
Apple Pomace and Performance, Intestinal Morphology and Microbiota of Weaned Piglets—A Weaning Strategy for Gut Health?

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Animals, Diets and Housing
2.1.2. Sampling of Feces
2.1.3. Sampling of Intestinal Tissues and Contents
2.2. Experimental Measurements
2.2.1. Chemical Analyses
2.2.2. Zootechnical Performance
2.2.3. Scoring of Piglet’s Fecal Consistency
2.2.4. Excreted Pathogens
2.2.5. Intestinal Morphology
2.2.6. Short Chain Fatty Acids (SCFA)
2.2.7. Composition and Richness of the Microbiota
2.3. Statistical Analyses
2.3.1. Parametric Tests
2.3.2. Non-Parametric Tests
3. Results
3.1. Feed Chemical Analyses
3.2. Zootechnical Performance
3.3. Scoring of Piglet’s Fecal Consistency and Excreted Pathogens
3.4. Intestinal Morphology
3.5. Short Chain Fatty Acids (SCFA)
3.6. Richness and Composition of the Microbiota
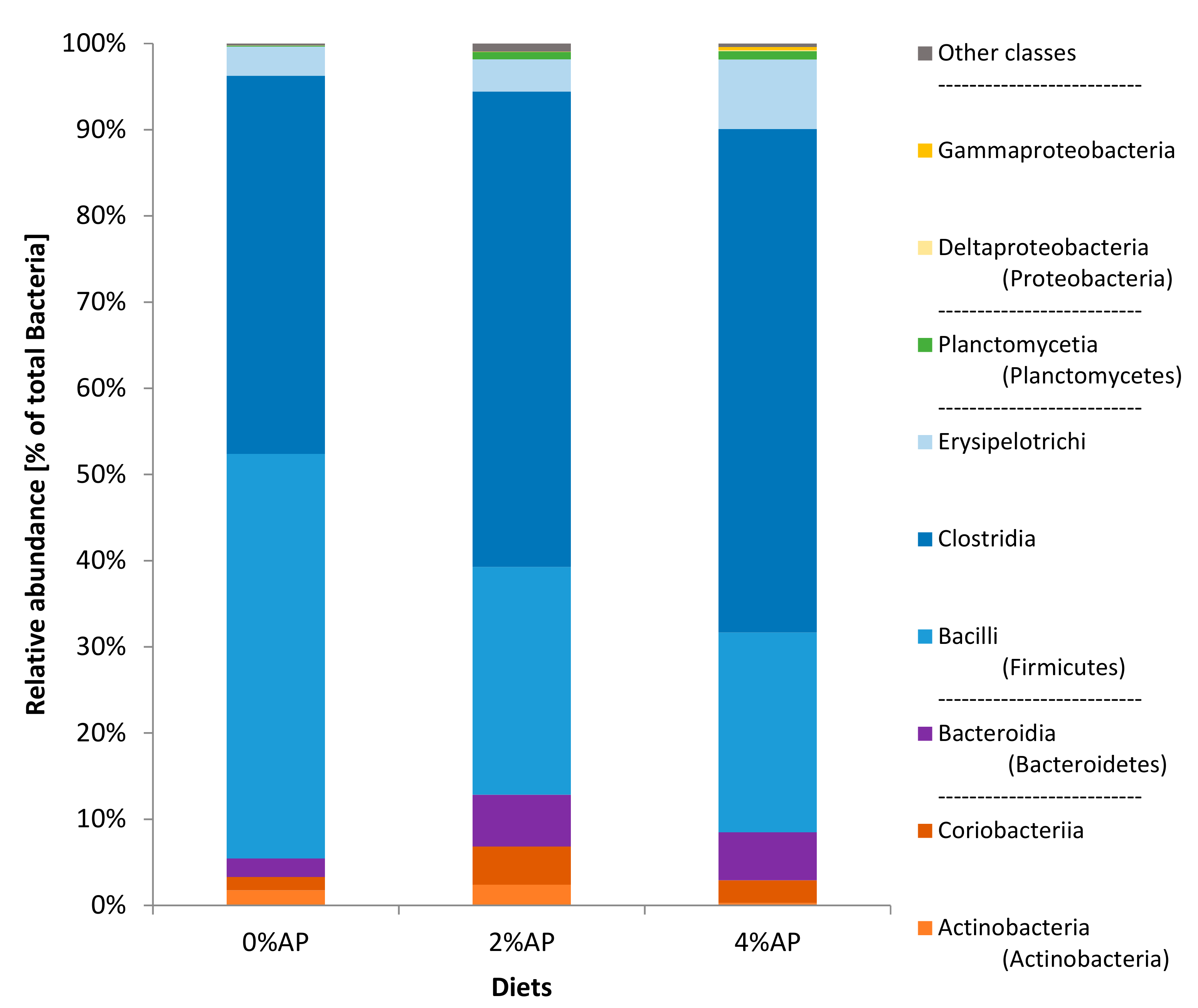
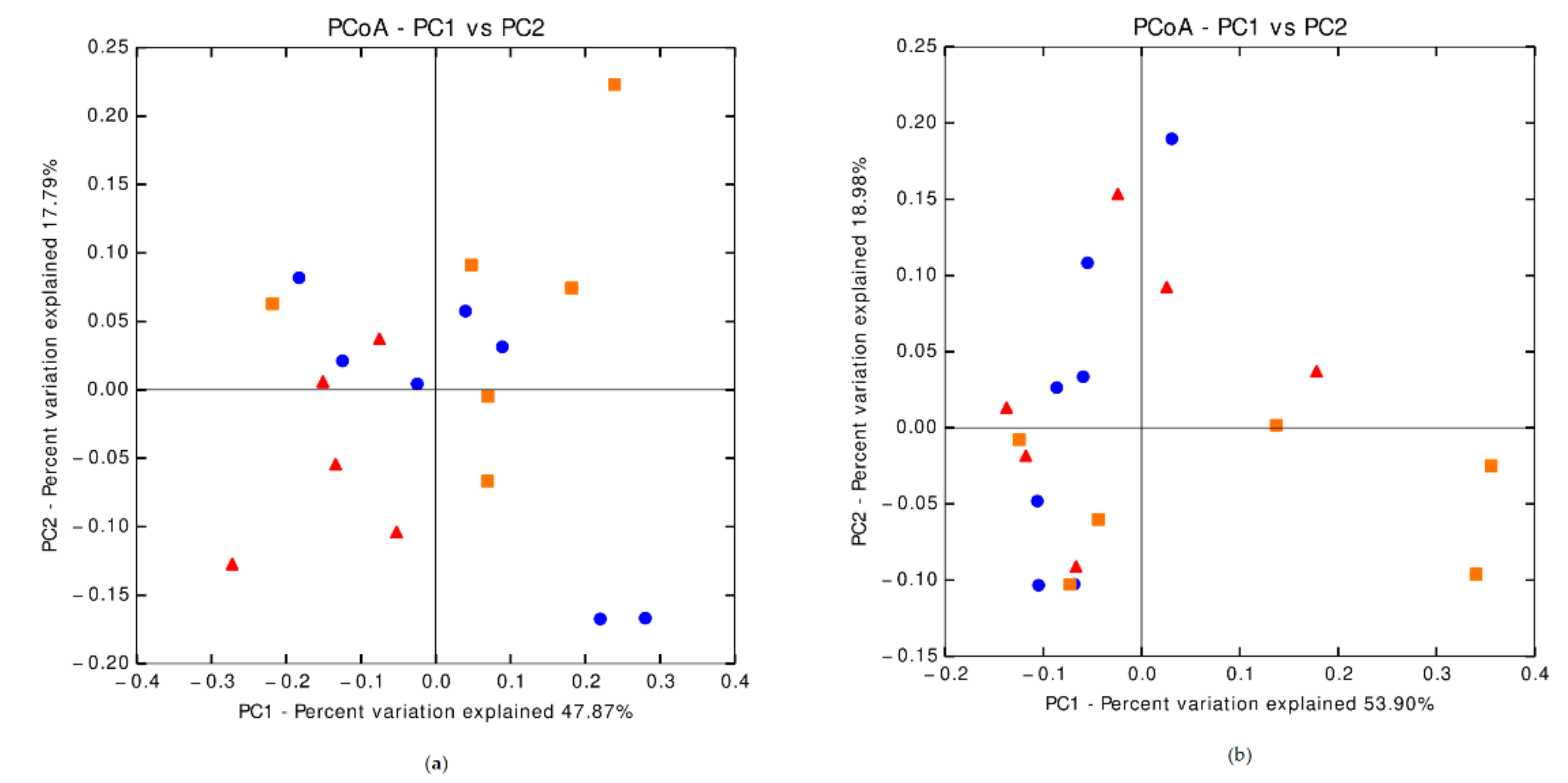
3.6.1. Fecal Microbiota Composition on Day 8 Post-Weaning
3.6.2. Fecal Microbiota Composition on Day 28 Post-Weaning
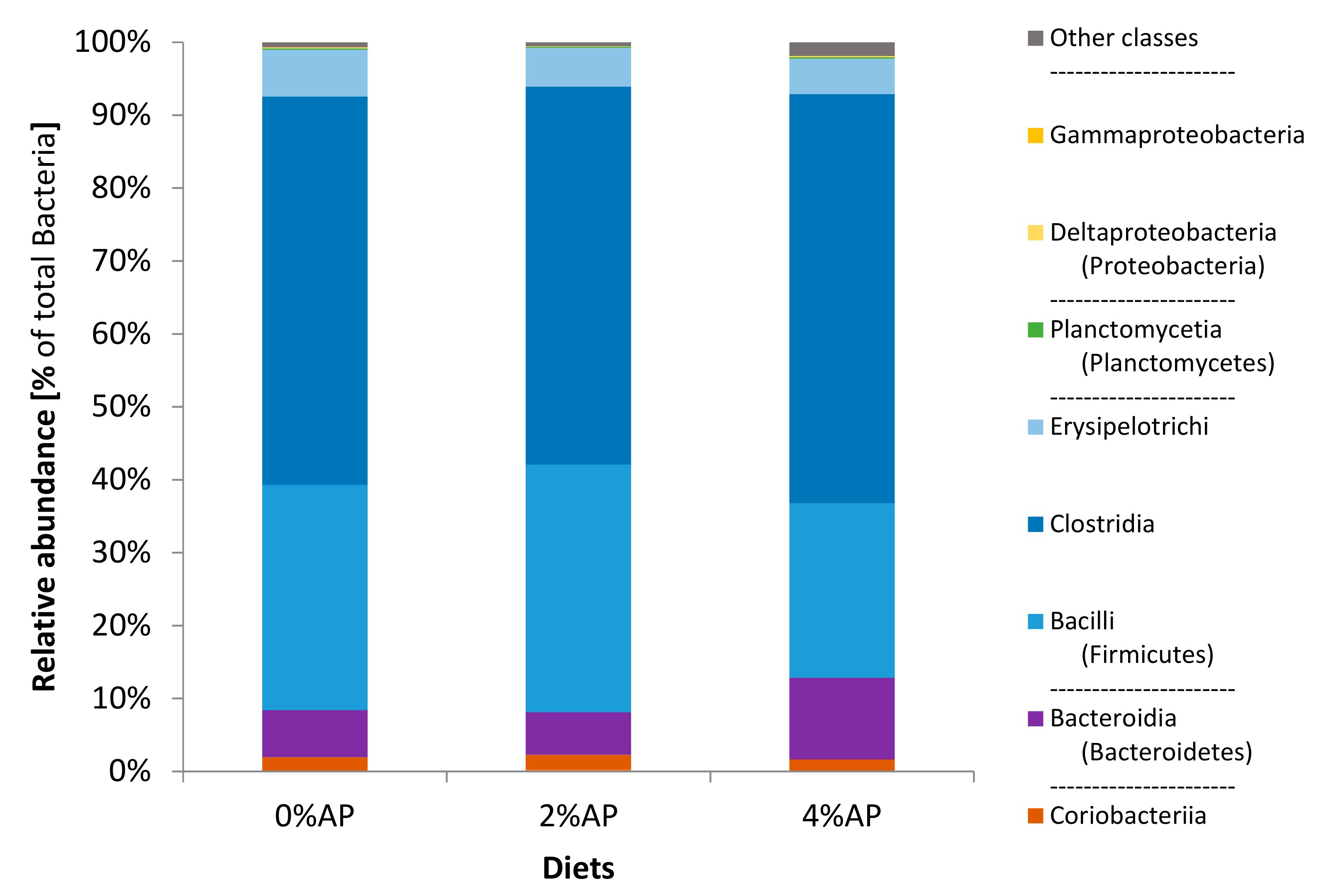
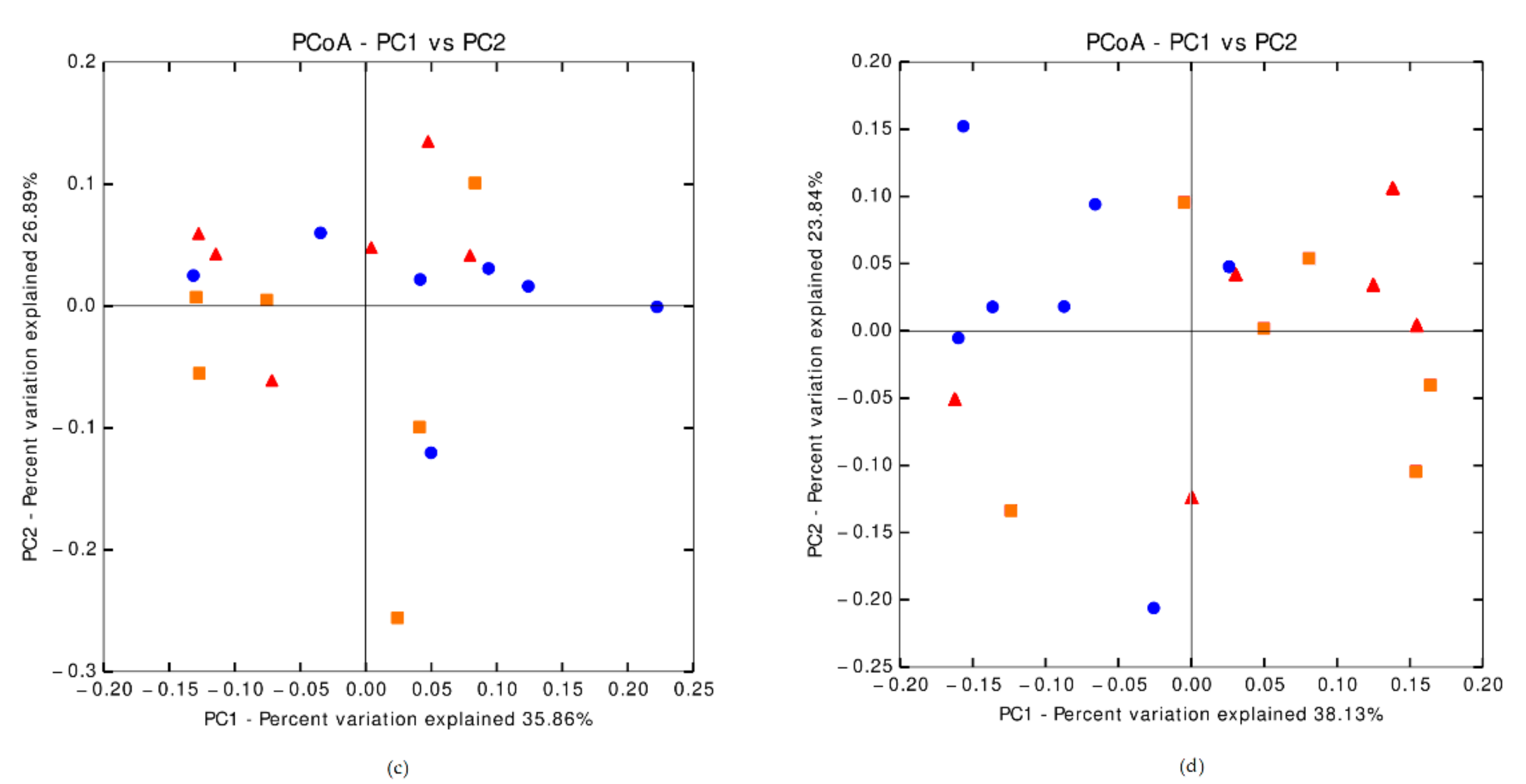
3.6.3. Cecal Microbiota Composition on Day 35 Post-Weaning
4. Discussion
4.1. Zootechnical Performance and Intestinal Morphology
4.2. Scoring of Piglet’s Fecal Consistency and Excreted Pathogens
4.3. Microbiota and SCFA in Feces and Cecum
4.4. AP as a Weaning Strategy for Gut Health
4.5. Emerging Concept from the Results
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0% AP | 2% AP | 4% AP | p-Value KW/FDR | |
|---|---|---|---|---|
| Feces on day 8 post-weaning (family) | ||||
| Coriobacteriaceae | 1.6% | 4.5% | 2.5% | 0.080/ns |
| Lactobacillaceae | 45.5% | 25.5% | 23.1% | 0.053/ns |
| Enterobacteriaceae | 0.0% | 0.1% | 0.4% | 0.020/ns |
| Feces on day 8 post-weaning (genus) | ||||
| Dorea | 1.3% a | 0.2% b | 0.7% ab | 0.024/ns |
| Slackia | 0.0% | 0.1% | 0.0% | 0.039 1/ns |
| Ruminococcus | 1.4% | 0.6% | 1.0% | 0.049 1/ns |
| Catenibacterium | 0.5% | 0.7% | 4.3% | 0.037 1/ns |
References
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Foo, L. Identification and quantification of major polyphenols in apple pomace. Food Chem. 1997, 59, 187–194. [Google Scholar] [CrossRef]
- Grigoras, C.G.; Destandau, E.; Fougère, L.; Elfakir, C. Evaluation of apple pomace extracts as a source of bioactive compounds. Ind. Crop. Prod. 2013, 49, 794–804. [Google Scholar] [CrossRef]
- Gullón, B.; Falqué, E.; Alonso, J.L.; Parajó, J.C. Evaluation of apple pomace as a raw material for alternative applications in food industries. Food Technol. Biotechnol. 2007, 45, 426–433. [Google Scholar]
- Kołodziejczyk, K.; Markowski, J.; Kosmala, M.; Król, B.; Płocharski, W. Apple pomace as a potential source of nutraceutical products. Polish J. Food Nutr. Sci. 2007, 57, 291–295. [Google Scholar]
