1. Introduction
Many studies have demonstrated that a high-fat diet (HFD) can modulate gut microbiota composition and render the gut more permissive to immunogenic bacterial products including lipopolysaccharide (LPS) [
1]. In addition to a Western high-fat diet, the composition of fat can also have an impact on gut microbial composition; for example, saturated, monounsaturated (MUFA), and industrial trans fats (iTFA) are characterized by reduced microbial diversity while polyunsaturated fats (PUFA) seem to exhibit a neutral effect on microbial diversity [
2]. However, the effects of naturally occurring trans fats on gut microbial composition have not been investigated.
Trans fats are molecules that contain a long hydrocarbon chain with a double bond in the trans configuration. Some trans fats are made synthetically through the partial hydrogenation of vegetable oils and have been widely used in industrial food production, called iTFA [
3,
4]. Vaccenic acid (VA), the most abundant trans isomer in meat and dairy products, is produced by microorganisms in the rumen through the incomplete biohydrogenation of polyunsaturated fatty acids (PUFAs), α-linoleic acid (ALA), and linolenic acid (LA) [
3]. Trans fats have received substantial negative attention over the past decade regarding undesirable health effects [
4,
5,
6]. Currently, trans fat content on a food label does not distinguish iTFA from naturally occurring ruminant trans fats such as vaccenic acid. Industrially produced vegetable fats have been demonstrated to increase risk of coronary heart disease through an increase in circulating total and LDL cholesterol [
7]. The structural differences between synthetic and natural trans fats have led to research investigating the specific properties and health effects between the two types of molecules. In particular, vaccenic acid has been shown to have a variety of health benefits including increasing insulin sensitivity as well as anti-inflammatory properties in the intestine [
8]. It has also been shown that it is possible to drastically increase the levels of VA in dairy and beef fat by supplementing the diet with flax [
9], therefore providing an opportunity to make beef healthier for the consumer.
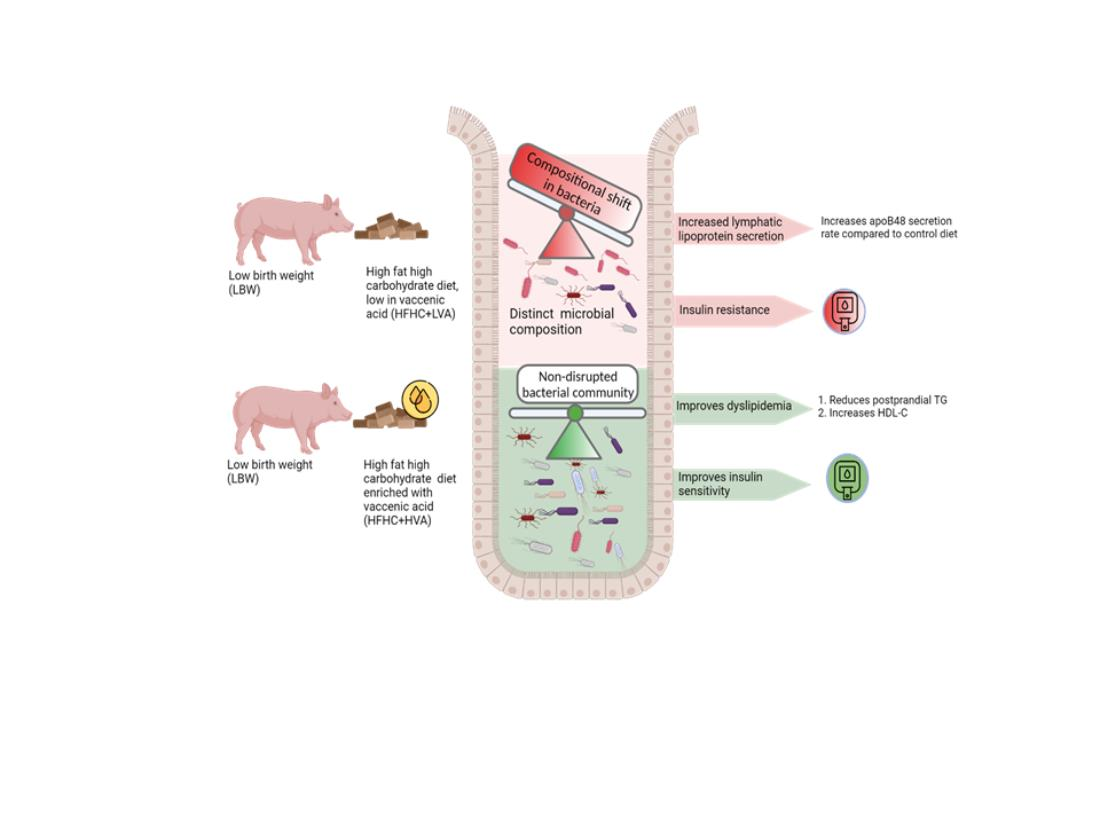
We have previously shown that low birth weight (LBW) pigs fed a Western diet (high fat, high fructose, and cholesterol; HFHC) develop metabolic complications (upregulate intestinal triglyceride absorption and secretion, develop dyslipidemia and muscular steatosis), display early signs of insulin resistance (IR), and induce changes in gut microbial composition relative to their LBW littermates on control diet [
10]. For this study we hypothesized that HFHC diet made with beef fat rich in VA (1.7%
w/w of diet or 10% of the fat) would protect the host from HFHC diet-induced microbial alterations and improve lipid metabolism relative to standard beef fat.
