
Exogenous Probiotics Improve Fermentation Quality, Microflora Phenotypes, and Trophic Modes of Fermented Vegetable Waste for Animal Feed

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Materials and Additives
2.2. Fermented Feed Set-Up and Sampling
2.3. Analytical Produce
2.4. Analyses of the Physicochemical Properties
2.5. DNA Extraction and MiSeq Sequencing
2.6. Bioinformatics Analyses
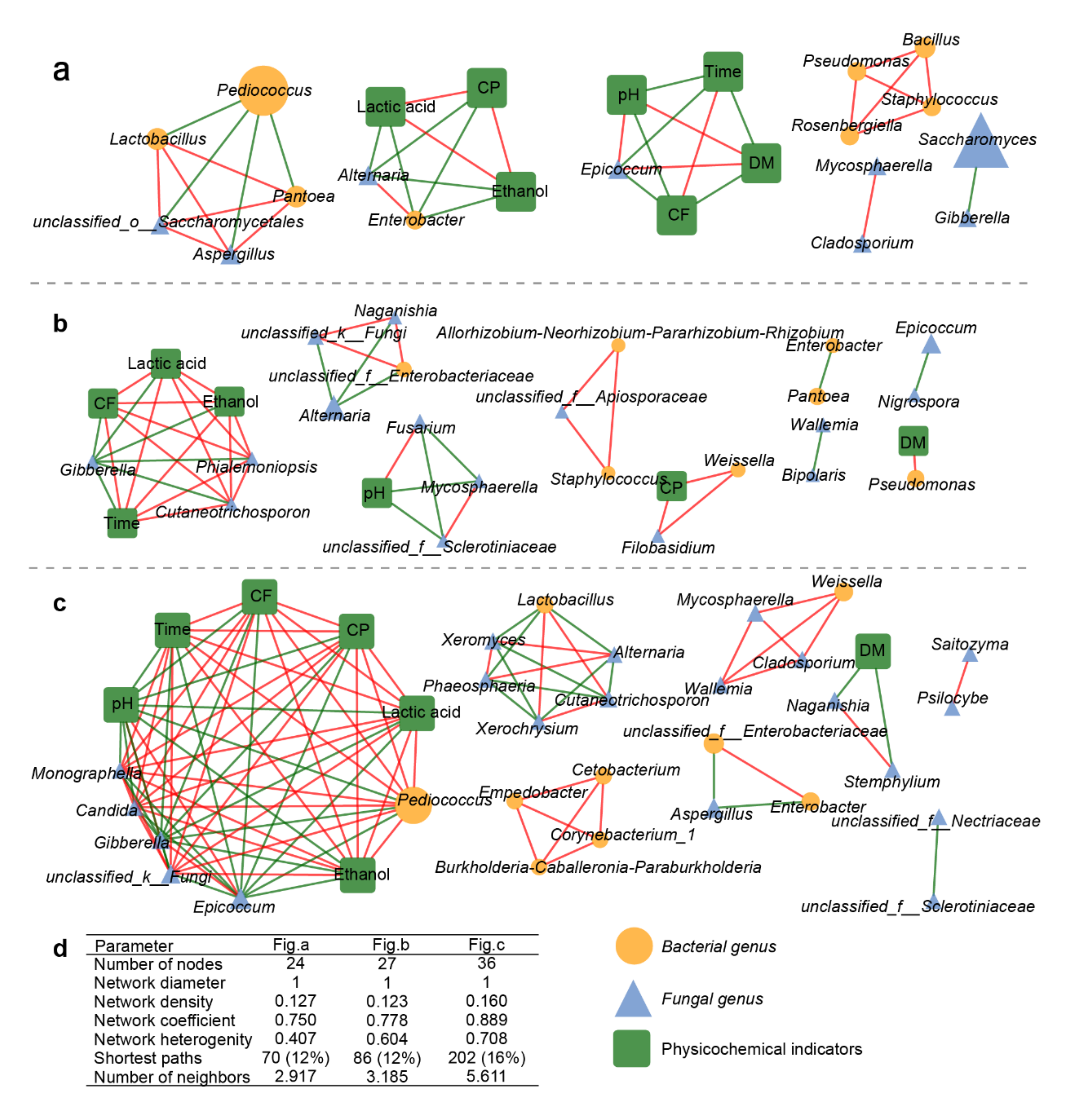
2.7. Correlation between Microbial Genus and Physicochemical Indexes
2.8. Statistical Analysis
3. Results and Discussion
3.1. Effects on the Physicochemical Properties
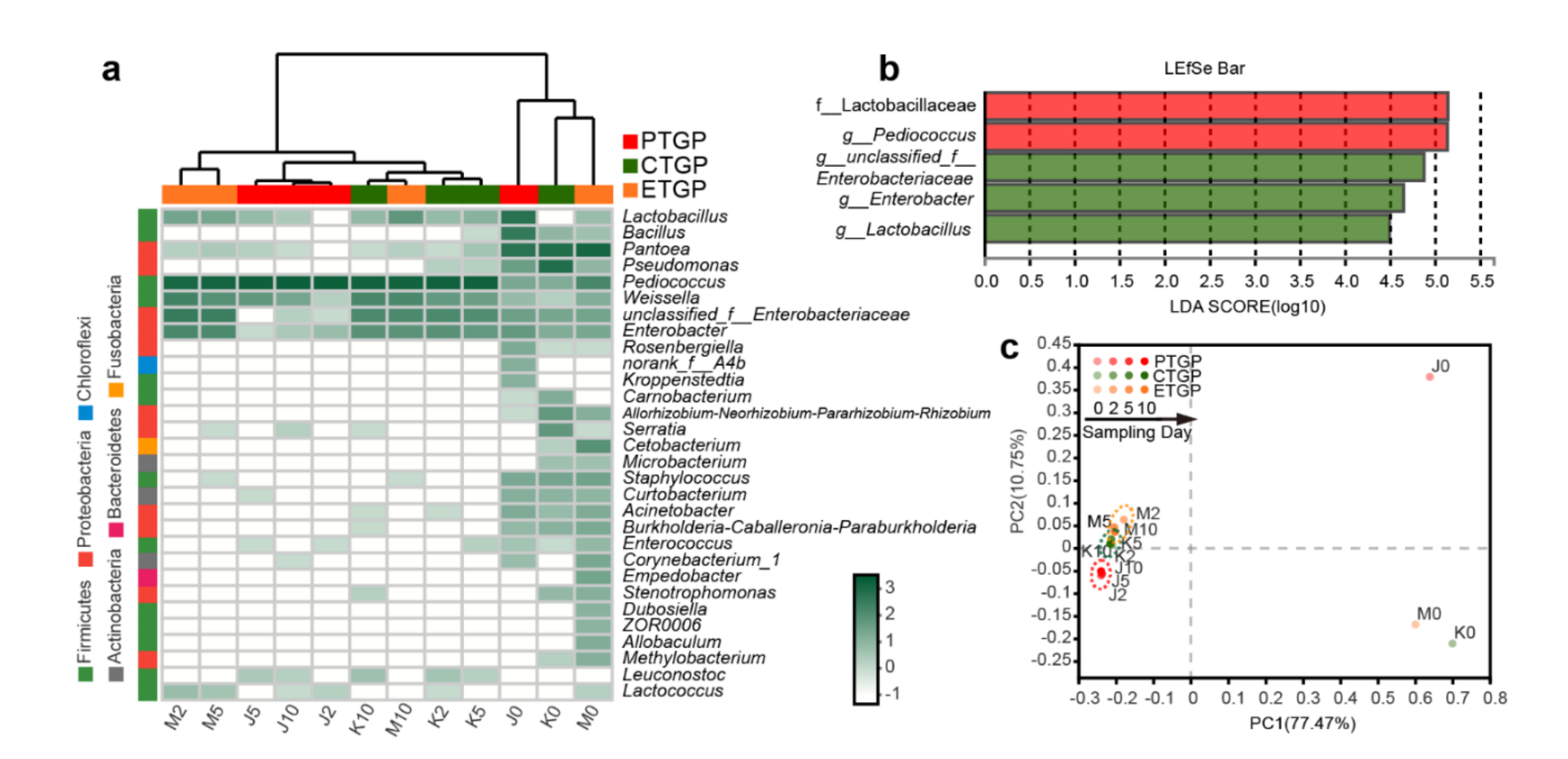
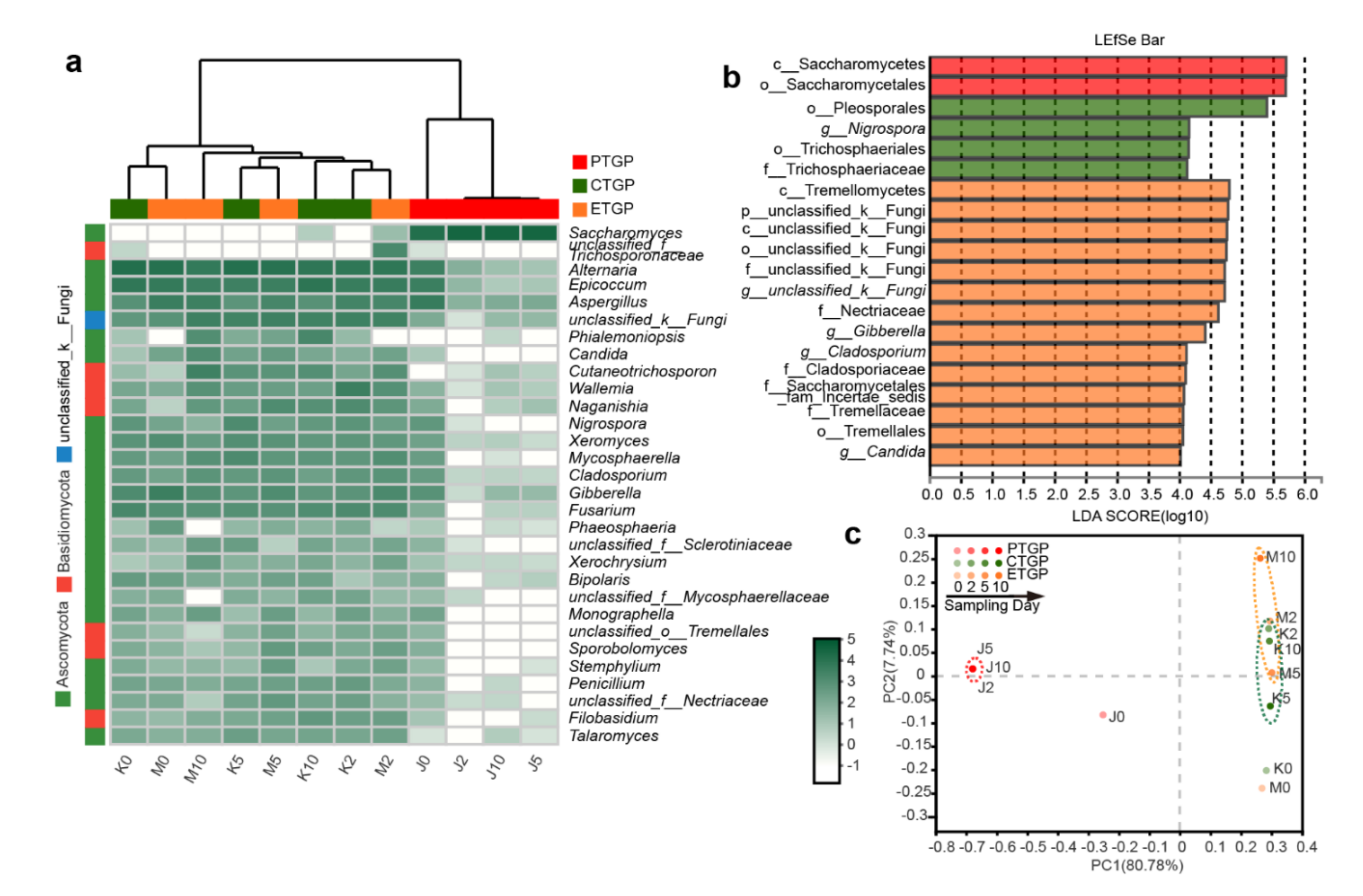
3.2. Effects on the Microflora Diversity and Richness
3.3. Effects on the Microflora Succession
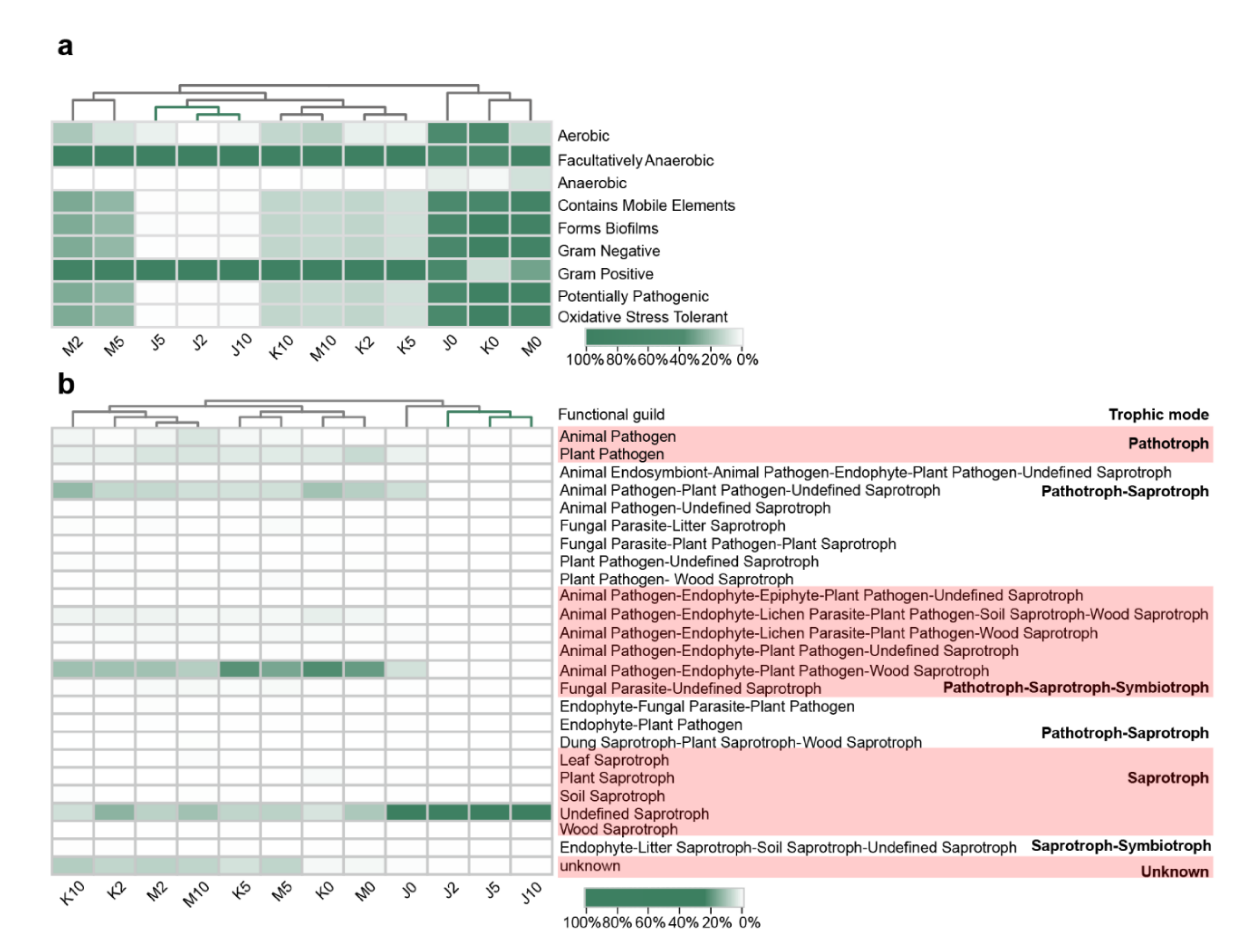
3.4. Effects on the Microflora Characteristics
3.4.1. Effects on the Phenotype of Bacterial Communities
3.4.2. Effects on the Trophic Modes of the Fungal Communities
3.5. Relationships between Microflora and Their Physicochemical Characteristics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, M.; Wang, C.; Wang, B.; Bai, X.; Gao, H.; Huang, Y. Enzymatic mechanism of organic nitrogen conversion and ammonia formation during vegetable waste composting using two amendments. Waste Manag. 2019, 95, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Joensuu, K.; Hartikainen, H.; Karppinen, S.; Jaakkonen, A.K.; Kuoppa-Aho, M. Developing the collection of statistical food waste data on the primary production of fruit and vegetables. Environ. Sci. Pollut. Res. Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus plantarum additive and temperature on the ensiling quality and microbial community dynamics of cauliflower leaf silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, R.; Thoma, G.; Matlock, M.D. Environmental sustainability of fruit and vegetable production supply chains in the face of climate change: A review. Sci. Total Environ. 2019, 650, 2863–2879. [Google Scholar] [CrossRef]
- Wolna-Maruwka, A.; Dach, J.; Rafaela, C.; Czekala, W.; Niewiadomska, A.; Janczak, D.; Budka, A. An effective method of utilizing vegetable waste in the form of carriers for Trichoderma strains with phytosanitary properties. Sci. Total Environ. 2019, 671, 795–804. [Google Scholar] [CrossRef]