- Rana, S.; Gupta, S.; Rana, A.; Bhushan, S. Functional properties, phenolic constituents and antioxidant potential of industrial apple pomace for utilization as active food ingredient. Food Sci. Hum. Wellness 2015, 4, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Reis, S.F.; Rai, D.K.; Abu-Ghannam, N. Apple pomace as a potential ingredient for the development of new functional foods. Int. J. Food Sci. Technol. 2014, 49, 1743–1750. [Google Scholar] [CrossRef]
- Juśkiewicz, J.; Żary-Sikorska, E.; Zduńczyk, Z.; Król, B.; Jarosławska, J.; Jurgoński, A. Effect of dietary supplementation with unprocessed and ethanol-extracted apple pomaces on caecal fermentation, antioxidant and blood biomarkers in rats. Br. J. Nutr. 2011, 107, 1138–1146. [Google Scholar] [CrossRef] [Green Version]
- Sehm, J.; Lindermayer, H.; Dummer, C.; Treutter, D.; Pfaffl, M.W. The influence of polyphenol rich apple pomace or red-wine pomace diet on the gut morphology in weaning piglets. J. Anim. Physiol. Anim. Nutr. 2007, 91, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Sehm, J.; Treutter, D.; Lindermayer, H.; Meyer, H.H.D.; Pfaffl, M.W. The Influence of Apple- or Red-Grape Pomace Enriched Piglet Diet on Blood Parameters, Bacterial Colonisation, and Marker Gene Expression in Piglet White Blood Cells. Food Nutr. Sci. 2011, 2, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.F.; Vieira, R.G.; Zardo, D.M.; Falcão, L.D.; Nogueira, A.; Wosiacki, G. Apple pomace from eleven cultivars: An approach to identify sources of bioactive compounds. Acta Sci. Agron. 2010, 32, 29–35. [Google Scholar] [CrossRef]
- Andre, C.M.; Greenwood, J.M.; Walker, E.G.; Rassam, M.; Sullivan, M.; Evers, D.; Perry, N.B.; Laing, W.A. Anti-Inflammatory Procyanidins and Triterpenes in 109 Apple Varieties. J. Agric. Food Chem. 2012, 60, 10546–10554. [Google Scholar] [CrossRef]
- Thakur, B.R.; Singh, R.K.; Handa, A.K.; Rao, M.A. Chemistry and uses of pectin—A review. Crit. Rev. Food Sci. Nutr. 1997, 37, 47–73. [Google Scholar] [CrossRef]
- Heras-Ramírez, M.E.; Quintero-Ramos, A.; Camacho-Dávila, A.A.; Barnard, J.; Talamás-Abbud, R.; Torres-Muñoz, J.V.; Salas-Muñoz, E. Effect of Blanching and Drying Temperature on Polyphenolic Compound Stability and Antioxidant Capacity of Apple Pomace. Food Bioprocess Technol. 2011, 5, 2201–2210. [Google Scholar] [CrossRef]
- Pluske, J.R.; Le Dividich, J.; Verstegen, M.W.A. Weaning the Pig. Concepts and Consequences; Wageningen Academic Publishers: Wageningen, The Netherlands, 2003. [Google Scholar]
- Weary, D.M.; Jasper, J.; Hötzel, M.J. Understanding weaning distress. Appl. Anim. Behav. Sci. 2008, 110, 24–41. [Google Scholar] [CrossRef]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Niewold, T. Intestinal Health. Key to Maximise Growth Performance in Livestock; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015. [Google Scholar]
- Lallès, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Weaning—A challenge to gut physiologists. Livest. Sci. 2007, 108, 82–93. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Van Beers-Schreurs, H.; Vellenga, L.; Wensing, T.; Breukink, H. The pathogenesis of the post-weaning syndrome in weaned piglets; a review. Vet. Q. 1992, 14, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef]