2. Materials and Methods
2.1. Animal Housing and Ethics
Piglets were obtained from the Swine Research and Technology Center (SRTC), Department of Agriculture, Food and Nutritional Science, University of Alberta, Canada. All procedures were approved by the University’s Animal Care and Use Committee—Livestock (ACUC) that follow guidelines from the Canadian Council on Animal Care (CCAC) (AUP00001184). Piglets used in the study were Duroc X Large White/Landrace cross. All food, water, temperature, and routine care were provided by trained staff in accordance with animal ethics guidelines. From birth, piglets were weighed weekly, feed intake was measured daily (once housed individually), and each piglet was socialized at minimum of twice weekly to ensure a low stress response during experimental collection.
2.2. Study Design
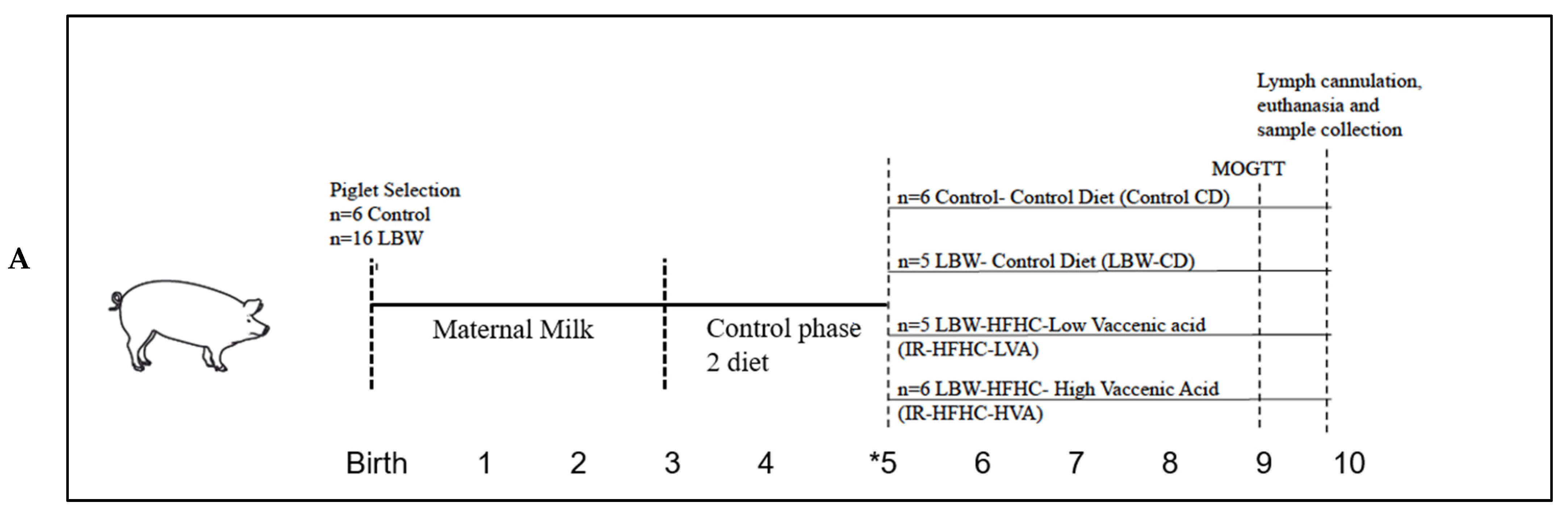
A mean litter weight and standard deviation (SD) was determined to find a 95% confidence interval (CI), categorizing piglets as LBW (less than the 95% CI) or Control (within the 95% CI). Newborn male piglets (n = 6 Control and n = 16 LBW) (Landrace-Large White × Duroc) were selected and weighed within 24 h of full-term birth from a total of 6 sows. Parity structure was as follows: sow #1: NBW n = 1, LBW n = 1; sow#2: NBW n = 2, LBW n = 2; sow#3: NBW n = 1, LBW n = 2; sow#4: NBW n = 2, LBW n = 4; sow#5: NBW n = 2, LBW n = 5; sow#6: NBW n = 2, LBW n = 2.