- National Bureau of Statistics of People’s Republic of China. China Statistical Yearbook; China Statistics Press: Beijing, China, 2019.
- Viaene, J.; Agneessens, L.; Capito, C.; Ameloot, N.; Reubens, B.; Willekens, K.; Vandecasteele, B.; De Neve, S. Co-ensiling, co-composting and anaerobic co-digestion of vegetable crop residues: Product stability and effect on soil carbon and nitrogen dynamics. Sci. Hortic. 2017, 220, 214–225. [Google Scholar] [CrossRef]
- Ghinea, C.; Apostol, L.C.; Prisacaru, A.E.; Leahu, A. Development of a model for food waste composting. Environ. Sci. Pollut. Res. Int. 2019, 26, 4056–4069. [Google Scholar] [CrossRef] [PubMed]
- Halmemies-Beauchet-Filleau, A.; Rinne, M.; Lamminen, M.; Mapato, C.; Ampapon, T.; Wanapat, M.; Vanhatalo, A. Review: Alternative and novel feeds for ruminants: Nutritive value, product quality and environmental aspects. Animal 2018, 12, S295–S309. [Google Scholar] [CrossRef] [Green Version]
- San Martin, D.; Orive, M.; Martinez, E.; Inarra, B.; Ramos, S.; Gonzalez, N.; Guinea de Salas, A.; Vazquez, L.A.; Zufia, J. Multi-criteria assessment of the viability of valorising vegetable by-products from the distribution as secondary raw material for animal feed. Environ. Sci. Pollut. Res. Int. 2020. [Google Scholar] [CrossRef]
- Huang, L.; Ren, P.; Ouyang, Z.; Wei, T.; Kong, X.; Li, T.; Yin, Y.; He, S.; Yang, C.; He, Q. Effect of fermented feed on growth performance, holistic metabolism and fecal microbiota in weanling piglets. Anim. Feed Sci. Technol. 2020, 266. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Y.; Lu, Z.; Wang, Y. Solid-state fermentation of corn-soybean meal mixed feed with Bacillus subtilis and Enterococcus faecium for degrading antinutritional factors and enhancing nutritional value. J. Anim. Sci. Biotechnol. 2017, 8, 50. [Google Scholar] [CrossRef]
- Szmigiel, I.; Konkol, D.; Korczyński, M.; Łukaszewicz, M.; Krasowska, A. Changes in the Microbial Composition of the Cecum and Histomorphometric Analysis of Its Epithelium in Broilers Fed with Feed Mixture Containing Fermented Rapeseed Meal. Microorganisms 2021, 9, 360. [Google Scholar] [CrossRef] [PubMed]