- Aguedo, M.; Vanderghem, C.; Goffin, D.; Richel, A.; Paquot, M. Fast and high yield recovery of arabinose from destarched wheat bran. Ind. Crop. Prod. 2013, 43, 318–325. [Google Scholar] [CrossRef]
- Uerlings, J.; Schroyen, M.; Bautil, A.; Courtin, C.; Richel, A.; Sureda, E.A.; Bruggeman, G.; Tanghe, S.; Willems, E.; Bindelle, J.; et al. In Vitro prebiotic potential of agricultural by-products on intestinal fermentation, gut barrier and inflammatory status of piglets. Br. J. Nutr. 2020, 123, 293–307. [Google Scholar] [CrossRef]
- Dedrie, M.; Jacquet, N.; Bombeck, P.; Hébert, J.; Richel, A. Oak barks as raw materials for the extraction of polyphenols for the chemical and pharmaceutical sectors: A regional case study. Ind. Crop. Prod. 2015, 70, 316–321. [Google Scholar] [CrossRef]
- Oliviero, C.; Kokkonen, T.; Heinonen, M.; Sankari, S.; Peltoniemi, O. Feeding sows with high fibre diet around farrowing and early lactation: Impact on intestinal activity, energy balance related parameters and litter performance. Res. Vet. Sci. 2009, 86, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Laitat, M.; Antoine, N.; Cabaraux, J.; Cassart, D.; Mainil, J.; Moula, N.; Nicks, B.; Wavreille, J.; Philippe, F.-X. Influence of sugar beet pulp on feeding behaviour, growth performance, carcass quality and gut health of fattening pigs. Biotechnol. Agron. Soc. Environ. 2015, 19, 20–31. [Google Scholar]
- Douny, C.; Dufourny, S.; Brose, F.; Verachtert, P.; Rondia, P.; Lebrun, S.; Marzorati, M.; Everaert, N.; Delcenserie, V.; Scippo, M.-L. Development of an analytical method to detect short-chain fatty acids by SPME-GC–MS in samples coming from an In Vitro gastrointestinal model. J. Chromatogr. B 2019, 1124, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Dagnelie, P. Principes D’expérimentation. Planification des Expériences et Analyse de Leurs Résultats, 2nd ed.; Presses Agronomiques: Gembloux, Belgium, 2012; 413p. [Google Scholar]
- Montagne, L.; Pluske, J.; Hampson, D. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Jang, C.; Hui, S.; Lu, W.; Cowan, A.J.; Morscher, R.J.; Lee, G.; Liu, W.; Tesz, G.J.; Birnbaum, M.J.; Rabinowitz, J.D. The Small Intestine Converts Dietary Fructose into Glucose and Organic Acids. Cell Metab. 2018, 27, 351–361.e3. [Google Scholar] [CrossRef] [Green Version]
- Delarras, C. Pratique en Microbiologie de Laboratoire: Recherche de Bactéries et de Levures-Moisissures; Tec&Doc: Paris, France, 2014; ISBN 978-2-7430-1565-7. [Google Scholar]
- Brambillasca, S.; Zunino, P.; Cajarville, C. Addition of inulin, alfalfa and citrus pulp in diets for piglets: Influence on nutritional and faecal parameters, intestinal organs, and colonic fermentation and bacterial populations. Livestig. Sci. 2015, 178, 243–250. [Google Scholar] [CrossRef]
