
Investigation of Lipolytic-Secreting Bacteria from an Artificially Polluted Soil Using a Modified Culture Method and Optimization of Their Lipase Production



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Lipase-Producing Bacteria
2.2. Screening of Microbial Lipase Production on Agar Plates with Tween-80
2.3. Screening of Microbial Lipase Production on Agar Plates with Olive Oil
2.4. Optimal Conditions for Lipase Production
2.4.1. Determination of a Suitable pH and Temperature
2.4.2. Determination of Optimal Carbon and Nitrogen Sources
2.4.3. Effect of Different Ions on Lipase Activity
2.4.4. Effect of Agitation on Lipase Production
2.5. Lipase Activity Assay
2.6. Molecular and Phylogenetic Analysis
3. Results
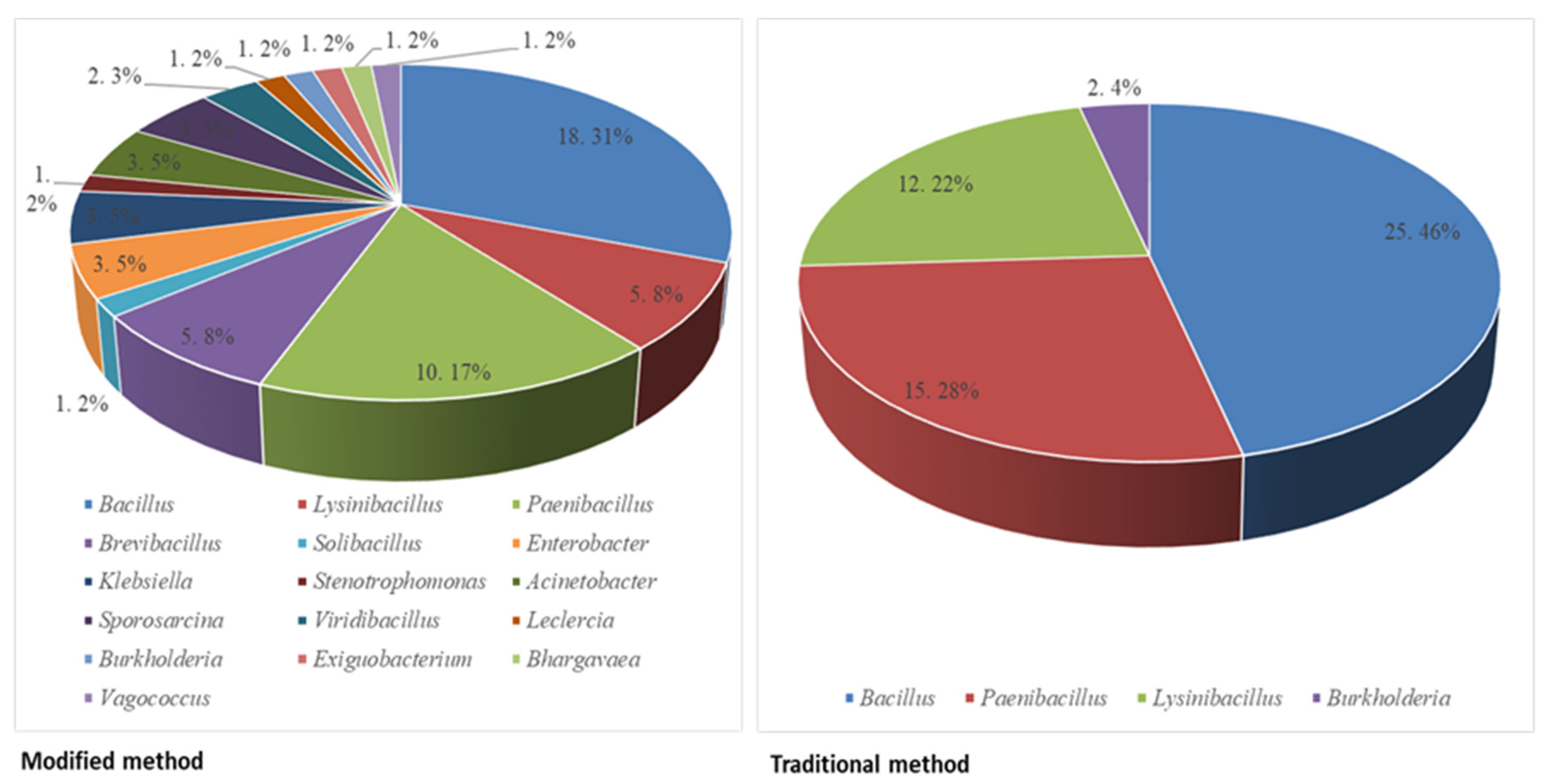
3.1. Phylogenetic Analysis
3.2. Quantitative Screening of Isolated Lipase-Producing Bacteria
3.3. Optimal Fermentation Conditions Affecting Lipase Production
3.3.1. The Effect of Temperature on Lipase Production
3.3.2. The Effect of pH on Lipase Production
3.3.3. The Effect of Culture Variable Carbon and Nitrogen Sources
3.3.4. The Effect of Metal Ions on Lipase Activity
3.3.5. The effect of Agitation on lipase production
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ergan, F.; Trani, M.; Andre, G. Production of glycerides from glycerol and fatty acid by immobilized lipases in non-aqueous media. Biotechnol. Bioeng. 1990, 35, 195–200. [Google Scholar] [CrossRef]
- Hasan, F.; Shah, A.A.; Hameed, A. Industrial applications of microbial lipases. Enzym. Microb. Technol. 2006, 39, 235–251. [Google Scholar] [CrossRef]
- Casas-Godoy, L.; Duquesne, S.; Bordes, F.; Sandoval, G.; Marty, A. Lipases: An overview. Methods Mol. Biol. 2012, 861, 3–30. [Google Scholar]
- Andualema, B.; Gessesse, A. Microbial Lipases and Their Industrial Applications: Review. Biotechnology 2012, 11, 100–118. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Olsen, C.E.; Parmar, V.S. Lipase catalyzed synthesis of optically enriched a-haloamides. Bioorg. Med. Chem. 2001, 9, 1345–1348. [Google Scholar]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef]
- Thirunavukarasu, K.; Edwinoliver, N.G.; Anbarasan, S.D.; Gowthaman, M.K.; Iefuji, H.; Kamini, N.R. Removal of triglyceride soil from fabrics by a novel lipase from Cryptococcus sp. S-2. Process. Biochem. 2008, 43, 701–706. [Google Scholar] [CrossRef]
- Chary, P.; Devi, Y. Lipases and their applications-An applied research in West Asia. Asian J. Biotechnol. Bioresour. Technol. 2018, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, N.; Revathi, D.; Sheelu, G.; Rani, K.Y.; Sridhar, S.; Mehtab, V.; Sumana, C. Recent advances on sources and industrial applications of lipases. Biotechnol. Prog. 2018, 34, 5–28. [Google Scholar] [CrossRef]
- Lindsay, D.; Brozel, V.S.; Mostert, J.F.; von Holy, A. Physiology of dairy associated Bacillus sp. over a wide pH range. Int. J. Food Microbiol. 2000, 54, 49–62. [Google Scholar] [CrossRef]