- Cotarlet, M.; Stanciuc, N.; Bahrim, G.E. Yarrowia lipolytica and Lactobacillus paracasei Solid State Fermentation as a Valuable Biotechnological Tool for the Pork Lard and Okara’s Biotransformation. Microorganisms 2020, 8, 1098. [Google Scholar] [CrossRef] [PubMed]
- Babini, E.; Taneyo-Saa, D.L.; Tassoni, A.; Ferri, M.; Kraft, A.; Gran-Heedfeld, J.; Bretz, K.; Roda, A.; Michelini, E.; Calabretta, M.M.; et al. Microbial Fermentation of Industrial Rice-Starch Byproduct as Valuable Source of Peptide Fractions with Health-Related Activity. Microorganisms 2020, 8, 986. [Google Scholar] [CrossRef]
- Dzuman, Z.; Zachariasova, M.; Lacina, O.; Veprikova, Z.; Slavikova, P.; Hajslova, J. A rugged high-throughput analytical approach for the determination and quantification of multiple mycotoxins in complex feed. Talanta 2014, 121, 263–272. [Google Scholar] [CrossRef]
- Wang, C.; Shi, C.; Zhang, Y.; Song, D.; Lu, Z.; Wang, Y. Microbiota in fermented feed and swine gut. Appl. Microbiol. Biotechnol. 2018, 102, 2941–2948. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Bacterial Dynamics of Wheat Silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef]
- Yeh, R.H.; Hsieh, C.W.; Chen, K.L. Screening lactic acid bacteria to manufacture two-stage fermented feed and pelleting to investigate the feeding effect on broilers. Poult. Sci. 2018, 97, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Masebinu, S.O.; Akinlabi, E.T.; Muzenda, E.; Aboyade, A.O.; Mbohwa, C. Experimental and feasibility assessment of biogas production by anaerobic digestion of fruit and vegetable waste from Joburg Market. Waste Manag. 2018, 75, 236–250. [Google Scholar] [CrossRef]
- Song, D.; Li, X.; Cheng, Y.; Wu, G.; Xiao, X.; Wang, F.; Wang, Y.; Lu, Z. Effects of supplementing sow diets with refermented sorghum dried distiller’s grains with solubles from late gestation to weaning on the performance of sows and progeny. J. Anim. Sci. 2017, 95. [Google Scholar] [CrossRef]