- McRorie, J.W.; McKeown, N.M. Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, M.G.D.; Knight, R.; Gilbert, J.A.; Blaser, M.J. Preserving microbial diversity. Science 2018, 362, 33–34. [Google Scholar] [CrossRef]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenterol. 2012, 6, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.; Hehemann, J.-H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and Gut Bacteroidetes: The Food Connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios-Covian, D.; Salazar, N.; Gueimonde, M.; Reyes-Gavilan, C.G.D.L. Shaping the Metabolism of Intestinal Bacteroides Population through Diet to Improve Human Health. Front. Microbiol. 2017, 8, 376. [Google Scholar] [CrossRef] [Green Version]
- Wexler, H.M.; Daya, S.; Berns, K.I. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E. Prevotella in the gut: Choose carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Spees, A.M.; Lopez, C.A.; Kingsbury, D.D.; Winter, S.E.; Bäumler, A.J. Colonization Resistance: Battle of the Bugs or Ménage à Trois with the Host? PLoS Pathog. 2013, 9, e1003730. [Google Scholar] [CrossRef] [Green Version]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroyen, M.; Liu, H.; Tuggle, C.K. Applications of systems biology to improve pig health. In Systems Biology in Animal Production and Health; Kadarmideen, H.N., Ed.; Springer: Cham, Switzerland, 2016; pp. 33–59. [Google Scholar]
- Mon, K.K.Z.; Saelao, P.; Halstead, M.M.; Chanthavixay, G.; Chang, H.-C.; Garas, L.; Maga, E.A.; Zhou, H. Salmonella enterica Serovars Enteritidis Infection Alters the Indigenous Microbiota Diversity in Young Layer Chicks. Front. Vet. Sci. 2015, 2, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popov, S.V.; Ovodov, Y.S. Polypotency of the immunomodulatory effect of pectins. Biochemistry (Moscow) 2013, 78, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; De Souza, C.B.; Krych, L.; Cahú, T.B.; Wiese, M.; Kot, W.; Hansen, K.M.; Blennow, A.; Venema, K.; Jespersen, L. Potential of Pectins to Beneficially Modulate the Gut Microbiota Depends on Their Structural Properties. Front. Microbiol. 2019, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Bruggeman, G.; Berg, M.V.D.; Borewicz, K.; Scheurink, A.J.W.; Bruininx, E.; De Vos, P.; Smidt, H.; Schols, H.A.; Gruppen, H. Effects of pectin on fermentation characteristics, carbohydrate utilization, and microbial community composition in the gastrointestinal tract of weaning pigs. Mol. Nutr. Food Res. 2017, 61, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bland, E.J.; Keshavarz, T.; Bucke, C. The influence of small oligosaccharides on the immune system. Carbohydr. Res. 2004, 339, 1673–1678. [Google Scholar] [CrossRef]
- Olano-Martin, E.; Gibson, G.; Rastall, R. Comparison of the in vitro bifidogenic properties of pectins and pectic-oligosaccharides. J. Appl. Microbiol. 2002, 93, 505–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Tamimi, M.; Palframan, R.; Cooper, J.; Gibson, G.; Rastall, R. In vitro fermentation of sugar beet arabinan and arabino-oligosaccharides by the human gut microflora. J. Appl. Microbiol. 2005, 100, 407–414. [Google Scholar] [CrossRef]