- Awad, G.; Mostafa, H.; Danial, E.N.; Abdelwahed, N.A.; Awad, H.M. Enhanced production of thermostable lipase from Bacillus cereus ASSCRC-P1 in waste frying oil based medium using statistical experimental design. J. Appl. Pharm. Sci. 2015, 5, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Silva-Bedoya, L.M.; Sánchez-Pinzón, M.S.; Cadavid-Restrepo, G.E.; MorenoHerrera, C.X. Bacterial community analysis of an industrial wastewater treatment plant in Colombia with screening for lipid-degrading microorganisms. Microbiol. Res. 2016, 192, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cai, W.; Wang, C.; Du, Z.; Lin, J.; Cai, J. Purification and characterization of alkaline lipase production by Pseudomonas aeruginosa HFE733 and application for biodegradation in food wastewater treatment. Biotechnol. Biotechnol. Equip. 2018, 32, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Sztajer, H.; Maliszewska, I.; Wieczorek, J. Production of exogenous lipases by bacteria, fungi, and actinomycetes. Enzym. Microb. Technol. 1998, 10, 492–497. [Google Scholar] [CrossRef]
- Wang, Y.; Srivastava, K.C.; Shen, G.J.; Wang, H.Y. Thermostable alkaline lipase from a newly isolated thermophilic Bacillus strain A30-1 (ATCC 53841). J. Ferment. Bioeng. 1995, 79, 433–438. [Google Scholar] [CrossRef]
- Beisson, F.; Tiss, A.; Riviere, C.; Verger, R. Methods for lipase detection and assay: A critical review. Eur. J. Lipid Sci. Technol. 2000, 133–153. [Google Scholar] [CrossRef]
- Ilesanmi, O.I.; Adekunle, A.E.; Omolaiye, J.A.; Olorode, E.M.; Ogunkanmi, A.L. Isolation, optimization and molecular characterization of lipase producing bacteria from contaminated soil. Sci. Afr. 2020, 8, e00279. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J. Cultivation of unculturable soil bacteria. Trends Biotechnol. 2012, 30, 475–484. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J. Improvement for isolation of soil bacteria by using common culture media. J. Pure Appl. Microbiol. 2016, 10, 49–59. [Google Scholar]
- Samad, M.Y.A.; Razak, C.N.A.; Salleh, A.B.; Yunus, W.M.Z.W.; Ampon, K.; Basri, M. A plate assay for primary screening of lipase activity. J. Microbiol. Methods 1989, 9, 51–56. [Google Scholar] [CrossRef]
- Yamada, K.; Ota, Y.; Machida, H. A modified method for lipase activity assay with emulsified olive oil as substrate. Nippon Nogeikagaku Kaishi 1962, 36, 860. [Google Scholar] [CrossRef]
- Lotti, M.; Monticelli, S.; Montisenos, J.L.; Brocca, S.; Valero, F.; Lafuente, J. Physiological control on the expression and secretion of Candida rugosa lipase. Chem. Phys. Lipids 1998, 93, 143–148. [Google Scholar] [CrossRef]
- Siddiqui, K.S.; Azhar, M.J.; Rashid, M.H.; Rajoka, M.I. Stability and identification of active-site residues of carboxymethylcellulases from Aspergillus niger and Cellulomonas biazotea. Folia. Microbiol. 1997, 42, 312–318. [Google Scholar] [CrossRef]
