
Inoculation of the Leishmania infantum HSP70-II Null Mutant Induces Long-Term Protection against L. amazonensis Infection in BALB/c Mice

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice, Parasites, Vaccination, and Challenge
2.2. Vaccination, Challenge, and Disease Follow-Up
2.3. Sera Preparation and ELISA
2.4. In Vitro Spleen Cell Stimulation and Analysis of Cytokine Concentration in Culture Supernatants
2.5. In Vitro DNLs Cell Stimulation and Analysis of Cytokine Concentration in Culture Supernatants
2.6. Analysis of T Cell Populations by Flow Cytometry
2.7. Statistical Analysis
3. Results
3.1. The LiΔHSP70-II Based Vaccine Induces Long-Term Protection Against a L. amazonensis Virulent Challenge in BALB/c Mice
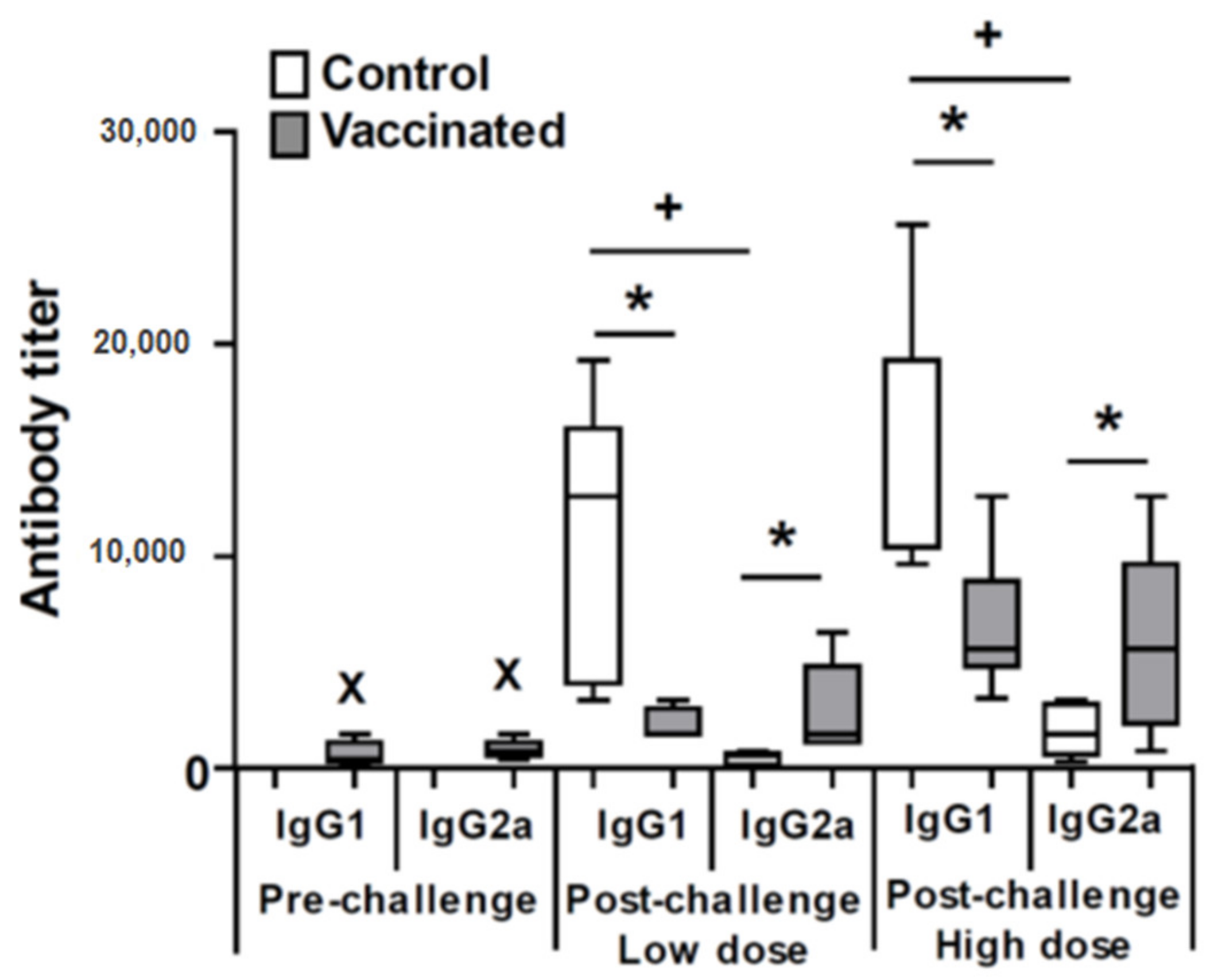
3.2. Vaccination with the LiΔHSP70-II Attenuated Cell Line Modulates the Profile of the Anti-Leishmanial Antibodies and Parasite-Specific Systemic Cytokine Secretion after Challenge with L. amazonensis
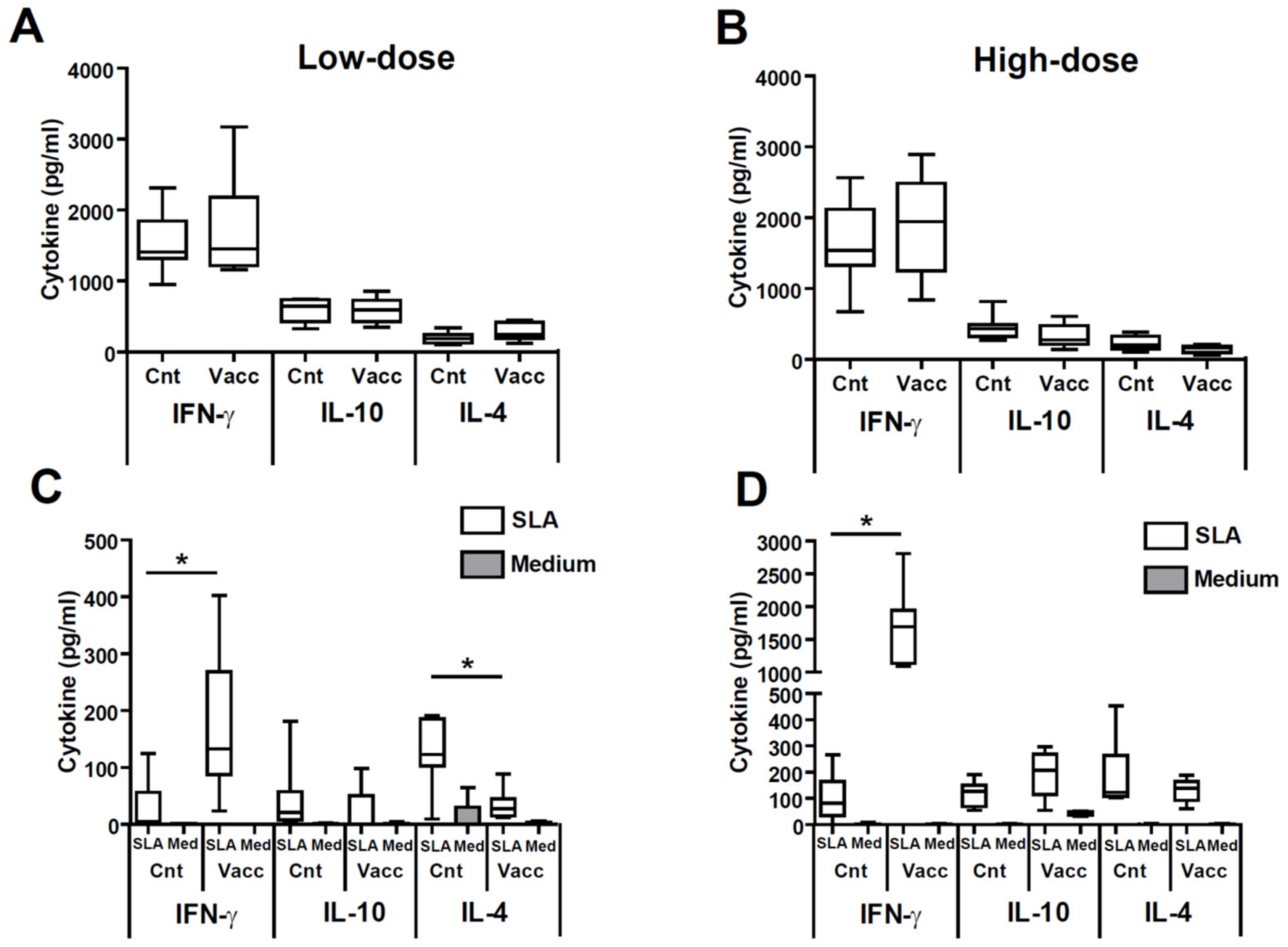
3.3. Vaccinated Animals Show a Local Cytokine Response Directed towards the Production of IFN-γ against the Parasite
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silveira, F.T. What makes mucosal and anergic diffuse cutaneous leishmaniases so clinically and immunopathogically different? A review in Brazil. Trans. R. Soc. Trop. Med. Hyg. 2019. [Google Scholar] [CrossRef]
- Silveira, F.T.; Blackwell, J.M.; Ishikawa, E.A.; Braga, R.; Shaw, J.J.; Quinnell, R.J.; Soong, L.; Kima, P.; McMahon-Pratt, D.; Black, G.F.; et al. T cell responses to crude and defined leishmanial antigens in patients from the lower Amazon region of Brazil infected with different species of Leishmania of the subgenera Leishmania and Viannia. Parasite Iimmunol. 1998, 20, 19–26. [Google Scholar] [CrossRef]
- Silveira, F.T.; Lainson, R.; Corbett, C.E. Clinical and immunopathological spectrum of American cutaneous leishmaniasis with special reference to the disease in Amazonian Brazil: A review. Mem. Inst. Oswaldo Cruz 2004, 99, 239–251. [Google Scholar] [CrossRef]
- Alexander, J.; Brombacher, F. T helper1/t helper2 cells and resistance/susceptibility to Leishmania infection: Is this paradigm still relevant? Front. Immunol. 2012, 3, 80. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.; Novais, F.O. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat. Rev. Immunol. 2016, 16, 581–592. [Google Scholar] [CrossRef]
- Pereira, B.A.; Alves, C.R. Immunological characteristics of experimental murine infection with Leishmania (Leishmania) amazonensis. Vet. Parasitol. 2008, 158, 239–255. [Google Scholar] [CrossRef]