- Maki, K.C.; Gibson, G.R.; Dickmann, R.S.; Kendall, C.W.; Chen, C.-Y.O.; Costabile, A.; Comelli, E.M.; McKay, D.L.; Almeida, N.G.; Jenkins, D.; et al. Digestive and physiologic effects of a wheat bran extract, arabino-xylan-oligosaccharide, in breakfast cereal. Nutrition 2012, 28, 1115–1121. [Google Scholar] [CrossRef]
- Li, W.; Yang, R.; Ying, D.; Yu, J.; Sanguansri, L.; Augustin, M.A. Analysis of polyphenols in apple pomace: A comparative study of different extraction and hydrolysis procedures. Ind. Crop. Prod. 2020, 147, 112250. [Google Scholar] [CrossRef]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Aprikian, O.; Duclos, V.; Guyot, S.; Besson, C.; Manach, C.; Bernalier, A.; Morand, C.; Rémésy, C.; Demigné, C. Apple Pectin and a Polyphenol-Rich Apple Concentrate Are More Effective Together Than Separately on Cecal Fermentations and Plasma Lipids in Rats. J. Nutr. 2003, 133, 1860–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, K.E.B.; Hedemann, M.S.; Lærke, H.N. The role of carbohydrates in intestinal health of pigs. Anim. Feed Sci. Technol. 2012, 173, 41–53. [Google Scholar] [CrossRef]


| Ingredients | 0% AP | 2% AP | 4% AP |
|---|---|---|---|
| Wheat | 36.89 | 34.16 | 31.45 |
| Barley | 20.00 | 20.00 | 20.00 |
| Soybean meal | 16.40 | 16.83 | 17.28 |
| Maize | 15.00 | 15.00 | 15.00 |
| Premix10916 (Inve Belgie, Dendermonde, Belgium) | 7.50 | 7.50 | 7.50 |
| Sugar beet pulp | 2.50 | 2.50 | 2.50 |
| AP | 0.00 | 2.00 | 4.00 |
| Soya oil | 1.45 | 1.72 | 1.97 |
| L-threonine | 0.115 | 0.120 | 0.120 |
| Monocalcium phosphate | 0.100 | 0.100 | 0.100 |
| Tryptophan | 0.035 | 0.035 | 0.040 |
| DL-Methionine | 0.015 | 0.025 | 0.030 |
| Rovimix® E50 | 0.005 | 0.005 | 0.005 |
| Chemical Component | AP |
|---|---|
| Rhamnose | <0.1 g/100 g DM |
| Arabinose | <0.1 g/100 g DM |
| Xylose | <0.1 g/100 g DM |
| Mannose | <0.1 g/100 g DM |
| Glucose | 3.4 g/100 g DM |
| Galactose | 2.1 g/100 g DM |
| Fructose | 13.5 g/100 g DM |
| Saccharose | 2.9 g/100 g DM |
| Arabino-oligosaccharide | 0.5 g/100 g DM |
| Xylo-oligosaccharide | 0.3 g/100 g DM |
| Galacto-oligosaccharide | 0.2 g/100 g DM |
| Galacturonic acid | <0.1 g/100 g DM |
| Glucuronic acid | <0.1 g/100 g DM |
| Total polyphenols (Folin-Ciocalteu) | 26.7 mg gallic acid/g DM |
| Quercetin | 23.1 µg/g DM |
| Phloretin | 2.8 µg/g DM |
| Insoluble HMWDF | 45.2 g/100 g DM |
| Soluble HMWDF | 12.6 g/100 g DM |
| Soluble LMWDF | 0.5 g/100 g DM |
| Soluble DF/insoluble DF | 0.29 |
| Chemical Component | 0% AP | 2% AP | 4% AP |
|---|---|---|---|
| Dry Matter (%) | 88.0 | 87.3 | 88.2 |
| Crude protein (% DM) | 18.1 | 18.7 | 18.6 |
| Crude ash (% DM) | 6.13 | 5.51 | 5.45 |
| Reducing sugars (% DM) | 1.43 | 1.76 | 2.47 |
| Total sugars (% DM) | 6.40 | 7.17 | 7.93 |
| Starch (% DM) | 44.10 | 43.41 | 42.23 |
| NDF (% DM) | 13.49 | 13.20 | 13.20 |
| ADF (% DM) | 6.49 | 6.56 | 6.68 |
| ADL (% DM) | 1.21 | 1.15 | 1.38 |
| Cellulose (% DM) | 5.13 | 5.12 | 4.93 |
