
Genomic Characterization of Clinical Extensively Drug-Resistant Acinetobacter pittii Isolates

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Bacterial Identification
2.3. Antimicrobial Susceptibility Testing
2.4. Detection of Antimicrobial Resistance Genes
2.5. PCR-Based Replicon Typing
2.6. Multilocus Sequence Typing
2.7. Whole-Genome Sequencing and Analysis
2.8. Statistical Analysis
2.9. Nucleotide Sequence Accession Numbers
3. Results
3.1. Identification, Susceptibility, and Genotyping
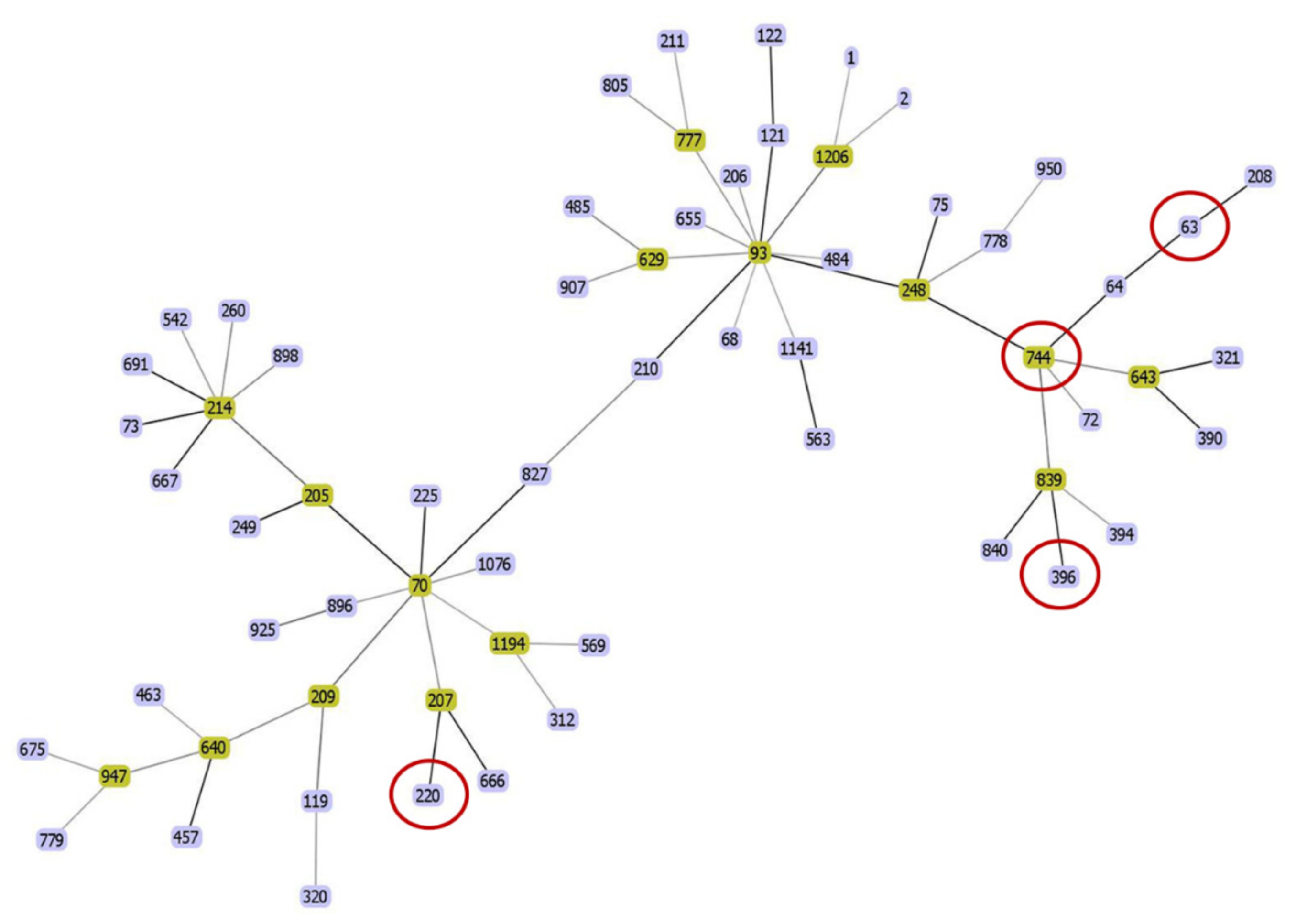
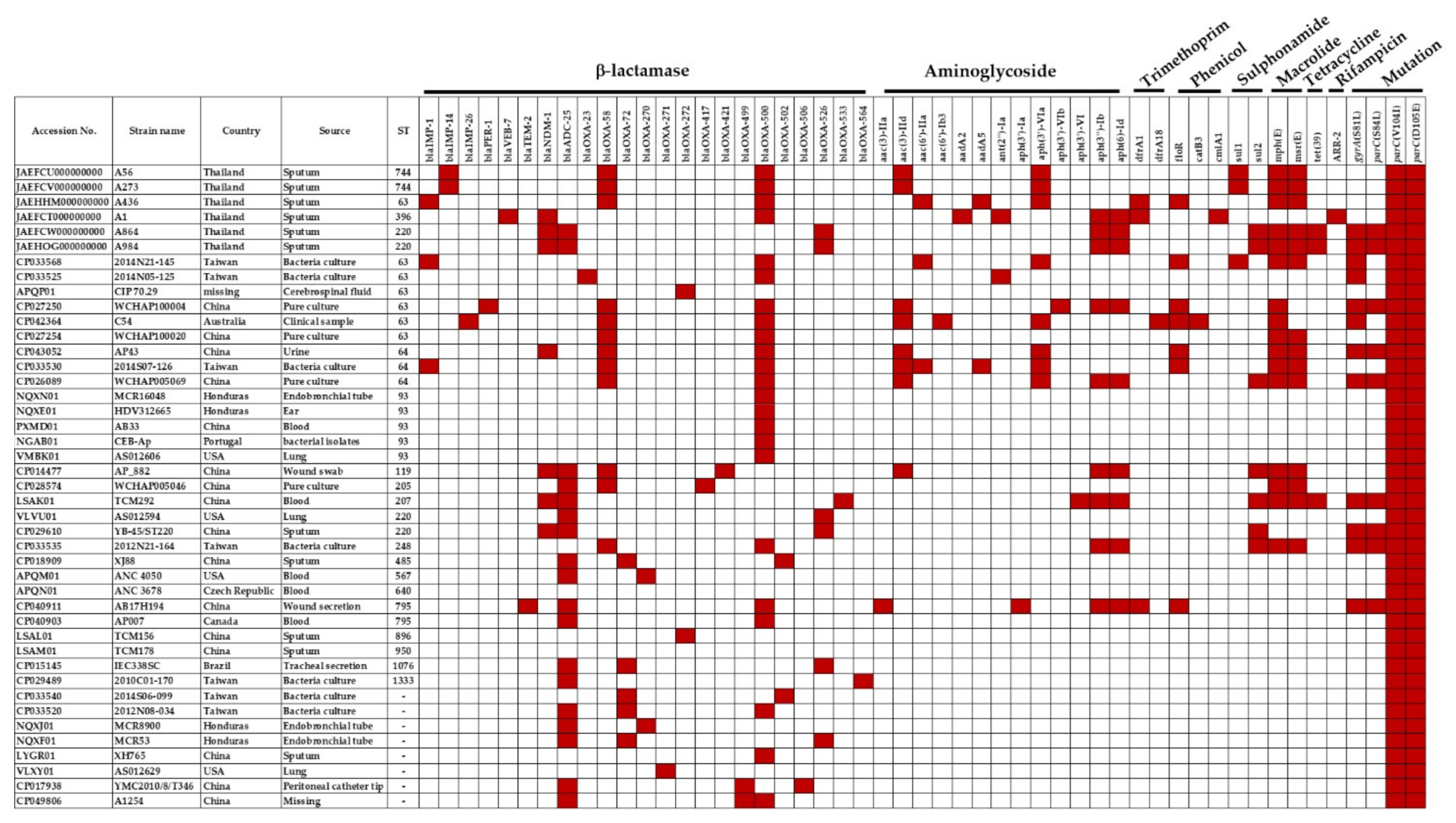
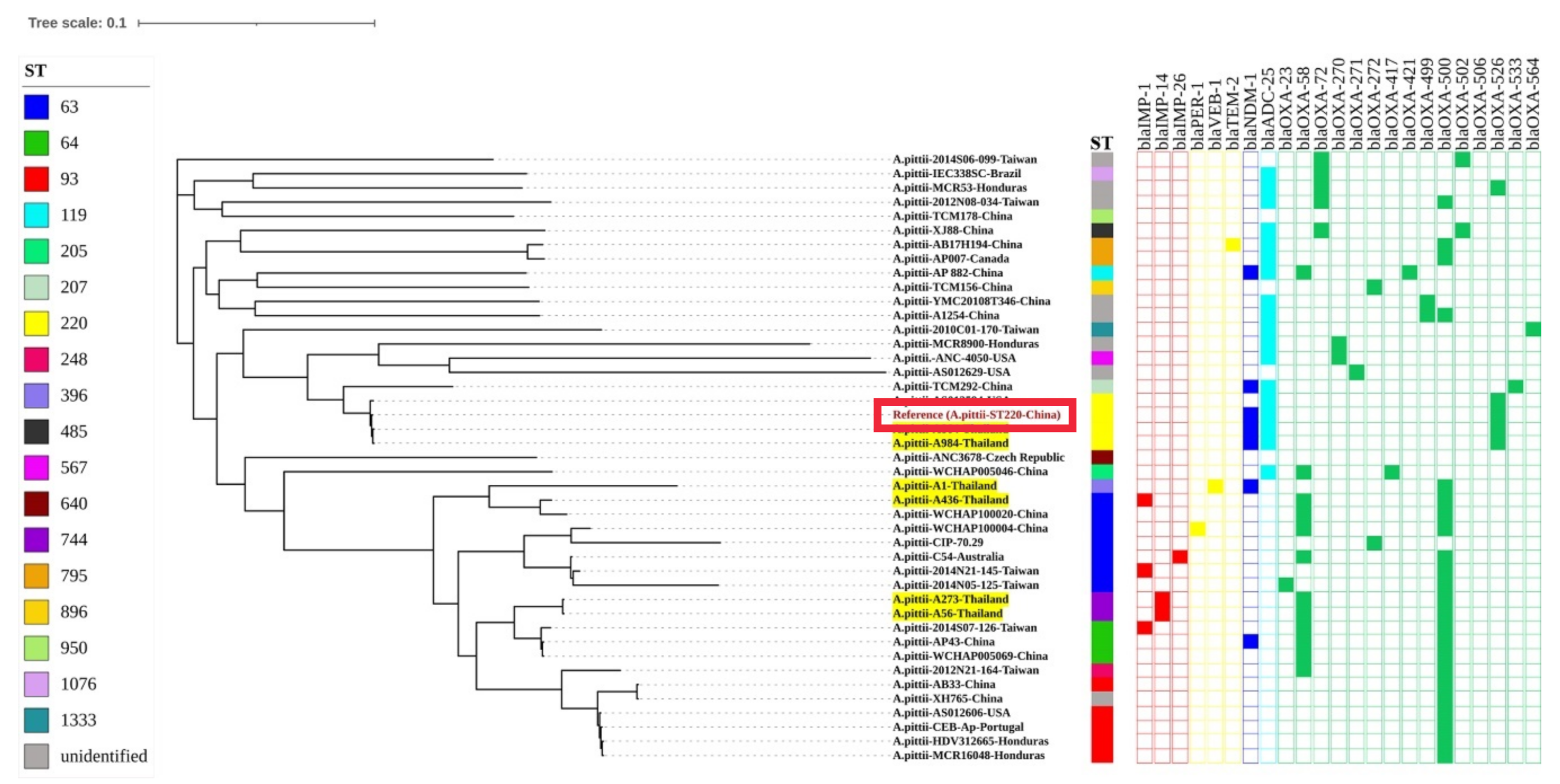
3.2. Genomic Characterization of Extensively-Drug Resistant A. pittii
3.3. Clinical Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.K.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus–Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov.(formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov.(formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2015, 65, 934–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgaya, C.; Marí-Almirall, M.; van Assche, A.; Assche, V.; Fern Andez-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; et al. Acinetobacter dijkshoorniae sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [CrossRef] [PubMed]
- Iimura, M.; Hayashi, W.; Arai, E.; Natori, T.; Horiuchi, K.; Go, M.; Tanaka, H.; Soga, E.; Nagano, Y.; Nagano, N. Detection of Acinetobacter pittii ST220 co-producing NDM-1 and OXA-820 carbapenemases from a hospital sink in a non-endemic country of NDM. J. Glob. Antimicrob. Resist. 2019. [Google Scholar] [CrossRef] [PubMed]
- Pailhoriès, H.; Tiry, C.; Eveillard, M.; Kempf, M. Acinetobacter pittii isolated more frequently than Acinetobacter baumannii in blood cultures: The experience of a French hospital. J. Hosp. Infect. 2018, 99, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.J.; Huang, W.C.; Liao, Y.C.; Wang, H.Y.; Lai, J.F.; Kuo, S.C.; Lauderdale, T.L.; Sytwu, H.K. Molecular epidemiology of emerging carbapenem resistance in Acinetobacter nosocomialis and Acinetobacter pittii in Taiwan, 2010 to 2014. