plasmid transmitted small multidrug resistant (SMR) efflux pumps differ in gene regulation and enhance tolerance to quaternary ammonium compounds (QAC) when grown as biofilms. bioRxiv 2019, 768630. [Google Scholar] [CrossRef]
- Chen, N.H.; Djoko, K.Y.; Veyrier, F.J.; McEwan, A.G. Formaldehyde Stress Responses in Bacterial Pathogens. Front. Microbiol. 2016, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Kaulfers, P.-M.; Brandt, D. Isolation of a conjugative plasmid in Escherichia coli determining formaldehyde resistance. FEMS Microbiol. Lett. 1987, 43, 161–163. [Google Scholar] [CrossRef]
- Kaulfers, P.-M.; Marquardt, A. Demonstration of formaldehyde dehydrogenase activity in formaldehyde-resistant Enterobacteriaceae. FEMS Microbiol. Lett. 1991, 79, 335–338. [Google Scholar] [CrossRef]
- Dorsey, C.W.; Actis, L.A. Analysis of pVU3695, a plasmid encoding glutathione-dependent formaldehyde dehydrogenase activity and formaldehyde resistance in the Escherichia coli VU3695 clinical strain. Plasmid 2004, 51, 116–126. [Google Scholar] [CrossRef]
- Chung, Y.J.; Saier, M.H., Jr. Overexpression of the Escherichia coli sugE gene confers resistance to a narrow range of quaternary ammonium compounds. J. Bacteriol. 2002, 184, 2543–2545. [Google Scholar] [CrossRef] [Green Version]
- Lewinson, O.; Adler, J.; Poelarends, G.J.; Mazurkiewicz, P.; Driessen, A.J.M.; Bibi, E. The Escherichia coli multidrug transporter MdfA catalyzes both electrogenic and electroneutral transport reactions. Proc. Natl. Acad. Sci. USA 2003, 100, 1667–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, K. Efflux-mediated antimicrobial resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotem, D.; Schuldiner, S. EmrE, a Multidrug Transporter from Escherichia coli, Transports Monovalent and Divalent Substrates with the Same Stoichiometry. J. Biol. Chem. 2004, 279, 48787–48793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ye, L.; Kromann, S.; Meng, H. Occurrence of Extended-Spectrum β-Lactamases, Plasmid-Mediated Quinolone Resistance, and Disinfectant Resistance Genes in Escherichia coli Isolated from Ready-To-Eat Meat Products. Foodborne Pathog. Dis. 2016, 14, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Meng, J.; McDermott, P.F.; Wang, F.; Yang, Q.; Cao, G.; Hoffmann, M.; Zhao, S. Presence of disinfectant resistance genes in Escherichia coli isolated from retail meats in the USA. J. Antimicrob. Chemother. 2014, 69, 2644–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slipski, C.J.; Zhanel, G.G.; Bay, D.C. Biocide Selective TolC-Independent Efflux Pumps in Enterobacteriaceae. J. Membr. Biol. 2018, 251, 15–33. [Google Scholar] [CrossRef]
- Wieland, N.; Boss, J.; Lettmann, S.; Fritz, B.; Schwaiger, K.; Bauer, J.; Hölzel, C.S. Susceptibility to disinfectants in antimicrobial-resistant and -susceptible isolates of Escherichia coli, Enterococcus faecalis and Enterococcus faecium from poultry–ESBL/AmpC-phenotype of E. coli is not associated with resistance to a quaternary ammonium compound, DDAC. J. Appl. Microbiol. 2017, 122, 1508–1517. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Littlejohn, T.G.; Radstrom, P.; Sundstrom, L.; Skold, O.; Swedberg, G.; Skurray, R.A. The 3’ conserved segment of integrons contains a gene associated with multidrug resistance to antiseptics and disinfectants. Antimicrob. Agents Chemother. 1993, 37, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, R.; Sandle, T. A review on biocide reduced susceptibility due to plasmid-borne antiseptic-resistant genes-special notes on pharmaceutical environmental isolates. J. Appl. Microbiol. 2019, 126, 1011–1022. [Google Scholar] [CrossRef] [Green Version]
- Kücken, D.; Feucht, H.; Kaulfers, P. Association of qacE and qacEDelta1 with multiple resistance to antibiotics and antiseptics in clinical isolates of Gram-negative bacteria. FEMS Microbiol. Lett. 2000, 183, 95–98. [Google Scholar] [CrossRef]
- Bundesamt für Verbraucherschutz und Lebensmittelsicherheit (BVL). BVL-Report 12.2 Berichte zur Lebensmittelsicherheit Zoonosen-Monitoring 2016. Available online: https://www.bvl.bund.de/SharedDocs/Berichte/03_Zoonosen_Monitoring/2016_zoonosen_monitoring_bericht.pdf?__blob=publicationFile&v=5 (accessed on 29 November 2019).