- Winkler, U.K.; Stuckman, M. Glycogen, hyaluronate, and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J. Bacteriol. 1979, 138, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Anbu, P.; Noh, M.; Kim, D.; Seo, J.; Hur, B.; Min, K.H. Screening and optimization of extracellular lipases by Acinetobacter species isolated from oil-contaminated soil in South Korea. Afr. J. Biotechnol. 2011, 10, 4147–4156. [Google Scholar]
- Jigajinni, S.K.; Meti, B.S. Immobilization optimization and characterization of immobilized lipase from Lysinibacillus macroides FS1 for biodiesel production. Int. J. Curr. Microbiol. App. Sci. 2021, 10, 232–245. [Google Scholar]
- Iboyo, A.; Asitok, A.; Ekpenyong, M.; Antai, S. Selection of Enterobacter cloacae Strain POPE6 for fermentative production of extracellular lipase on palm Kernel Oil processing effluent. Int. J. Sci. 2017, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.; Badar, R.; Khalique, N. Screening and optimization of submerged fermentation of lipolytic Aspergillus oryzae. BioResources 2019, 14, 7664–7674. [Google Scholar]
- Jia, J.; Yang, X.; Wu, Z.; Zhang, Q.; Lin, Z.; Guo, H.; Lin, C.S.K.; Wang, J.; Wang, Y. Optimization of fermentation medium for extracellular lipase production from Aspergillus niger using response surface methodology. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar]
- Guan, S.; Ji, C.; Zhou, T.; Li, J.; Ma, Q.; Niu, T. Aflatoxin B1 degradation by Stenotrophomonas maltophilia and other microbes. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.W.M.; Latif, H.A.E.; Ali, S.M. Production of Cold-Active Lipase by Free and Immobilized Marine Bacillus cereus HSS: Application in Wastewater Treatment. Front. Microbiol. 2018, 9, 2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adetunji, A.I.; Olaniran, A.O. Optimization of culture conditions for enhanced lipase production by an indigenous Bacillus aryabhattai SE3-PB using response surface methodology. Biotechnol. Biotechnol. Equip. 2018, 32, 1514–1526. [Google Scholar] [CrossRef] [Green Version]
- Devi, S.P.; Jha, D.K. Isolation of a lipolytic and proteolytic Bacillus licheniformis from refinery oily sludge and optimization of culture conditions for production of the enzymes. Microbiol. Biotechnol. Lett. 2020, 48, 515–524. [Google Scholar] [CrossRef]
- Ugaras, S.; Uzmes, S. Characterization of a newly identified lipase from a lipase-producing bacterium. Front. Biol. 2016, 11, 323–330. [Google Scholar] [CrossRef]
- Dharmsthiti, S.; Kuhasuntisuk, B. Lipase from Pseudomonas aeruginosa LP602: Biochemical properties and application for wastewater treatment. J. Ind. Microbiol. Biotechnol. 1998, 21, 75–80. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Salehghamari, E.; Khajeh, K.; Kabiri, M.; Naddaf, S. Production of an extracellular thermohalophilic lipase from a moderately halophilic bacterium, Salinivibrio sp. strain SA-2. J. Basic Microbiol. 2008, 48, 160–167. [Google Scholar] [CrossRef]