| Crude Fat (% DM) | 3.34 | 3.55 | 3.50 |
| Insoluble HMWDF (% DM) | 17.3 | 17.4 | 16.7 |
| Soluble HMWDF (% DM) | 4.7 | 4.9 | 5.1 |
| Soluble LMWDF (% DM) | 3.3 | 3.0 | 3.1 |
| Soluble DF/insoluble DF | 0.46 | 0.45 | 0.49 |
| Zootechnical Parameter | 0% AP | 2% AP | 4% AP | SEM | p-Value Diet | p-Value Block |
|---|---|---|---|---|---|---|
| Initial weight (kg) | 8.4 | 8.4 | 8.4 | 0.1 | ns | ns |
| Final weight (kg) | 21.0 | 22.6 | 23.1 | 0.4 | ns | ns |
| Final ADG (kg/d) | 0.361 | 0.406 | 0.421 | 0.012 | ns | ns |
| TFI (kg DM) | 19.8 | 21.7 | 20.5 | 0.5 | ns | ns |
| FCR | 1.79 a | 1.75 a | 1.59 b | 0.03 | 0.002 | ns |
| EFE (MJ NE/kg gain) | 16.9 a | 16.8 a | 15.2 b | 0.2 | 0.004 | ns |
| ADG | 0% AP | 2% AP | 4% AP | SEM | p-Value Diet | p-Value Block |
|---|---|---|---|---|---|---|
| ADG week 1 (d0–d + 7) | 0.118 | 0.134 | 0.143 | 0.013 | ns | ns |
| ADG week 2 (d + 7–d + 14) | 0.304 | 0.261 | 0.350 | 0.020 | 0.083 | ns |
| ADG week 3 (d + 14–d + 21) | 0.367 b | 0.459 a | 0.458 a | 0.017 | 0.038 | ns |
| ADG week 4 (d + 21–d + 28) | 0.489 | 0.563 | 0.537 | 0.018 | 0.085 | ns |
| ADG week 5 (d + 28–d + 35) | 0.530 | 0.613 | 0.614 | 0.020 | ns | ns |
| Diet | Statistical Parameter | Fecal Consistency Scoring 1 | Pathogens in Feces 2 | ||||
|---|---|---|---|---|---|---|---|
| A | B | Total | Negative | Positive | Total | ||
| Apple pomace | Count | 51 | 144 | 195 | 84 | 20 | 104 |
| Expected count | 57.47 | 137.53 | 87.58 | 16.42 | |||
| Adjusted Residuals | −1.809 | 1.809 | −1.713 | 1.713 | |||
| No apple pomace | Count | 33 | 57 | 90 | 44 | 4 | 48 |
| Expected count | 26.53 | 63.47 | 40.42 | 7.58 | |||
| Adjusted Residuals | 1.809 | −1.809 | 1.713 | −1.713 | |||
| Total | Count | 84 | 201 | 285 | 128 | 24 | 152 |
| p-value | 0.096 | 0.072 | |||||
| Intestinal Parameter | 0% AP | 2% AP | 4% AP | SEM | p-Value Diet | p-Value Block |
|---|---|---|---|---|---|---|
| Duodenum | (n = 6) | (n = 7) | (n = 6) | |||
| Villus length (µm) | 320 b | 381 ab | 429 a | 16.4 | 0.018 | ns |
| Crypts depth (µm) | 436 | 471 | 448 | 10.5 | ns | ns |
| VL/CD | 0.74 | 0.81 | 0.96 | 0.039 | ns | ns |
| Jejunum | (n = 6) | (n = 6) | (n = 5) | |||
| Villus length (µm) | 354 | 366 | 420 | 20.5 | ns | ns |
| Crypts depth (µm) | 314 | 325 | 324 | 8.9 | ns | ns |
| VL/CD | 1.14 | 1.16 | 1.31 | 0.07 | ns | ns |
| Ileum | (n = 6) | (n = 7) | (n = 6) | |||
| Villus length (µm) | 240 | 273 | 315 | 13.4 | ns | ns |
| Crypts depth (µm) | 255 | 270 | 242 | 7.9 | ns | ns |
| VL/CD | 0.94 b | 1.03 ab | 1.33 a | 0.07 | 0.039 | ns |
| Phylum-Class Family—Genus (If app.)—Species (If app.) | 0%AP | 2%AP | 4%AP | SEM | p-Value KW/FDR |
|---|---|---|---|---|---|
| Actinobacteria–Actinobacteria- | |||||
| Bifidobacteriaceae | 0.0 | 0.3 | 0.0 | 0.06 | ns/ns |
| Actinobacteria–Coriobacteriia- | |||||
| Coriobacteriaceae | 1.9 | 1.9 | 1.8 | 0.21 | ns/ns |
| Bacteroidetes–Bacteroidia- | |||||
| Bacteroidales (undef. fam.) | 0.1 | 0.2 | 0.3 | 0.06 | ns/ns |
| Prevotellaceae | 2.2 | 3.2 | 1.4 | 0.31 | ns/ns |