Pigs, balanced for litter, were gradually switched onto their treatment group diets (see
Table 1 and
Table 2 for diets) at week 5 with experimental groups as follows (metabolic type/diet): Control-Control Diet (Control-CD,
n = 6), LBW-Control Diet (LBW-CD,
n = 5), LBW fed high-fat, high-carbohydrate, low vaccenic acid diet denoted as Insulin Resistant-HFHC-LVA (IR-HFHC-LVA,
n = 5) and LBW fed high-fat, high-carbohydrate diet with high VA denoted as Insulin Resistant-HFHC-HVA (IR-HFHC-HVA,
n = 6). At 8 weeks of age the pigs were fasted overnight and jugular catheters were implanted as previously described [
11]. An adapted 2-step modified oral glucose tolerance and fat challenge test (MOGTT) was conducted at 9 weeks of age. Pigs were anesthetized at 10 weeks of age, lymph was sampled, and tissues were collected after euthanasia (
Figure 1).
2.3. Diet
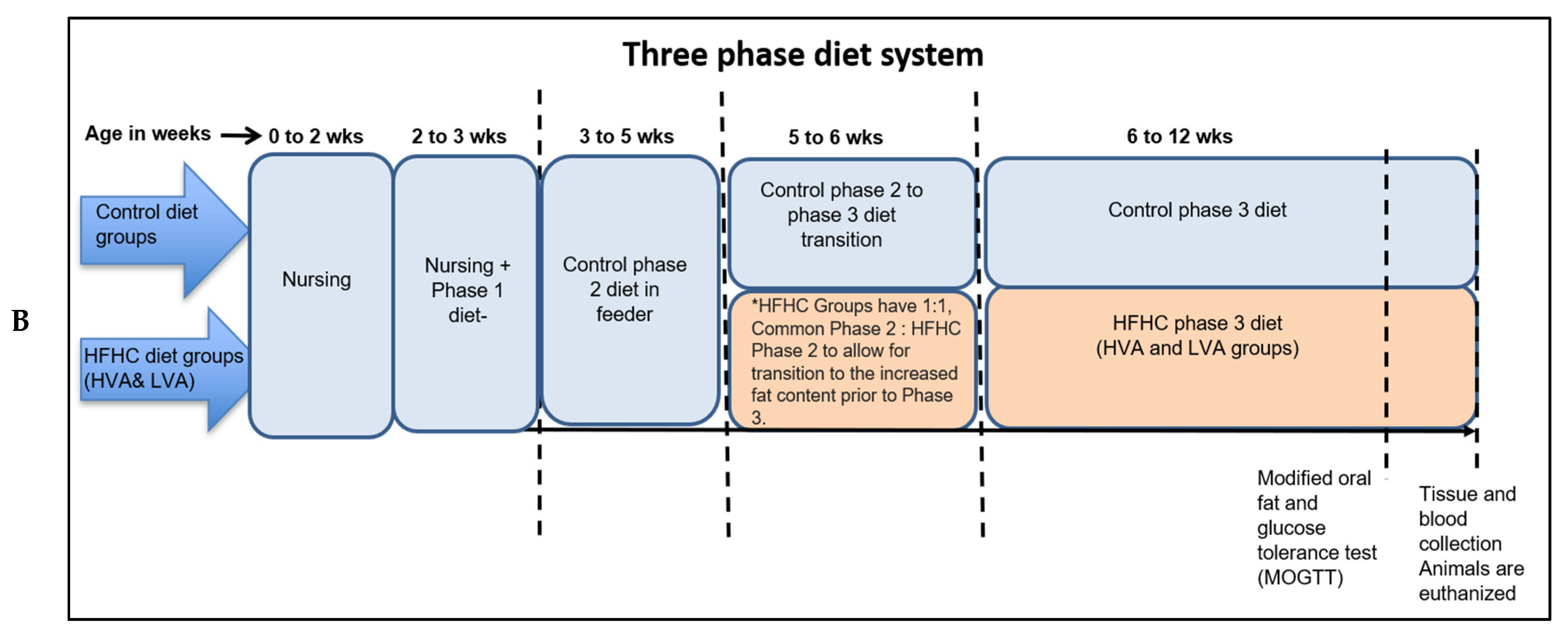
Pigs were fed diets in a three-phase system as shown in
Table 1 and
Figure 1. From 2–3 weeks of age pigs were fed a phase 1 diet, which is crumbled solid feed and is introduced by creep feeder. From 3 to 5 weeks of age all pigs were on a control phase 2 diet, fed from feeder. At 5 weeks of age, control diet groups were switched to phase 3 control diet. However, for the HFHC diet groups, the diets were gradually transitioned, to allow the pigs to accommodate to the changing diet. For HFHC groups, HFHC phase 2 diet (respective HVA and LVA diets) partially started at 5 weeks (mixed 1:1 with control Phase 2 diet), and was completely switched to HFHC phase 3 diet at 6 weeks Phase 2 and 3 of the HFHC-HVA diets were made using fat obtained from the Lacombe Research and Development Center, Lacombe, Alberta, Canada. Peri-renal fat was collected from steers fed extruded flaxseed (25%) and hay (75%) sequentially, as previously described [