- Zhu, J.; Gao, M.; Zhang, R.; Sun, Z.; Wang, C.; Yang, F.; Huang, T.; Qu, S.; Zhao, L.; Li, Y.; et al. Effects of soybean meal fermented by L. plantarum, B. subtilis and S. cerevisieae on growth, immune function and intestinal morphology in weaned piglets. Microb. Cell. Fact. 2017, 16, 191. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.; Xie, Z.; Hu, L.; Chen, G.; Zhang, Z. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, Z.; Yu, Z. Pectin Degradation is an Important Determinant for Alfalfa Silage Fermentation through the Rescheduling of the Bacterial Community. Microorganisms 2020, 8, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, T.A.; Duniere, L.; Drouin, P.; Xu, S.; Wang, Y.; Munns, K.; Zaheer, R. Silage review: Using molecular approaches to define the microbial ecology of silage. J. Dairy Sci. 2018, 101, 4060–4074. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Song, Z.W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.R.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 133462. [Google Scholar] [CrossRef]
- Ali, N.; Wang, S.R.; Zhao, J.; Dong, Z.H.; Li, J.F.; Nazar, M.; Shao, T. Microbial diversity and fermentation profile of red clover silage inoculated with reconstituted indigenous and exogenous epiphytic microbiota. Bioresour. Technol. 2020, 314, 10. [Google Scholar] [CrossRef]
- Guan, H.; Shuai, Y.; Yan, Y.; Ran, Q.; Wang, X.; Li, D.; Cai, Y.; Zhang, X. Microbial Community and Fermentation Dynamics of Corn Silage Prepared with Heat-Resistant Lactic Acid Bacteria in a Hot Environment. Microorganisms 2020, 8, 719. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Liu, T.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Peng, Z. Effects of different simulated seasonal temperatures on the fermentation characteristics and microbial community diversities of the maize straw and cabbage waste co-ensiling system. Sci. Total Environ. 2020, 708, 135113. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, X.; Gu, Q.; Liang, M.; Mu, S.; Zhou, B.; Huang, F.; Lin, B.; Zou, C. Analysis of the correlation between bacteria and fungi in sugarcane tops silage prior to and after aerobic exposure. Bioresour. Technol. 2019, 291, 121835. [Google Scholar] [CrossRef]
- Du, G.; Feng, W.; Cai, H.; Ma, Z.; Liu, X.; Yuan, C.; Shi, J.; Zhang, B. Exogenous enzyme amendment accelerates maturity and changes microflora succession in horse and wildlife animal manure co-composting. Environ. Sci. Pollut. Res. Int. 2021. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinformatics 2003, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Lv, H.; Xing, Y.; Chen, X.; Zhang, Q. Intrinsic tannins affect ensiling characteristics and proteolysis of Neolamarckia cadamba leaf silage by largely altering bacterial community. Bioresour. Technol. 2020, 311, 123496. [Google Scholar] [CrossRef]