| Muribaculaceae (formerly called S24-7) | 4.0 | 2.5 | 8.7 | 1.46 | ns/ns |
| (Paraprevotellaceae) | 0.1 | 0.1 | 0.0 | 0.01 | ns/ns |
| Cyanobacteria–4C0d-2 | |||||
| YS2 (undef. fam.) | 0.1 | 0.2 | 0.2 | 0.04 | ns/ns |
| Firmicutes–Bacilli- | |||||
| Enterococcaceae | 0.1 | 0.0 | 0.0 | 0.03 | ns/ns |
| Lactobacillaceae | 25.3 | 31.3 | 23.2 | 2.91 | ns/ns |
| Streptococcaceae | 6.1 | 3.0 | 1.8 | 0.84 | ns/ns |
| Firmicutes–Clostridia- | |||||
| Clostridiales (Other fam.) | 0.1 | 0.1 | 0.2 | 0.03 | ns/ns |
| Clostridiales (undef. fam.) | 3.7 | 2.7 | 5.5 | 0.44 | 0.099/ns |
| Christensenellaceae | 1.5 | 0.9 | 3.1 | 0.61 | ns/ns |
| Clostridiaceae | 3.5 ab | 2.3 b | 5.9 a | 0.64 | 0.028/ns |
| Clostridiaceae -SMB53 (undef. sp) | 0.6 ab | 0.4 b | 1.1 a | 0.14 | 0.036/ns |
| Clostridiaceae (undefined genus) | 2.8 ab | 1.6 b | 4.6 a | 0.54 | 0.019/ns |
| Dehalobacteriaceae | 0.1 | 0.0 | 0.1 | 0.02 | ns/ns |
| Eubacteriaceae | 0.0 | 0.2 | 0.0 | 0.05 | ns/ns |
| Lachnospiraceae | 16.3 | 19.8 | 14.1 | 1.50 | ns/ns |
| Lachnospiraceae -Blautia (other sp.) | 0.1 a | 0.0 b | 0.0 b | 0.02 | 0.024/ns |
| Lachnospiraceae -Dorea (other sp.) | 0.8 a | 0.6 a | 0.2 b | 0.12 | 0.024/ns |
| Lachnospiraceae -Lachnospira (undef. sp) | 0.4 ab | 0.4 a | 0.1 b | 0.06 | 0.036/ns |
| Peptococcaceae | 0.2 | 0.1 | 0.2 | 0.02 | ns/ns |
| Peptostreptococcaceae | 0.1 | 0.1 | 0.1 | 0.02 | ns/ns |
| Ruminococcaceae | 13.6 ab | 12.9 b | 19.8 a | 1.27 | 0.047/ns |
| Veillonellaceae | 12.2 a | 11.4 a | 4.8 b | 1.12 | 0.009/ns |
| Veillonellaceae -Dialister (undef. sp) | 2.3 a | 2.1 ab | 0.5 b | 0.32 | 0.016/ns |
| Veillonellaceae -Megasphaera (undef. sp) | 5.7 a | 6.6 a | 2.8 b | 0.59 | 0.015/ns |
| Veillonellaceae -Mitsuokella (undef. sp) | 2.4 a | 1.1 b | 0.6 b | 0.29 | 0.023/ns |
| Veillonellaceae -Mitsuokella multacida | 0.1 | 0.5 | 0.1 | 0.09 | 0.063/ns |
| Veillonellaceae -(undef. genus) | 0.5 | 0.3 | 0.1 | 0.08 | 0.064/ns |
| [Mogibacteriaceae] | 1.4 | 1.2 | 1.6 | 0.16 | ns/ns |
| Firmicutes–Erysipelotrichi- | |||||
| Erysipelotrichaceae | 6.6 | 5.0 | 5.3 | 0.62 | ns/ns |
| Erysipelotrichaceae -[Eubacterium] cylindroides | 0.7 | 0.3 | 0.6 | 0.09 | 0.092/ns |
| Erysipelotrichaceae -Bulleidia p-1630-c5 | 1.3 | 0.9 | 0.6 | 0.01 | 0.085/ns |
| Erysipelotrichaceae -L7A_E11 (undef. sp) | 0.0 | 0.0 | 0.1 | 0.03 | 0.060/ns |
| Erysipelotrichaceae -(undef. genus) | 0.1 | 0.1 | 0.2 | 0.02 | 0.085/ns |
| Planctomycetes–Planctomycetia- | |||||
| Pirellulaceae | 0.2 | 0.2 | 0.2 | 0.08 | ns/ns |
| Proteobacteria–Deltaproteobacteria- | |||||
| Desulfovibrionaceae | 0.1 | 0.0 | 0.1 | 0.02 | 0.071/ns |
| -Desulfovibrio (undef. sp) | 0.1 | 0.0 | 0.1 | 0.100/ns | |
| Proteobacteria–Gammaproteobacteria- | |||||
| Succinivibrionaceae | 0.0 | 0.1 | 0.0 | 0.01 | ns/ns |
| TM7–TM7-3- | |||||
| F16 | 0.0 | 0.1 | 0.1 | 0.03 | ns/ns |
| Tenericutes–Mollicutes- | |||||
| RF39 (undef. fam.) | 0.1 | 0.1 | 0.1 | 0.01 | ns/ns |
| WPS-2—(undefined class)- | |||||
| (undef. fam.) | 0.0 | 0.0 | 0.6 | 0.18 | 0.065/ns |