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zander, E.; Fernández-González, A.; Schleicher, X.; Dammhayn, C.; Kamolvit, W.; Seifert, H.; Higgins, P.G. Worldwide dissemination of acquired carbapenem-hydrolysing class D β-lactamases in Acinetobacter spp. other than Acinetobacter baumannii. Int. J. Antimicrob. Agents 2014, 43, 375–377. [Google Scholar] [CrossRef] [PubMed]
- Singkham-In, U.; Chatsuwan, T. Mechanisms of carbapenem resistance in Acinetobacter pittii and Acinetobacter nosocomialis isolates from Thailand. J. Med. Microbiol. 2018, 67, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- Deglmann, R.C.; Kobs, V.C.; de Oliveira, D.; Burgardt, P.; de França, P.H.C.; Pillonetto, M. Earliest identification of new delhi metallo-β-lactamase 1 (NDM-1) in Acinetobacter pittii in Brazil. Rev. Soc. Bras. Med. Trop. 2019, 52. [Google Scholar] [CrossRef] [Green Version]
- Ang, G.Y.; Yu, C.Y.; Cheong, Y.M.; Yin, W.-F.; Chan, K.-G. Emergence of ST119 Acinetobacter pittii co-harbouring NDM-1 and OXA-58 in Malaysia. Int. J. Antimicrob. Agents 2016, 47, 168–169. [Google Scholar] [CrossRef]
- Sung, J.Y.; Koo, S.H.; Kim, S.; Kwon, G.C. Emergence of Acinetobacter pittii harboring New Delhi metallo-β-lactamase genes in Daejeon, Korea. Ann. Lab. Med. 2015, 35, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Gerner-Smidt, P.; Tjernberg, I.; Ursing, J. Reliability of phenotypic tests for identification of Acinetobacter species. J. Clin. Microbiol. 1991, 29, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Lehmann, M.; Wisplinghoff, H.; Seifert, H. gyrB Multiplex PCR to Differentiate between Acinetobacter calcoaceticus and Acinetobacter Genomic Species 3. J. Clin. Microbiol. 2010, 48, 4592–4594. [Google Scholar] [CrossRef] [Green Version]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by detection of the bla OXA-51-like carbapenemase gene intrinsic to this species. J. Clin. Microbiol. 2006, 44, 2974–2976. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: 30th Informational Supplement; CLSI Document M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Woodford, N.; Ellington, M.J.; Coelho, J.M.; Turton, J.F.; Ward, M.E.; Brown, S.; Amyes, S.G.B.; Livermore, D.M. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 2006, 27, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Cao, V.; Lambert, T.; Nhu, D.Q.; Loan, H.K.; Hoang, N.K.; Arlet, G.; Courvalin, P. Distribution of Extended-Spectrum-Lactamases in Clinical Isolates of Enterobacteriaceae in Vietnam. Antimicrob. Agents Chemother. 2002, 46, 3739–3743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatrongjit, R.; Kerdsin, A.; Akeda, Y.; Hamada, S. Detection of plasmid-mediated colistin-resistant and carbapenem-resistant genes by multiplex PCR. MethodsX 2018, 5, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Bertini, A.; Poirel, L.; Mugnier, P.D.; Villa, L.; Nordmann, P.; Carattoli, A. Characterization and PCR-based replicon typing of resistance plasmids in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4168–4177. [Google Scholar] [CrossRef] [Green Version]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Deekae, S.; Chayangsu, S.; Hatrongjit, R.; Chopjitt, P.; Takeuchi, D.; Akeda, Y.; Tomono, K.; Hamada, S. Genomic characterization of an emerging bla KPC-2 carrying Enterobacteriaceae clinical isolates in Thailand. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Moller, N.; Aarestrup, F.M. Rapid whole-genome sequencing for detection and characterization of microorganisms directly from clinical samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Feng, Y.; Zou, S.; Chen, H.; Yu, Y.; Ruan, Z. BacWGSTdb 2.0: A one-stop repository for bacterial whole-genome sequence typing and source tracking. Nucleic Acids Res. 2020. [Google Scholar] [CrossRef]
- Ruan, Z.; Feng, Y. BacWGSTdb, a database for genotyping and source tracking bacterial pathogens. Nucleic Acids Res. 2016, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014. [Google Scholar] [CrossRef] [Green Version]