- Novinscak, A.; DeCoste, N.J.; Surette, C.; Filion, M. Characterization of bacterial and fungal communities in composted biosolids over a 2 year period using denaturing gradient gel electrophoresis. Can. J. Microbiol. 2009, 55, 375–387. [Google Scholar] [CrossRef]
- Nath, R.; Sargiary, P.; Borkakoty, B.; Parida, P. Cutaneotrichosporon (Trichosporon) debeurmannianum: A Rare Yeast Isolated from Blood and Urine Samples. Mycopathologia 2018, 183, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Rhim, H.; Park, J.Y.; Lee, D.J.; Han, J.I. Epicoccum nigrum-induced respiratory infection in a wild Eurasian scops owl (Otus scops). J. Vet. Med. Sci. 2019, 81, 1348–1350. [Google Scholar] [CrossRef]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef]
- Avila, C.L.S.; Carvalho, B.F. Silage fermentation-updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef]
- Xu, Y.; Dhaouadi, Y.; Stoodley, P.; Ren, D. Sensing the unreachable: Challenges and opportunities in biofilm detection. Curr. Opin. Biotechnol. 2020, 64, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, M.; Zapparoli, G. Occurrence and infection of Cladosporium, Fusarium, Epicoccum and Aureobasidium in withered rotten grapes during post-harvest dehydration. Anton. Leeuw. Int. J. G. 2015, 108, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Materials | DM (g kg−1) | CF (g kg−1 DM) | CP (g kg−1 DM) | ADF (g kg−1 DM) | NDF (g kg−1 DM) |
|---|---|---|---|---|---|
| Soybean meal | 887.3 ± 1.2 | 89.4 ± 0.3 | 446.9 ± 4.8 | 105.9 ± 5.8 | 251.0 ± 6.7 |
| Corn flour | 877.3 ± 1.9 | 52.9 ± 3.5 | 90.5 ± 6.1 | 11.8 ± 1.3 | 24.7 ± 2.0 |
| Wheat bran | 880.2 ± 1.5 | 88.2 ± 2.2 | 195.4 ± 6.1 | 100.7 ± 1.7 | 411.4 ± 1.0 |
| Cabbage waste | 59.4 ± 0.7 | 425 ± 2.6 | 220.0 ± 5.7 | 176.9 ± 7.8 | 233.6 ± 4.8 |
| Experimental Group | Formulations | Additive |
|---|---|---|
| CTGP | 15% Soybean meal + 15% Corn flour + 15% Wheat bran + 55% Cabbage waste | - |