| Index | 0% AP | 2% AP | 4% AP | p-Value KW |
|---|---|---|---|---|
| Chyme | ||||
| Chao 1 | 1103 ab | 990 b | 1189 a | 0.015 |
| OTU | 707 ab | 633 b | 790 a | 0.024 |
| PD Whole Tree | 40.0 ab | 36.1 b | 44.3 a | 0.030 |
| Shannon | 5.4 | 5.3 | 5.7 | ns |
| Mucosa | ||||
| Chao 1 | 771 ab | 668 b | 849 a | 0.045 |
| OTU | 491 | 423 | 527 | 0.084 |
| PD Whole Tree | 34.1 | 30.2 | 36.4 | ns |
| Shannon | 6.0 | 5.5 | 6.1 | ns |
| Phylum-Class: Family–Genus–Species | 0% AP | 2% AP | 4% AP | SEM | p-Value KW/FDR |
|---|---|---|---|---|---|
| Bacteroidetes | 14.6 a | 6.5 b | 13.6 a | 1.27 | 0.049/ns |
| Bacteroidia: | 14.6 a | 6.5 b | 13.6 a | 1.27 | 0.049/ns |
| Prevotellaceae | 11.9 | 5.2 | 10.5 | 1.08 | 0.060/ns |
| Prevotellaceae–Prevotella (undef. sp.) | 3.0 a | 1.0 b | 3.6 a | 0.36 | 0.010/ns |
| Muribaculaceae (formerly called s24-7) | 0.2 ab | 0.1 b | 0.3 a | 0.03 | 0.037/ns |
| Firmicutes | 78.3 b | 90.1 a | 82.4 b | 1.68 | 0.045/ns |
| Clostridia: | 46.5 | 49.4 | 54.5 | 2.02 | ns/ns |
| Lachnospiraceae–Coprococcus (undef. sp.) | 0.7 | 0.5 | 0.9 | 0.07 | 0.073/ns |
| Peptostreptococcaceae | 0.1 | 0.1 | 0.1 | 0.01 | 0.074/ns |
| Ruminococcaceae–Oscillospira (undef. sp.) | 0.2 a | 0.1 b | 0.3 a | 0.03 | 0.037/ns |
| Veillonellaceae–Acidaminococcus (undef. sp.) | 0.1 | 0.2 | 0.3 | 0.04 | 0.088/ns |
| Veillonellaceae–Megamonas (undef. sp.) | 0.4 | 3.5 | 0.1 | 0.97 | 0.070/ns |
| Veillonellaceae–Mitsuokella multacida | 0.0 | 0.5 | 0.1 | 0.10 | 0.094/ns |
| Proteobacteria | 5.6 | 1.3 | 3.0 | 0.63 | 0.053/ns |
| Epsilonproteobacteria: | 3.2 | 0.9 | 1.4 | 0.39 | ns/ns |
| Campylobacteraceae–Campylobacter (undef. sp.) | 2.5 | 0.5 | 0.9 | 0.34 | 0.083/ns |
| Gammaproteobacteria: | 2.1 | 0.3 | 1.4 | 0.33 | ns/ns |
| Pasteurellaceae | 0.2 | 0.0 | 0.0 | 0.03 | 0.057/ns |
| Pasteurellaceae–Actinobacillus (Other sp.) | 0.1 a | 0.0 b | 0.0 a | 0.02 | 0.030/ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufourny, S.; Antoine, N.; Pitchugina, E.; Delcenserie, V.; Godbout, S.; Douny, C.; Scippo, M.-L.; Froidmont, E.; Rondia, P.; Wavreille, J.; et al. Apple Pomace and Performance, Intestinal Morphology and Microbiota of Weaned Piglets—A Weaning Strategy for Gut Health? Microorganisms 2021, 9, 572. https://doi.org/10.3390/microorganisms9030572
Dufourny S, Antoine N, Pitchugina E, Delcenserie V, Godbout S, Douny C, Scippo M-L, Froidmont E, Rondia P, Wavreille J, et al. Apple Pomace and Performance, Intestinal Morphology and Microbiota of Weaned Piglets—A Weaning Strategy for Gut Health? Microorganisms. 2021; 9(3):572. https://doi.org/10.3390/microorganisms9030572
Chicago/Turabian StyleDufourny, Sandrine, Nadine Antoine, Elena Pitchugina, Véronique Delcenserie, Stéphane Godbout, Caroline Douny, Marie-Louise Scippo, Eric Froidmont, Pierre Rondia, José Wavreille, and et al. 2021. "Apple Pomace and Performance, Intestinal Morphology and Microbiota of Weaned Piglets—A Weaning Strategy for Gut Health?" Microorganisms 9, no. 3: 572. https://doi.org/10.3390/microorganisms9030572