- De Lara da Silva, C.E.; Oyama, J.; Ferreira, F.B.P.; de Paula Lalucci-Silva, M.P.; Lordani, T.V.A.; de Lara da Silva, R.C.; de Souza Terron Monich, M.; Teixeira, J.J.V.; Lonardoni, M.V.C. Effect of essential oils on Leishmania amazonensis: A systematic review. Parasitology 2020, 147, 1392–1407. [Google Scholar] [CrossRef]
- Sanchez, M.V.; Elicabe, R.J.; Di Genaro, M.S.; Germano, M.J.; Gea, S.; Garcia Bustos, M.F.; Salomon, M.C.; Scodeller, E.A.; Cargnelutti, D.E. Total Leishmania antigens with Poly(I:C) induce Th1 protective response. Parasite Immunol. 2017, 39. [Google Scholar] [CrossRef]
- Cargnelutti, D.E.; Salomon, M.C.; Celedon, V.; Garcia Bustos, M.F.; Morea, G.; Cuello-Carrion, F.D.; Scodeller, E.A. Immunization with antigenic extracts of Leishmania associated with Montanide ISA 763 adjuvant induces partial protection in BALB/c mice against Leishmania (Leishmania) amazonensis infection. J. Microbiol. Immunol. Infect. 2016, 49, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, M.X.; Barcante, T.A.; Vilela, L.; Tafuri, W.L.; Afonso, L.C.; Vieira, L.Q. Vaccine-induced protection against Leishmania amazonensis is obtained in the absence of IL-12/23p40. Immunol. Lett. 2006, 105, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.F.; Pinheiro, R.O.; Rayol, A.; Larraga, V.; Rossi-Bergmann, B. Intranasal vaccination against cutaneous leishmaniasis with a particulated leishmanial antigen or DNA encoding LACK. Infect. Immun. 2004, 72, 4521–4527. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.F.; de Mello Cortezia, M.; Rossi-Bergmann, B. Interferon-gamma-inducing oral vaccination with Leishmania amazonensis antigens protects BALB/c and C57BL/6 mice against cutaneous leishmaniasis. Vaccine 2003, 21, 3534–3541. [Google Scholar] [CrossRef]
- De Matos Guedes, H.L.; da Silva Costa, B.L.; Chaves, S.P.; de Oliveira Gomes, D.C.; Nosanchuk, J.D.; De Simone, S.G.; Rossi-Bergmann, B. Intranasal vaccination with extracellular serine proteases of Leishmania amazonensis confers protective immunity to BALB/c mice against infection. Parasites Vectors 2014, 7, 448. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.; Diao, H.; Ji, J.; Soong, L. DNA immunization with the gene encoding P4 nuclease of Leishmania amazonensis protects mice against cutaneous leishmaniasis. Infect. Immun. 2003, 71, 6270–6278. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.; Popov, V.; Soong, L. Identification and molecular characterization of a gene encoding a protective Leishmania amazonensis Trp-Asp (WD) protein. Infect. Immun. 