| PTGP | CTGP | S. cerevisia 9.0 × 107 CFU g−1 FM B. subtilis 6.6 × 106 CFU g−1 FM L. plantarum 6.6 × 106 CFU g−1 FM |
| ETGP | CTGP | 0.5 wt.% FM of exogenous enzyme |
| Items | TR | Time (d) | Mean | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 5 | 10 | 15 | TR | TI | TR × TI | ||||
| DM (g kg−1) | PTGP | 376 ± 9.7 Aa | 334 ± 4.7 Bb | 299 ± 9.3 Bbc | 269 ± 7.4 Bc | 270 ± 15.2 Bc | 303.0 | 11.72 | *** | *** | *** |
| CTGP | 377 ± 5.2 Aa | 375 ± 13.4 Aa | 375 ± 9 Aa | 373 ± 12.4 Aa | 357 ± 14.3 Aa | 369.9 | |||||
| ETGP | 378 ± 14.3 Aa | 351 ± 6.7 Aba | 346 ± 17.9 Aa | 353 ± 6.3 Aa | 351 ± 3.4 Aa | 356.5 | |||||
| CP (g kg−1 DM) | PTGP | 233 ± 12.1 Ac | 275 ± 3.1 Bb | 301 ± 6.7 Aa | 291 ± 7.6 Aab | 292 ± 9.8 Aab | 275.6 | 7.34 | *** | *** | *** |
| CTGP | 232 ± 7.2 Ac | 245 ± 2.9 Babc | 241 ± 6.3 Bbc | 260 ± 4.2 Ba | 251 ± 7.6 Bab | 246.6 | |||||
| ETGP | 245 ± 7.9 Ab | 249 ± 6.3 Aab | 260 ± 6.9 Bab | 266 ± 3.1 Ba | 263 ± 8.3 Bab | 256.5 | |||||
| CF (g kg−1 DM) | PTGP | 103 ± 2.6 Ac | 121 ± 2.3 Cc | 148 ± 10.6 Bb | 166 ± 10.3 Bab | 183.7 ± 6.1 Ba | 147.3 | 8.28 | *** | *** | *** |
| CTGP | 110 ± 8.3 Ac | 128 ± 1.7 Bbc | 142 ± 9.3 Bb | 167 ± 3.6 Ba | 181.6 ± 10.9 Ba | 143.2 | |||||
| ETGP | 102 ± 3.8 Ac | 186 ± 2.7 Ab | 199 ± 9.5 Aab | 218 ± 17.8 Aa | 208.4 ± 0.8 Aab | 180.9 | |||||
| ADF (g kg−1 DM) | PTGP | 91.2 ± 4.3 Aa | 85.7 ± 4.0 Ba | 84.6 ± 0.6 Ca | 89.6 ± 2.5 Aa | 92.8 ± 3.5 Ba | 88.8 | 6.54 | *** | *** | *** |
| CTGP | 90.9 ± 4.0 Ab | 120 ± 4.1 Aa | 116.5 ± 0.8 Ba | 110 ± 20.2 Aab | 95.6 ± 2.4 Bab | 106.3 | |||||
| ETGP | 86.9 ± 2.5 Ab | 129 ± 3.7 Aa | 136.2 ± 6.1 Aa | 123 ± 9.2 Aa | 126.2 ± 5.3 Aa | 120.1 | |||||
| NDF (g kg−1 DM) | PTGP | 211 ± 9.1 Ac | 305 ± 9.3 Ca | 230 ± 7.2 Bbc | 235 ± 2.2 Cb | 224 ± 2.5 Bbc | 236.9 | 11.75 | *** | *** | *** |
| CTGP | 212 ± 5.4 Ac | 384 ± 11.1 Ba | 372 ± 8.2 Aa | 296 ± 2.2 Bb | 295 ± 15.1 Bb | 314.4 | |||||
| ETGP | 207 ± 5.6 Ab | 350 ± 2.5 Aa | 341 ± 14.6 Aa | 322 ± 12.8 Aa | 326 ± 29.0 Aa | 315.9 | |||||
| Items | TR | Time (d) | Mean | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 5 | 10 | 15 | TR | TI | TR × TI | ||||
| pH | PTGP | 6.20 ± 0.11 Aa | 5.00 ± 0.05 Ab | 4.44 ± 0.14 Ac | 4.32 ± 0.02 Ac | 4.34 ± 0.14 Ac | 4.76 | 0.07 | *** | *** | *** |