12]. Phase 2 and 3 of the control and HFHC-LVA diets were made with peri-renal fat without VA, which was obtained from a commercial packing plant from steers fed a high barley-grain diet. Methods for fat collection and rendering are as previously described [
13]. Fatty acid composition of the diet (
Table 2) was determined as previously described using an adapted method [
13,
14]. The diets were formulated to meet or exceed nutrient requirements of starter pigs [
15] (
Table 1). Pigs were given ad libitum access to feed and water.
2.4. Modified Oral Glucose Tolerance Test
At 10 weeks of age the pigs were fasted overnight and general anesthetic was induced with isoflurane. Under sterile conditions, a catheter was implanted into the left jugular vein allowing for blood collection through a pouch on the back of each swine containing the catheter tubing. Post operatively, each pig was given buprenorphine (0.1 mg/kg every 4–8 h intramuscular for the first 12 h) to decrease pain while trimethoprim-sulpha (50 mg/kg intramuscular) was only given if there were any signs of post-surgical complications as per standard farm protocol [
16]. Each catheter was flushed once daily with 1.5 mL to 3 mL (100 IU/mL) heparin in saline solution depending on catheter length [
16].
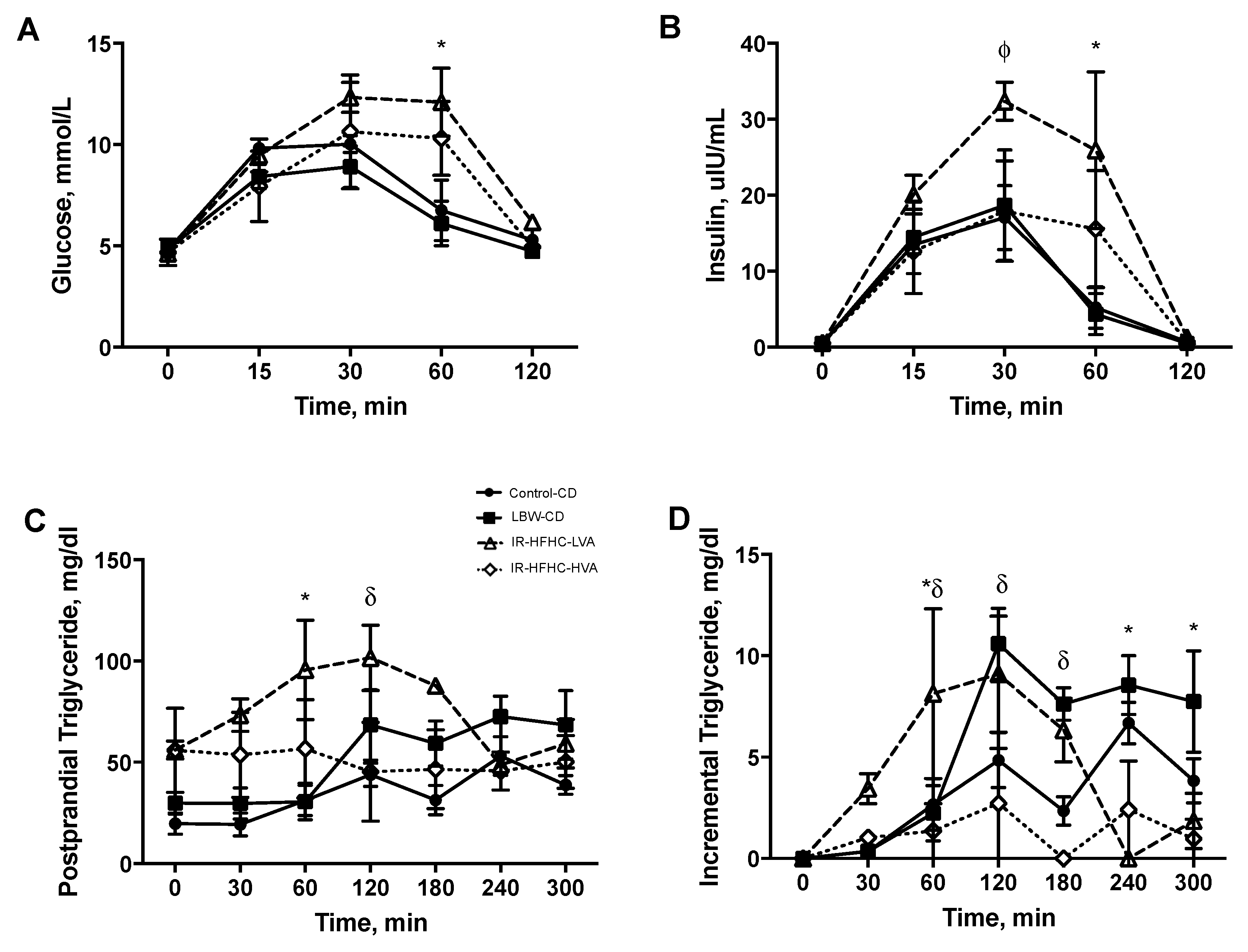
Piglets were fasted overnight at 11 weeks of age and subject to an adapted 2-step modified oral glucose tolerance and fat challenge test (MOGTT). Pigs were weighed and a fasted (time 0) sample of blood was collected via the jugular catheter. Pigs then consumed a 25 g control diet mixed with 1 g/kg body weight Devonshire cream (40% milk fat w/w) and 2 g/kg body weight 50% glucose solution. A second meal was given at the 120 min time point containing only 25 g control diet supplemented with Devonshire cream (1 g/kg body weight). Blood was collected at timed intervals (15, 30, 60, 120, 180, 240, 300 min) into EDTA-coated tubes and immediately tested for glucose. Plasma was isolated by centrifugation at 3500× g for 10 min at 4 °C and aliquots stored at −80 °C.