2004, 72, 2194–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez-Fumagalli, M.A.; Costa, M.A.; Oliveira, D.M.; Ramirez, L.; Costa, L.E.; Duarte, M.C.; Martins, V.T.; Oliveira, J.S.; Olortegi, C.C.; Bonay, P.; et al. Vaccination with the Leishmania infantum ribosomal proteins induces protection in BALB/c mice against Leishmania chagasi and Leishmania amazonensis challenge. Microbes. Infect. 2010, 12, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.C.; Lage, D.P.; Martins, V.T.; Costa, L.E.; Carvalho, A.; Ludolf, F.; Santos, T.T.O.; Vale, D.L.; Roatt, B.M.; Menezes-Souza, D.; et al. A vaccine composed of a hypothetical protein and the eukaryotic initiation factor 5a from Leishmania braziliensis cross-protection against Leishmania amazonensis infection. Immunobiology 2017, 222, 251–260. [Google Scholar] [CrossRef]
- Martinez-Rodrigo, A.; Dias, D.S.; Ribeiro, P.A.F.; Roatt, B.M.; Mas, A.; Carrion, J.; Coelho, E.A.F.; Dominguez-Bernal, G. Immunization with the HisAK70 DNA Vaccine induces resistance against Leishmania amazonensis infection in BALB/c mice. Vaccines 2019, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- Saljoughian, N.; Taheri, T.; Rafati, S. Live vaccination tactics: Possible approaches for controlling visceral leishmaniasis. Front. Immunol. 2014, 5, 134. [Google Scholar] [CrossRef] [Green Version]
- Zabala-Penafiel, A.; Todd, D.; Daneshvar, H.; Burchmore, R. The potential of live attenuated vaccines against Cutaneous Leishmaniasis. Exp. Parasitol. 2020, 210, 107849. [Google Scholar] [CrossRef]
- Viana, S.M.; Celes, F.S.; Ramirez, L.; Kolli, B.; Ng, D.K.P.; Chang, K.P.; de Oliveira, C.I. Photodynamic Vaccination of BALB/c Mice for Prophylaxis of Cutaneous Leishmaniasis Caused by Leishmania amazonensis. Front. Microbiol. 2018, 9, 165. [Google Scholar] [CrossRef]
- Zorgi, N.E.; Arruda, L.V.; Paladine, I.; Roque, G.A.S.; Araujo, T.F.; Brocchi, M.; Barral, M.; Sanchiz, A.; Requena, J.M.; Abanades, D.R.; et al. Leishmania infantum transfected with toxic plasmid induces protection in mice infected with wild type L. infantum or L. amazonensis. Mol. Immunol. 2020, 127, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Veras, P.; Brodskyn, C.; Balestieri, F.; Freitas, L.; Ramos, A.; Queiroz, A.; Barral, A.; Beverley, S.; Barral-Netto, M. A dhfr-ts- Leishmania major knockout mutant cross-protects against Leishmania amazonensis. Mem. Inst. Oswaldo Cruz 1999, 94, 491–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrion, J.; Folgueira, C.; Soto, M.; Fresno, M.; Requena, J.M. Leishmania infantum HSP70-II null mutant as candidate vaccine against leishmaniasis: A preliminary evaluation. Parasites Vectors 2011, 4, 150. [Google Scholar] [CrossRef] [Green Version]