- Vielva, L.; Ia De Toro, M.; Lanza, V.F.; De La Cruz, F. PLACNETw: A web-based tool for plasmid reconstruction from bacterial genomes. Bioinformatics 2017, 33, 3796–3798. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, 104984. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Web Serv. Issue Publ. Online 2019, 47. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Kiyasu, Y.; Hitomi, S.; Funayama, Y.; Saito, K.; Ishikawa, H. Characteristics of invasive Acinetobacter infection: A multicenter investigation with molecular identification of causative organisms. J. Infect. Chemother. 2020, 26, 475–482. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Paulus, T.; Lugenheim, M.; Stefanik, D.; Higgins, P.G.; Edmond, M.B.; Wenzel, R.P.; Seifert, H. Nosocomial bloodstream infections due to Acinetobacter baumannii, Acinetobacter pittii and Acinetobacter nosocomialis in the United States. J. Infect. 2012, 64, 282–290. [Google Scholar] [CrossRef]
- Liu, Y.M.; Lee, Y.T.; Kuo, S.C.; Chen, T.L.; Liu, C.P.; Liu, C.E. Comparison between bacteremia caused by Acinetobacter pittii and Acinetobacter nosocomialis. J. Microbiol. Immunol. Infect. 2017, 50, 62–67. [Google Scholar] [CrossRef]
- Chusri, S.; Chongsuvivatwong, V.; Rivera, J.I.; Silpapojakul, K.; Singkhamanan, K.; McNeil, E.; Doi, Y. Clinical outcomes of hospital-acquired infection with Acinetobacter nosocomialis and Acinetobacter pittii. Antimicrob. Agents Chemother. 2014, 58, 4172–4179. [Google Scholar] [CrossRef] [Green Version]
- Cosgaya, C.; Ratia, C.; Marí-almirall, M.; Rubio, L.; Higgins, P.G.; Seifert, H.; Roca, I.; Vila, J. In vitro and in vivo Virulence Potential of the Emergent Species of the Acinetobacter baumannii (Ab) Group. Front Microbiol. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Hamouda, A.; Towner, K.J.; Amyes, S.G.B. Novel genetic context of multiple blaOXA-58 genes in Acinetobacter genospecies 3. J. Antimicrob. Chemother. 2010, 65, 1586–1588. [Google Scholar] [CrossRef]