- Bora, L.; Bora, M. Optimization of extracellular thermophilic highly alkaline lipase from thermophilic Bacillus sp isolated from hotspring of Arunachal Pradesh, India. Braz. J. Microbiol. 2012, 43, 30–42. [Google Scholar] [CrossRef]
- Dharmsthiti, S.; Luchai, S. Production, purification and characterization of thermophilic lipase from Bacillus sp. THL027. FEMS Microbiol. Lett. 1999, 179, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Faiz, O.; Colak, A.; Saglam, N.; Çanakçi, S.; Beldüz, A.O. Determination and characterization of thermostable esterolytic activity from a novel thermophilic bacterium Anoxybacillus gonensis A4. J. Biochem. Mol. Biol. 2007, 40, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Khan, F.I.; Nizami, B.; Anwer, R.; Gu, K.R.; Bisetty, K.; Hassan, M.I.; Wei, D.Q. Structure prediction and functional analyses of a thermostable lipase obtained from Shewanella putrefaciens. J. Biomol. Struct. Dyn. 2017, 35, 23–2135. [Google Scholar] [CrossRef] [PubMed]
- Febriani; Aura, N.; Kemala, P.; Saidi, N.; Iqbalsyah, T.M. Novel thermostable lipase produced by a thermo-halophilic bacterium that catalyses hydrolytic and transesterification reactions. Heliyon 2020, 6, e04520. [Google Scholar] [CrossRef] [PubMed]
- Castilla, A.; Panizza, P.; Rodríguez, D.; Bonino, L.; Díaz, P.; Irazoqui, G.; Giordano, S.R. A novel thermophilic and halophilic esterase from Janibacter sp. R02, the first member of a new lipase family (Family XVII). Enzym. Microb. Technol. 2017, 98, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, A.; Tani, T.; Tominaga, Y. Purification and characterization of a novel thermostable lipase from Bacillus sp. J. Biochem. 1991, 109, 211–216. [Google Scholar] [PubMed]
- Kandasamy, R.; Kennedy, L.J.; Vidya, C.; Boopathy, R.; Sekaran, G. Immobilization of acidic lipase derived from Pseudomonas gessardii onto mesoporous activated carbon for the hydrolysis of olive oil. J. Mol. Catal. B Enzym. 2010, 62, 58–65. [Google Scholar] [CrossRef]
- Ramakrishnan, V.; Goveas, L.C.; Halami, P.M.; Narayan, B. Kinetic modeling, production and characterization of an acidic lipase produced by Enterococcus durans NCIM5427 from fish waste. J. Food Sci. Technol. 2015, 52, 1328–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kikon, K.; Upadhyay, A.; Kanwar, S.S.; Gupta, R. Production, purification, and characterization of lipase from thermophilic and alkaliphilic Bacillus coagulans BTS-3. Protein Expr. Purif. 2005, 41, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Lianghua, T.; Liming, X. Purification and partial characterization of a lipase from Bacillus coagulans ZJU318. Appl. Biochem. Biotechnol. 2005, 125, 139–146. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, Z.; Wang, B.; Kong, X.; Wang, Y.; Cao, S.; Fen, Y. Overexpression and characterization of a lipase from Bacillus subtilis. Protein Expr. Purif. 2006, 45, 22–29. [Google Scholar] [CrossRef]