- World Health Organization. Critically Important Antimicrobials for Human Medicine, 6th Revision; World Health Organization: Geneva, Switzerland, 2019; Licence: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Davies, J.; Wright, G.D. Bacterial resistance to aminoglycoside antibiotics. Trends Microbiol. 1997, 5, 234–240. [Google Scholar] [CrossRef]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of Resistance to Aminoglycoside Antibiotics: Overview and Perspectives. Medchemcomm 2016, 7, 11–27. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; CLSI: Wayne, PA, USA, 2019. [Google Scholar]
- European Medicines Agency. Reflection Paper on Use of Aminoglycosides in Animals in the European Union: Development of Resistance And Impact on Human and Animal Health; European Medicines Agency: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Soumet, C.F.; Legrandois, E.P.; Maris, P. Resistance to phenicol compounds following adaptation to quaternary ammonium compounds in Escherichia coli. Vet. Microbiol. 2012, 158, 147–152. [Google Scholar] [CrossRef]
- Wand, M.E.; Bock, L.J.; Bonney, L.C.; Sutton, J.M. Mechanisms of Increased Resistance to Chlorhexidine and Cross-Resistance to Colistin following Exposure of Klebsiella pneumoniae Clinical Isolates to Chlorhexidine. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, D.M.; Xu, W.; Loewen, P.C.; Zhanel, G.G.; Kumar, A. Triclosan can select for an AdeIJK-overexpressing mutant of Acinetobacter baumannii ATCC 17978 that displays reduced susceptibility to multiple antibiotics. Antimicrob. Agents Chemother. 2014, 58, 6424–6431. [Google Scholar] [CrossRef] [Green Version]
- Wales, A.D.; Davies, R.H. Co-Selection of Resistance to Antibiotics, Biocides and Heavy Metals, and Its Relevance to Foodborne Pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef] [Green Version]
- Kazama, H.; Hamashima, H.; Sasatsu, M.; Arai, T. Distribution of the antiseptic-resistance genes qacE and qacE delta 1 in gram-negative bacteria. FEMS Microbiol. Lett. 1998, 159, 173–178. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G. Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC Genom. 2015, 16, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Mo, S.S.; Slettemeas, J.S.; Berg, E.S.; Norstrom, M.; Sunde, M. Plasmid and Host Strain Characteristics of Escherichia coli Resistant to Extended-Spectrum Cephalosporins in the Norwegian Broiler Production. PLoS ONE 2016, 11, e0154019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porres-Osante, N.; Saenz, Y.; Somalo, S.; Torres, C. Characterization of Beta-lactamases in Faecal Enterobacteriaceae Recovered from Healthy Humans in Spain: Focusing on AmpC Polymorphisms. Microb. Ecol. 2015, 70, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, S.N.; Carattoli, A.; Schwendener, S.; Collaud, A.; Endimiani, A.; Perreten, V. Plasmids Carrying blaCMY-2/4 in Escherichia coli from Poultry, Poultry Meat, and Humans Belong to a Novel IncK Subgroup Designated IncK2. Front. Microbiol. 2017, 8, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietsch, M.; Irrgang, A.; Roschanski, N.; Brenner Michael, G.; Hamprecht, A.; Rieber, H.; Käsbohrer, A.; Schwarz, S.; Rösler, U.; Kreienbrock, L.; et al. Whole genome analyses of CMY-2-producing Escherichia coli isolates from humans, animals and food in Germany. BMC Genom. 2018, 19, 601. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roedel, A.; Vincze, S.; Projahn, M.; Roesler, U.; Robé, C.; Hammerl, J.A.; Noll, M.; Al Dahouk, S.; Dieckmann, R. Genetic but No Phenotypic Associations between Biocide Tolerance and Antibiotic Resistance in Escherichia coli from German Broiler Fattening Farms. Microorganisms 2021, 9, 651. https://doi.org/10.3390/microorganisms9030651
Roedel A, Vincze S, Projahn M, Roesler U, Robé C, Hammerl JA, Noll M, Al Dahouk S, Dieckmann R. Genetic but No Phenotypic Associations between Biocide Tolerance and Antibiotic Resistance in Escherichia coli from German Broiler Fattening Farms. Microorganisms. 2021; 9(3):651. https://doi.org/10.3390/microorganisms9030651
Chicago/Turabian StyleRoedel, Alice, Szilvia Vincze, Michaela Projahn, Uwe Roesler, Caroline Robé, Jens Andre Hammerl, Matthias Noll, Sascha Al Dahouk, and Ralf Dieckmann. 2021. "Genetic but No Phenotypic Associations between Biocide Tolerance and Antibiotic Resistance in Escherichia coli from German Broiler Fattening Farms" Microorganisms 9, no. 3: 651. https://doi.org/10.3390/microorganisms9030651