- Fu, Y.; Jiang, J.; Zhou, H.; Jiang, Y.; Fu, Y.; Yu, Y.; Zhou, J. Characterization of a novel plasmid type and various genetic contexts of blaOXA-58 in Acinetobacter spp. from multiple cities in China. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Koh, T.H.; Tan, T.T.; Khoo, C.T.; Ng, S.Y.; Tan, T.Y.; Hsu, L.Y.; Ooi, E.E.; Van Der Reijden, T.J.K.; Dijkshoorn, L. Acinetobacter calcoaceticus-Acinetobacter baumannii complex species in clinical specimens in Singapore. Epidemiol. Infect. 2012, 140, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Sheng, W.H.; Chen, Y.C.; Chang, S.C.; Hsia, K.C.; Li, S.Y. Differences in carbapenem resistance genes among Acinetobacter baumannii, Acinetobacter genospecies 3 and Acinetobacter genospecies 13TU in Taiwan. Int. J. Antimicrob. Agents 2010, 35, 439–443. [Google Scholar] [CrossRef]
- Chu, Y.W.; Afzal-Shah, M.; Houang, E.T.S.; Palepou, M.F.I.; Lyon, D.J.; Woodford, N.; Livermore, D.M. IMP-4, a novel metallo-β-lactamase from nosocomial Acinetobacter spp. Collected in Hong Kong between 1994 and 1998. Antimicrob. Agents Chemother. 2001, 45, 710–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerum, A.M.; Hansen, F.; Littauer, P. Use of whole-genome sequencing for characterisation of a ST119 NDM-1-producing Acinetobacter pittii from a patient in Denmark with no history of recent travel. Int. J. Antimicrob. Agents 2015, 46, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.; Poirel, L.; Martins, A.F.; Rozales, F.P.; Zavascki, A.P.; Barth, A.L.; Nordmann, P. Emergence of NDM-1-producing Acinetobacter pittii in Brazil. Int. J. Antimicrob. Agents 2015, 45, 444–445. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.K.; Jung, S.I.; Park, K.H.; Kim, S.H.; Ko, K.S. Characteristics of carbapenem-resistant Acinetobacter spp. other than Acinetobacter baumannii in South Korea. Int. J. Antimicrob. Agents 2012, 39, 81–85. [Google Scholar] [CrossRef]