- Nawani, N.; Khurana, J.; Kaur, J. A thermostable lipolytic enzyme from a thermophilic Bacillus sp.: Purification and characterization. Mol. Cell. Biochem. 2006, 290, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.; Souza, C.E.; Peclat, V.R.O.L.; Salgado, J.M.; Ribeiro, B.D.; Coelho, M.A.Z.; Venâncio, A.; Belo, I. Optimization of lipase production by Aspergillus ibericus from oilcakes and its application in esterification reactions. Food Bioprod. Process. 2017, 102, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Plou, F.J.; Ferrera, M.; Nuerob, O.M.; Calvoa, M.V.; Alcaldea, M.; FReyesb, F.; Ballesterosa, A. Analysis of Tween 80 as an esterase/lipase substrate for lipolytic activity assay. Biotechnol. Tech. 1998, 12, 183–186. [Google Scholar] [CrossRef]
- Cania, A.; Oetari, A.; Sjamsuridzal, W. Detection of olive oil and Tween 80 utilization by Rhizopus azygosporus UICC 539 at various temperatures. AIP Conf. Proc. 2020, 2242, 050009. [Google Scholar]
- Hermansyah, H.; Maresya, A.; Putri, D.N.; Sahlan, M.; Meyer, M. Production of dry extract lipase from Pseudomonas aeruginosa by the submerged fermentation method in palm oil mill effluent. Int. J. Technol. 2018, 9, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, T.; Cybulska, J.; Podleśny, M.; Frąc, M. Various perspectives on microbial lipase production using agri-food waste and renewable products. Agriculture 2021, 11, 540. [Google Scholar] [CrossRef]
- Golaki, B.P.; Aminzadeh, S.; Karkhane, A.A. Cloning, expression, purification, and characterization of lipase 3646 from thermophilic indigenous Cohnella sp. A01. Protein Expr. Purif. 2015, 109, 120–126. [Google Scholar] [CrossRef]
- Sharma, A.; Bardhan, D.; Patel, R. Optimization of physical parameters for lipase production from Arthrobacter sp. BGCC#490. Indian J. Biochem. Biophys. 2009, 46, 178–183. [Google Scholar]
- Sharma, P.; Sharma, N.; Pathania, S.; Handa, S. Purification and characterization of lipase by Bacillus methylotrophicus PS3 under submerged fermentation and its application in detergent industry. J. Genet. Eng. Biotechnol. 2017, 15, 369–377. [Google Scholar] [CrossRef]
- Shah, R.; Bhatt, S. Purification and characterization of lipase from Bacillus subtilis Pa2. J. Biochem. Technol. 2011, 3, 292–295. [Google Scholar]
- Mazhar, H.; Abbas, N.; Ali, S.; Sohail, A.; Hussain, Z.; Ali, S.S. Optimized production of lipase from Bacillus subtilis PCSIRNL-39. Afr. J. Biotechnol. 2017, 16, 1106–1115. [Google Scholar]
- Saraswat, R.; Verma, V.; Sistla, S.; Bhushan, I. Evaluation of alkali and thermotolerant lipase from an indigenous isolated Bacillus strain for detergent formulation. Electron. J. Biotechnol. 2017, 30, 33–38. [Google Scholar] [CrossRef]