| CTGP | 6.22 ± 0.05 Aa | 3.96 ± 0.07 Bb | 3.89 ± 0.01 Bb | 3.89 ± 0.04 Bb | 3.88 ± 0.02 Bb | 4.23 | |||||
| ETGP | 6.14 ± 0.12 Aa | 4.07 ± 0.01 Bb | 3.93 ± 0.02 Bbc | 3.92 ± 0.02 Bc | 3.96 ± 0.01 Bc | 4.28 | |||||
| Lactic acid (g kg−1 DM) | PTGP | 0.0 ± 0.0 Cc | 36.2 ± 2.7 Bb | 63.4 ± 11.9 Aa | 52.7 ± 8.1 Bab | 39.9 ± 2.9 Bb | 36.4 | 4.43 | *** | *** | *** |
| CTGP | 2.5 ± 0.1 Bc | 75.5 ± 3.3 Ab | 79.6 ± 6.3 Aab | 82.2 ± 4.3 Aab | 88.0 ± 2.6 Aa | 64.0 | |||||
| ETGP | 4.1 ± 0.2 Ac | 68.9 ± 3.3 Ab | 83.0 ± 3.8 Aa | 87.1 ± 4.2 Aa | 89.7 ± 1.2 Aa | 66.5 | |||||
| Acetic acid (g kg−1 DM) | PTGP | 0.0 ± 0.0 Cc | 10.7 ± 0.8 Aab | 13.4 ± 4.3 Aa | 8.8 ± 1.5 Aab | 5.5 ± 1.6 Bb | 7.3 | 1.14 | *** | *** | *** |
| CTGP | 0.5 ± 0.0 Bb | 5.6 ± 0.7 Ba | 4.8 ± 0.9 Ba | 5.2 ± 0.3 Ba | 5.3 ± 0.9 Ba | 4.3 | |||||
| ETGP | 0.3 ± 0.1 Ac | 9.2 ± 0.5 Ab | 11.3 ± 0.8 ABa | 10.2 ± 0.6 Aab | 9.6 ± 0.6 Ab | 7.9 | |||||
| Butyric acid (g kg−1 DM) | PTGP | 0.0 ± 0.0 Ab | 0.9 ± 0.0 Aab | 1.7 ± 0.9 Aa | 0.7 ± 0.0 Aab | 0.5 ± 0.0 Aab | 0.7 | 0.21 | *** | *** | *** |
| CTGP | 0.0 ± 0.0 Ab | 0.4 ± 0.2 Bab | 0.4 ± 0.0 Aa | 0.6 ± 0.1 Aa | 0.7 ± 0.2 Aa | 0.4 | |||||
| ETGP | 0.0 ± 0.0 Ad | 0.5 ± 0.1 ABc | 0.7 ± 0.1 Ab | 0.7 ± 0.0 Ab | 0.9 ± 0.0 Aa | 0.5 | |||||
| Ethanol (g kg−1 DM) | PTGP | 0.0 ± 0.0 Ab | 55.5 ± 4.7 Ba | 66.3 ± 19 Ba | 62.7 ± 10.3 Ba | 39.2 ± 1.4 Ca | 41.9 | 5.08 | *** | *** | *** |
| CTGP | 0.0 ± 0.0 Ac | 1.9 ± 0.1 Bb | 1.9 ± 0.2 Bb | 21.5 ± 1.2 Ba | 20.3 ± 0.0 Ba | 7.8 | |||||
| ETGP | 0.0 ± 0.0 Ac | 1.4 ± 0.1 Abc | 2.9 ± 0.5 Ab | 19.1 ± 0.9 Aa | 1.6 ± 0.0 Ab | 4.5 | |||||
| NH4+/TN (%) | PTGP | 0.46 ± 0.02 Bc | 1.44 ± 0.21 Bb | 2.21 ± 0.39 Aab 2.45 ± 0.40 Ba | 1.90 ± 0.34 Bab | 1.90 ± 0.34 Bab | 1.78 | 0.23 | *** | *** | *** |
| CTGP | 0.79 ± 0.06 Ae | 1.36 ± 0.15 Bd | 2.30 ± 0.02 Ab | 3.27 ± 0.17 Aa | 1.87 ± 0.06 Bc | 2.00 | |||||
| ETGP | 0.53 ± 0.02 Bd | 2.01 ± 0.10 Ac | 2.39 ± 0.19 Abc 2.68 ± 0.34 ABb | 3.52 ± 0.14 Aa | 3.52 ± 0.14 Aa | 2.26 | |||||
| Item | TR | Time (d) | Mean | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 5 | 10 | 15 | TR | TI | TR × TI | ||||
| Formic acid (g kg−1 DM) | PTGP | 7.99 ± 0.29 Ac | 34.01 ± 2.1 Aa | 30.44 ± 3.69 Aa | 31.47 ± 1.73 Aa | 20.2 ± 2.96 ABb | 24.19 | 1.48 | *** | *** | *** |