- Solana, J.C.; Ramirez, L.; Corvo, L.; de Oliveira, C.I.; Barral-Netto, M.; Requena, J.M.; Iborra, S.; Soto, M. Vaccination with a Leishmania infantum HSP70-II null mutant confers long-term protective immunity against Leishmania major infection in two mice models. PLoS Negl. Trop. Dis. 2017, 11, e0005644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solana, J.C.; Ramirez, L.; Cook, E.C.; Hernandez-Garcia, E.; Sacristan, S.; Martin, M.E.; Manuel Gonzalez, V.; Reguera, R.M.; Balana-Fouce, R.; Fresno, M.; et al. Subcutaneous immunization of Leishmania HSP70-II null mutant line reduces the severity of the experimental visceral leishmaniasis in BALB/c mice. Vaccines 2020, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Folgueira, C.; Quijada, L.; Soto, M.; Abanades, D.R.; Alonso, C.; Requena, J.M. The Translational Efficiencies of the two Leishmania infantum HSP70 mRNAs, differing in their 3′-Untranslated regions, are affected by shifts in the temperature of growth through different mechanisms. J. Biol. Chem. 2005, 280, 35172–35183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buffet, P.A.; Sulahian, A.; Garin, Y.J.; Nassar, N.; Derouin, F. Culture microtitration: A sensitive method for quantifying Leishmania infantum in tissues of infected mice. Antimicrob. Agents Chemother. 1995, 39, 2167–2168. [Google Scholar] [CrossRef] [Green Version]
- Coffman, R.L. Mechanisms of helper T-cell regulation of B-cell activity. Ann. N. Y. Acad. Sci. 1993, 681, 25–28. [Google Scholar] [CrossRef]
- Mohebali, M.; Nadim, A.; Khamesipour, A. An overview of leishmanization experience: A successful control measure and a tool to evaluate candidate vaccines. Acta Trop. 2019, 200, 105173. [Google Scholar] [CrossRef]
- Courret, N.; Lang, T.; Milon, G.; Antoine, J.C. Intradermal inoculations of low doses of Leishmania major and Leishmania amazonensis metacyclic promastigotes induce different immunoparasitic processes and status of protection in BALB/c mice. Int. J. Parasitol. 2003, 33, 1373–1383. [Google Scholar] [CrossRef]
- Ferrua, B.; Luci, C.; Le Fichoux, Y.; Paul, A.; Marty, P. Imprinting of BALB/c mice with low Leishmania infantum parasite dose markedly protects spleen against high-dose challenge. Vaccine 2006, 24, 589–596. [Google Scholar] [CrossRef]