- Pasteran, F.; Mora, M.M.; Albornoz, E.; Faccone, D.; Franco, R.; Ortellado, J.; Melgarejo, N.; Gomez, S.; Riquelme, I.; Matheu, J.; et al. Emergence of genetically unrelated NDM-1-producing Acinetobacter pittii strains in Paraguay. J. Antimicrob. Chemother. 2014, 69, 2575–2578. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Hotta, G.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Regional dissemination of Acinetobacter species harbouring metallo-β-lactamase genes in Japan. Clin. Microbiol. Infect. 2013, 19, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Hu, Y.Y.; Yang, X.F.; Gu, D.X.; Zhou, H.W.; Hu, Q.F.; Zhao, K.; Yu, S.F.; Chen, G.X. Emergence of NDM-producing non-baumannii Acinetobacter spp. isolated from China. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Kamolvit, W.; Derrington, P.; Paterson, D.L.; Sidjabat, H.E. A case of IMP-4-, OXA-421-, OXA-96-, and CARB-2-producing Acinetobacter pittii sequence type 119 in Australia. J. Clin. Microbiol. 2015, 53, 727–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, M.; Suzuki, S.; Yamane, K.; Suzuki, M.; Konda, T.; Arakawa, Y.; Shibayama, K. Distribution of carbapenem resistance determinants among epidemic and non-epidemic types of Acinetobacter species in Japan. J. Med. Microbiol. 2014, 63, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, P.; Huang, T.-D.; Rezende De Castro, R.; Bouchahrouf, W.; Glupczynski, Y. Could Acinetobacter pittii act as an NDM-1 reservoir for Enterobacteriaceae? J. Antimicrob. Chemother. 2013, 68, 2414–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhou, S. Draft genome sequence of an NDM-1-, OXA-421- and AmpC-producing Acinetobacter pittii ST220 in Anhui Province, China. J. Glob. Antimicrob. Resist. 2018, 14, 176–177. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Interspecies dissemination of a novel class 1 integron carrying blaIMP-19 among Acinetobacter species in Japan. J. Antimicrob. Chemother. 2011, 66, 2480–2483. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, T.; Yu, R.; Lü, X.; Zong, Z. Acinetobacter pittii and Acinetobacter nosocomialis among clinical isolates of the Acinetobacter calcoaceticus-baumannii complex in Sichuan, China. Diagn. Microbiol. Infect. Dis. 2013, 76, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, L.; Xu, L.; Chen, Y.; Chen, Y. Draft genome sequence of a multidrug-resistant New Delhi metallo-β-lactamase NDM-1-producing Acinetobacter pittii sequence type 207 isolate from China. J. Glob. Antimicrob. Resist. 