- Ghori, M.I.; Iqbal, M.J.; Hameed, A. Characterization of a novel lipase from Bacillus sp. isolated from tannery waste. Braz. J. Microbiol. 2011, 42, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Barzkar, N.; Sohail, M.; Jahromi, S.T.; Gozari, M.; Poormozaffar, S.; Nahavandi, R.; Hafezieh, M. Marine bacterial esterases: Emerging biocatalysts for industrial applications. Appl. Biochem. Biotechnol. 2021, 193, 1187–1214. [Google Scholar] [CrossRef] [PubMed]
- Lesuisse, E.; Schanck, K.; Colson, C. Purification and preliminary characterization of the extracellular lipase of Bacillus subtilis 168, an extremely basic pH-tolerant enzyme. Eur. J. Biochem. 1993, 216, 155–160. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, M.; Xing, S.; Wu, T.; He, H.; Bielicki, J.K.; Chen, J. Identification and biochemical characterization of a novel hormone-sensitive lipase family esterase Est19 from the Antarctic bacterium Pseudomonas sp. E2-15. Biomolecules 2021, 11, 1552. [Google Scholar] [CrossRef]
- Kim, Y.O.; Khosasih, V.; Nam, B.H.; Lee, S.J.; Suwanto, A.; Kim, H.K. Gene cloning and catalytic characterization of cold-adapted lipase of Photobacterium sp. MA1-3 isolated from blood clam. J. Biosci. Bioeng. 2011, 114, 589–595. [Google Scholar] [CrossRef]
- Russi, A.E.; Brown, M.A. The Meninges: New therapeutic targets for multiple sclerosis. HHS Public Access. Transl. Res. 2016, 165, 255–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salwoom, L.; Rahman, R.N.Z.R.A.; Salleh, A.B.; Shariff, F.M.; Convey, P.; Pearce, D.; Ali, M.S.M. Isolation, characterisation, and lipase production of a cold-adapted bacterial strain Pseudomonas sp. LSK25 isolated from Signy Island, Antarctica. Molecules 2019, 24, 715. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Bacterial Strains | Isolates | Lipolytic Clearing Zone (mm) |
|---|---|---|
| 1 | Lysinibacillus PL33 | 22 |
| 2 | Lysinibacillus PL35 | 20 |
| 3 | Paenibacillus PL2 | 19 |
| 4 | Bacillus L8 | 18 |
| 5 | Bacillus FW2 | 17 |
| 6 | Enterobacter L7 | 15 |
| 7 | Bacillus N-PL4 | 14 |
| 8 | Enterobacter PL4 | 13 |
| 9 | Paenibacillus L2 | 12 |
| 10 | Stenotrophomonas N-PL7 | 10 |
| No. | Bacterial Strains | Relative Activity Affected by Metal Ions (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | Mg2+ | K+ | Fe2+ | Zn2+ | Cu2+ | Co2+ | Mn2+ | Sn2+ | ||
| 1 | Lysinibacillus PL33 | 100 | 100 | 98.2 | 100 | 55.7 | 65.2 | 35.6 | 100 | 65.8 |
| 2 | Lysinibacillus PL35 | 99.2 | 97.7 | 96.8 | 98.7 | 45.3 | 59.5 | 41.3 | 99.5 | 68.9 |
| 3 | Paenibacillus PL2 | 80.9 | 86.6 | 89.3 | 99.0 | 33.7 | 44.7 | 30.7 | 98.7 | 55.5 |
| 4 | Bacillus L8 | 85.3 | 98.4 | 85.7 | 98.6 | 46.8 | 35.8 | 45.8 | 99.2 | 66.3 |
| 5 | Bacillus FW2 | 98.8 | 100 | 100 | 100 | 58.3 | 49.3 | 55.6 | 99.6 | 68.7 |