- Cortes, D.F.; Carneiro, M.B.; Santos, L.M.; Souza, T.C.; Maioli, T.U.; Duz, A.L.; Ramos-Jorge, M.L.; Afonso, L.C.; Carneiro, C.; Vieira, L.Q. Low and high-dose intradermal infection with Leishmania major and Leishmania amazonensis in C57BL/6 mice. Mem. Inst. Oswaldo Cruz 2010, 105, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Porrozzi, R.; Santos da Costa, M.V.; Teva, A.; Falqueto, A.; Ferreira, A.L.; dos Santos, C.D.; Fernandes, A.P.; Gazzinelli, R.T.; Campos-Neto, A.; Grimaldi, G., Jr. Comparative evaluation of enzyme-linked immunosorbent assays based on crude and recombinant leishmanial antigens for serodiagnosis of symptomatic and asymptomatic Leishmania infantum visceral infections in dogs. Clin. Vaccine Immunol. 2007, 14, 544–548. [Google Scholar] [CrossRef] [Green Version]
- Jain, K.; Jain, N.K. Vaccines for visceral leishmaniasis: A review. J. Immunol. Methods 2015, 422, 1–12. [Google Scholar] [CrossRef]
- Mandell, M.A.; Beverley, S.M. Continual renewal and replication of persistent Leishmania major parasites in concomitantly immune hosts. Proc. Natl. Acad. Sci. USA 2017, 114, E801–E810. [Google Scholar] [CrossRef] [Green Version]
- Sacks, D.L. Vaccines against tropical parasitic diseases: A persisting answer to a persisting problem. Nat. Immunol. 2014, 15, 403–405. [Google Scholar] [CrossRef]
- Zhang, W.W.; Karmakar, S.; Gannavaram, S.; Dey, R.; Lypaczewski, P.; Ismail, N.; Siddiqui, A.; Simonyan, V.; Oliveira, F.; Coutinho-Abreu, I.V.; et al. A second generation leishmanization vaccine with a markerless attenuated Leishmania major strain using CRISPR gene editing. Nat. Commun. 2020, 11, 3461. [Google Scholar] [CrossRef]
- Fiuza, J.A.; Dey, R.; Davenport, D.; Abdeladhim, M.; Meneses, C.; Oliveira, F.; Kamhawi, S.; Valenzuela, J.G.; Gannavaram, S.; Nakhasi, H.L. Intradermal immunization of Leishmania donovani Centrin Knock-Out parasites in combination with salivary protein LJM19 from sand fly vector induces a durable protective immune response in hamsters. PLoS Negl. Trop. Dis. 2016, 10, e0004322. [Google Scholar] [CrossRef]
- Peters, N.C.; Pagan, A.J.; Lawyer, P.G.; Hand, T.W.; Henrique Roma, E.; Stamper, L.W.; Romano, A.; Sacks, D.L. Chronic parasitic infection maintains high frequencies of short-lived Ly6C+CD4+ effector T cells that are required for protection against re-infection. PLoS Pathog. 2014, 10, e1004538. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Cotrina, J.; Iniesta, V.; Belinchon-Lorenzo, S.; Munoz-Madrid, R.; Serrano, F.; Parejo, J.C.; Gomez-Gordo, L.; Soto, M.; Alonso, C.; Gomez-Nieto, L.C. Experimental model for reproduction of canine visceral leishmaniosis by Leishmania infantum. Vet. Parasitol. 