2016, 6, 88–89. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Wu, L.; Chen, Y.; Xu, L. Draft genome sequence of a multidrug-resistant beta-lactamase OXA-357-producing Acinetobacter pittii ST865 clinical isolate from China. Braz. J. Microbiol. 2017, 48, 196–197. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Specimen | Age | Sex | Status | Disease | Underlying Disease | Outcome | ST | Genes | Plasmids | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| OXA | MBL | ||||||||||
| A1 | Sputum | 71 | M | Colonization | Ischemic heart disease with Atrial fabulation with Staphylococcus | Ischemic heart disease | Survive | 396 | 23, 51 | NDM-1 | GR12, GR8 |
| A56 | Sputum | 75 | M | Colonization | Fever of unknown origin | Unknown | Unknown | 744 | 58 | IMP-14 | GR3, GR12, GR8, GR16 |
| A273 | Sputum | 66 | M | Colonization | Hypotension with Pneumonia with CKD-5 * with DM * | CKD-5 * with DM * | Survive | 744 | 58 | IMP-14 | GR3 |
| A436 | Sputum | 34 | F | Infection | Heart failure with Respiratory failure with Atrial fabulation with Hypertension with Bacterial pneumonia | Hypertension | Survive | 63 | 58 | IMP-1 | GR3, GR12, GR8, GR6 |
| A864 | Sputum | 46 | M | Infection | Gastroenteritis with DM * | DM * | Survive | 220 | 10 | NDM-1 | GR3, GR12, GR8 |
| A984 | Ascitic Fluid | 73 | M | Infection | CKD-5 * with Gout with Hypertension Migraine with Liver cell carcinoma with Ascitic | CKD-5 * with Gout with Hypertension with CA Liver | Survive | 220 | 10 | NDM-1 | GR3, GR12, GR8 |
| Isolate No. | MIC (µg/L) | Disk Diffusion Assay (mm) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAZ | FEP | CTX | CRO | DOR | IPM | MEM | CL | GM | AMK | NET | CIP | SXT | TE | TZP | PIP | ||
| A1 | >32 | >32 | >32 | >32 | >4 | >8 | >8 | ≤1 | ≤2 | ≤8 | ≤8 | 0.12 | >4 | 20 | 12 | 11 | |