| 6 | Enterobacter L7 | 98.5 | 97.5 | 95.8 | 82.5 | 23.8 | 27.5 | 16.7 | 63.4 | 42.6 |
| 7 | Bacillus N-PL4 | 90.3 | 100 | 85.3 | 80.9 | 34.9 | 55.8 | 22.5 | 79.8 | 43.9 |
| 8 | Enterobacter PL4 | 83.4 | 86.2 | 81.9 | 60.5 | 14.7 | 26.4 | 31.3 | 56.7 | 31.2 |
| 9 | Paenibacillus L2 | 84.6 | 99.4 | 78.7 | 75.8 | 22.5 | 13.9 | 23.8 | 60.3 | 20.8 |
| 10 | Stenotrophomonas N-PL7 | 85.9 | 87.7 | 83.5 | 80.6 | 13.8 | 20.5 | 17.8 | 64.6 | 17.7 |
| Microbial Source | Maximum Lipase Production (U/mL) | Application | References |
|---|---|---|---|
| Lysinibacillus macroides FS1 | 14.1 | Biocatalyst for production of biodiesel | [28] |
| Enterobacter cloacae Strain POPE6 | 2.031 | Apply to oil waste management | [29,30] |
| Stenotrophomonas maltophilia | 4559 | Degrade a harmful mycotoxin | [31,32] |
| Bacillus cereus HSS | 285 | Applied in waste water treatment | [33] |
| Bacillus aryabhattai SE3-PB | 265.82 | Potential biotechnological applications | [34] |
| Bacillus licheniformis | 37.6 | Industrial and environmental applications | [35] |
| Lysinibacillus PL33 | 480 | Industrial, environmental, and biological applications | This study |
| Lysinibacillus PL35 | 420 | Industrial, environmental, and biological applications | This study |
| Paenibacillus PL2 | 400 | Industrial, environmental, and biological applications | This study |
| Bacillus L8 | 390 | Industrial, environmental, and biological applications | This study |
| Bacillus FW2 | 380 | Industrial, environmental, and biological applications | This study |
| Enterobacter L7 | 340 | Industrial, environmental, and biological applications | This study |
| Bacillus N-PL4 | 330 | Industrial, environmental, and biological applications | This study |
| Enterobacter PL4 | 320 | Industrial, environmental, and biological applications | This study |
| Paenibacillus L2 | 290 | Industrial, environmental, and biological applications | This study |
| Stenotrophomonas N-PL7 | 170 | Industrial, environmental, and biological applications | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Investigation of Lipolytic-Secreting Bacteria from an Artificially Polluted Soil Using a Modified Culture Method and Optimization of Their Lipase Production. Microorganisms 2021, 9, 2590. https://doi.org/10.3390/microorganisms9122590
Pham VHT, Kim J, Chang S, Chung W. Investigation of Lipolytic-Secreting Bacteria from an Artificially Polluted Soil Using a Modified Culture Method and Optimization of Their Lipase Production. Microorganisms. 2021; 9(12):2590. https://doi.org/10.3390/microorganisms9122590
Chicago/Turabian StylePham, Van Hong Thi, Jaisoo Kim, Soonwoong Chang, and Woojin Chung. 2021. "Investigation of Lipolytic-Secreting Bacteria from an Artificially Polluted Soil Using a Modified Culture Method and Optimization of Their Lipase Production" Microorganisms 9, no. 12: 2590. https://doi.org/10.3390/microorganisms9122590