2013, 192, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvapandiyan, A.; Dey, R.; Nylen, S.; Duncan, R.; Sacks, D.; Nakhasi, H.L. Intracellular replication-deficient Leishmania donovani induces long lasting protective immunity against visceral leishmaniasis. J. Immunol. 2009, 183, 1813–1820. [Google Scholar] [CrossRef] [Green Version]
- Dey, R.; Natarajan, G.; Bhattacharya, P.; Cummings, H.; Dagur, P.K.; Terrazas, C.; Selvapandiyan, A.; McCoy, J.P., Jr.; Duncan, R.; Satoskar, A.R.; et al. Characterization of cross-protection by genetically modified live-attenuated Leishmania donovani parasites against Leishmania mexicana. J. Immunol. 2014, 193, 3513–3527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nico, D.; Gomes, D.C.; Alves-Silva, M.V.; Freitas, E.O.; Morrot, A.; Bahia, D.; Palatnik, M.; Rodrigues, M.M.; Palatnik-de-Sousa, C.B. Cross-Protective immunity to Leishmania amazonensis is mediated by CD4+ and CD8+ epitopes of Leishmania donovani Nucleoside Hydrolase terminal domains. Front. Immunol. 2014, 5, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratti, J.E.; Ramos, T.D.; Pereira, J.C.; da Fonseca-Martins, A.M.; Maciel-Oliveira, D.; Oliveira-Silva, G.; de Mello, M.F.; Chaves, S.P.; Gomes, D.C.; Diaz, B.L.; et al. Efficacy of intranasal LaAg vaccine against Leishmania amazonensis infection in partially resistant C57BL/6 mice. Parasites Vectors 2016, 9, 534. [Google Scholar] [CrossRef] [Green Version]
- Bezerra, I.; Oliveira-Silva, G.; Braga, D.; de Mello, M.F.; Pratti, J.E.S.; Pereira, J.C.; da Fonseca-Martins, A.M.; Firmino-Cruz, L.; Maciel-Oliveira, D.; Ramos, T.D.; et al. Dietary Vitamin D3 Deficiency increases resistance to Leishmania (Leishmania) amazonensis infection in mice. Front. Cell. Infect. Microbiol. 2019, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho De Oliveira, B.; Duthie, M.S.; Alves Pereira, V.R. Vaccines for leishmaniasis and the implications of their development for American tegumentary leishmaniasis. Hum. Vaccines Immunother. 2020, 16, 919–930. [Google Scholar] [CrossRef]
- Campos, B.L.; Silva, T.N.; Ribeiro, S.P.; Carvalho, K.I.; Kallas, E.G.; Laurenti, M.D.; Passero, L.F. Analysis of iron superoxide dismutase-encoding DNA vaccine on the evolution of the Leishmania amazonensis experimental infection. Parasite Immunol. 2015, 37, 407–416. [Google Scholar] [CrossRef]