| CTGP | 8.84 ± 0.44 Ab | 9.72 ± 1.22 Bb | 11.31 ± 1.01 Bb | 23.74 ± 0.54 Ba | 22.79 ± 1.78 Aa | 14.05 | |||||
| ETGP | 6.76 ± 0.29 Bc | 10.01 ± 0.82 Bb | 13.23 ± 1.41 Ba | 15.66 ± 0.98 Ca | 13.48 ± 0.08 Ba | 11.59 | |||||
| Propionic acid (g kg−1 DM) | PTGP | NA | NA | NA | NA | NA | - | - | - | - | - |
| CTGP | NA | NA | NA | NA | NA | - | |||||
| ETGP | NA | NA | NA | NA | NA | - | |||||
| Lactic acid/TOA (%) | PTGP | - | 44.25 | 58.20 | 56.26 | 60.37 | - | - | - | - | - |
| CTGP | - | 82.77 | 82.83 | 73.56 | 75.35 | - | |||||
| ETGP | - | 77.76 | 76.69 | 76.63 | 78.90 | - | |||||
| Acetic acid/TOA (%) | PTGP | - | 13.08 | 12.30 | 9.39 | 8.32 | - | - | - | - | - |
| CTGP | - | 6.14 | 4.99 | 4.65 | 4.54 | - | |||||
| ETGP | - | 10.38 | 10.44 | 8.97 | 8.44 | - | |||||
| Butyric acid/TOA (%) | PTGP | - | 1.10 | 1.56 | 0.75 | 0.76 | - | - | - | - | - |
| CTGP | - | 0.44 | 0.42 | 0.54 | 0.60 | - | |||||
| ETGP | - | 0.56 | 0.65 | 0.62 | 0.79 | ||||||
| Flieg’s score | PTGP | - | Good (80) | Very good (87) | Very good (86) | Very good (88) | - | - | - | - | - |
| CTGP | - | Very good (100) | Very good (100) | Very good (97) | Very good (100) | - | |||||
| ETGP | - | Very good (100) | Very good (100) | Very good (100) | Very good (100) | - | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, G.; Shi, J.; Zhang, J.; Ma, Z.; Liu, X.; Yuan, C.; Zhang, B.; Zhang, Z.; Harrison, M.D. Exogenous Probiotics Improve Fermentation Quality, Microflora Phenotypes, and Trophic Modes of Fermented Vegetable Waste for Animal Feed. Microorganisms 2021, 9, 644. https://doi.org/10.3390/microorganisms9030644
Du G, Shi J, Zhang J, Ma Z, Liu X, Yuan C, Zhang B, Zhang Z, Harrison MD. Exogenous Probiotics Improve Fermentation Quality, Microflora Phenotypes, and Trophic Modes of Fermented Vegetable Waste for Animal Feed. Microorganisms. 2021; 9(3):644. https://doi.org/10.3390/microorganisms9030644
Chicago/Turabian StyleDu, Guilin, Jiping Shi, Jingxian Zhang, Zhiguo Ma, Xiangcen Liu, Chenyang Yuan, Baoguo Zhang, Zhanying Zhang, and Mark D. Harrison. 2021. "Exogenous Probiotics Improve Fermentation Quality, Microflora Phenotypes, and Trophic Modes of Fermented Vegetable Waste for Animal Feed" Microorganisms 9, no. 3: 644. https://doi.org/10.3390/microorganisms9030644