| (R) | (R) | (R) | (R) | (R) | (R) | (R) | (I) | (S) | (S) | (S) | (S) | (R) | (S) | (R) | (R) | MDR | |
| A56 | 32 | 32 | >32 | >32 | >4 | >8 | >8 | ≤1 | >8 | >32 | ≤8 | ≤0.06 | >4 | 20 | 19 | 18 | |
| (R) | (R) | (R) | (R) | (R) | (R) | (R) | (I) | (R) | (R) | (S) | (S) | (R) | (S) | (R) | (R) | XDR | |
| A273 | 16 | 32 | >32 | >32 | >4 | >8 | >8 | ≤1 | >8 | >32 | >16 | 1 | >4 | 20 | 17 | 16 | |
| (I) | (R) | (R) | (R) | (R) | (R) | (R) | (I) | (R) | (R) | (I) | (S) | (R) | (S) | (R) | (R) | XDR | |
| A436 | >32 | 32 | >32 | >32 | >4 | >8 | >8 | ≤1 | 8 | >32 | ≤8 | 0.12 | >4 | 23 | 18 | 16 | |
| (R) | (R) | (R) | (R) | (R) | (R) | (R) | (I) | (I) | (R) | (S) | (S) | (R) | (S) | (I) | (R) | XDR | |
| A864 | 32 | >32 | 32 | >32 | >4 | >8 | >8 | ≤1 | ≤8 | ≤8 | ≤8 | >2 | >4 | 9 | 12 | 12 | |
| (R) | (R) | (R) | (R) | (R) | (R) | (R) | (I) | (S) | (S) | (S) | (R) | (R) | (R) | (R) | (R) | XDR | |
| A984 | 32 | >32 | 32 | >32 | >4 | >8 | >8 | ≤1 | 4 | ≤8 | ≤8 | >2 | >4 | 7 | 12 | 12 | |
| (R) | (R) | (R) | (R) | (R) | (R) | (R) | (I) | (S) | (S) | (S) | (R) | (R) | (R) | (R) | (R) | XDR | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chopjitt, P.; Putthanachote, N.; Ungcharoen, R.; Hatrongjit, R.; Boueroy, P.; Akeda, Y.; Tomono, K.; Hamada, S.; Kerdsin, A. Genomic Characterization of Clinical Extensively Drug-Resistant Acinetobacter pittii Isolates. Microorganisms 2021, 9, 242. https://doi.org/10.3390/microorganisms9020242
Chopjitt P, Putthanachote N, Ungcharoen R, Hatrongjit R, Boueroy P, Akeda Y, Tomono K, Hamada S, Kerdsin A. Genomic Characterization of Clinical Extensively Drug-Resistant Acinetobacter pittii Isolates. Microorganisms. 2021; 9(2):242. https://doi.org/10.3390/microorganisms9020242
Chicago/Turabian StyleChopjitt, Peechanika, Nuntiput Putthanachote, Ratchadaporn Ungcharoen, Rujirat Hatrongjit, Parichart Boueroy, Yukihiro Akeda, Kazunori Tomono, Shigeyuki Hamada, and Anusak Kerdsin. 2021. "Genomic Characterization of Clinical Extensively Drug-Resistant Acinetobacter pittii Isolates" Microorganisms 9, no. 2: 242. https://doi.org/10.3390/microorganisms9020242