- de Matos Guedes, H.L.; Pinheiro, R.O.; Chaves, S.P.; De-Simone, S.G.; Rossi-Bergmann, B. Serine proteases of Leishmania amazonensis as immunomodulatory and disease-aggravating components of the crude LaAg vaccine. Vaccine 2010, 28, 5491–5496. [Google Scholar] [CrossRef]
- Alves-Silva, M.V.; Nico, D.; Morrot, A.; Palatnik, M.; Palatnik-de-Sousa, C.B. A Chimera Containing CD4+ and CD8+ T-Cell Epitopes of the Leishmania donovani Nucleoside Hydrolase (NH36) optimizes cross-protection against Leishmania amazonesis infection. Front. Immunol. 2017, 8, 100. [Google Scholar] [CrossRef] [Green Version]
- Nico, D.; Gomes, D.C.; Palatnik-de-Sousa, I.; Morrot, A.; Palatnik, M.; Palatnik-de-Sousa, C.B. Leishmania donovani Nucleoside Hydrolase terminal domains in cross-protective immunotherapy against Leishmania amazonensis murine infection. Front. Immunol. 2014, 5, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firmino-Cruz, L.; Ramos, T.D.; da Fonseca-Martins, A.M.; Maciel-Oliveira, D.; Oliveira-Silva, G.; Pratti, J.E.S.; Cavazzoni, C.; Chaves, S.P.; Oliveira Gomes, D.C.; Morrot, A.; et al. Immunomodulating role of IL-10-producing B cells in Leishmania amazonensis infection. Cell. Immunol. 2018, 334, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Mosser, D.M.; Zhang, X. Activation of the MAPK, ERK, following Leishmania amazonensis infection of macrophages. J. Immunol. 2007, 178, 1077–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norsworthy, N.B.; Sun, J.; Elnaiem, D.; Lanzaro, G.; Soong, L. Sand fly saliva enhances Leishmania amazonensis infection by modulating interleukin-10 production. Infect. Immun. 2004, 72, 1240–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, M.; Ramírez, L.; Solana, J.C.; Cook, E.C.L.; Hernández-García, E.; Requena, J.M.; Iborra, S. Inoculation of the Leishmania infantum HSP70-II Null Mutant Induces Long-Term Protection against L. amazonensis Infection in BALB/c Mice. Microorganisms 2021, 9, 363. https://doi.org/10.3390/microorganisms9020363
Soto M, Ramírez L, Solana JC, Cook ECL, Hernández-García E, Requena JM, Iborra S. Inoculation of the Leishmania infantum HSP70-II Null Mutant Induces Long-Term Protection against L. amazonensis Infection in BALB/c Mice. Microorganisms. 2021; 9(2):363. https://doi.org/10.3390/microorganisms9020363
Chicago/Turabian StyleSoto, Manuel, Laura Ramírez, José Carlos Solana, Emma C. L. Cook, Elena Hernández-García, José María Requena, and Salvador Iborra. 2021. "Inoculation of the Leishmania infantum HSP70-II Null Mutant Induces Long-Term Protection against L. amazonensis Infection in BALB/c Mice" Microorganisms 9, no. 2: 363. https://doi.org/10.3390/microorganisms9020363