2.5. Mesenteric Lymph Duct Cannulation and Nascent Lymph Collection
At 13 weeks of age (7 weeks of diet), swine were fasted overnight, weighed and general anesthesia was induced. Anthropometric measurements were again taken as followed at birth. A cannula was implanted surgically into the superior mesenteric lymph duct and lymph was collected for 1 h into an EDTA-coated Vacutainer
TM as previously described [
17]. Total lymph volume as well as lymph flow rate (lymph volume/h) was recorded and the animal was terminated via exsanguination under anesthetic immediately upon completion of collection.
2.6. Sample Processing
Fasted blood was collected along with tissue samples, including heart, liver, kidney, adipose, muscle, intestine (jejunum and ileum) as well as intestinal scrapings (jejunum and ileum) and snap frozen in liquid nitrogen prior to storage at −80 °C. Prior to freezing, all tissue samples were flushed with ice-cold sterile saline. Blood and lymph were placed on ice until centrifuged at 3500× g for 10 min at 4 °C, aliquoted and stored at −80 °C.
2.7. Plasma and Lymph Biochemical Analysis
Plasma and lymph were assessed for lipid profiles using commercially available enzymatic colorimetric kits, including triglyceride (TG) (WAKO, Chemicals USA Inc., Richman, VA, USA, Cat#461-08992), glucose (WAKO, Cat#439-90901), LDL-cholesterol (WAKO, Cat#993-00404/999-00504), HDL-cholesterol (WAKO, Cat#993-72593/993-72693), and total cholesterol (TC, WAKO, Cat#439-17501). Glucose was measured at 505/600 nm wavelength while all other parameters were measured at 600/700 nm. Plasma insulin levels were assessed using a commercially available porcine-specific enzyme-linked immunosorbent assay (ELISA) with a detection limit of 0.007 ng/mL and intra-assay coefficient of variation (CV%) of 4.0% at 0.255 ng/mL (ALPCO, Salem, NH, USA) at 450 nm. For lipoprotein cholesterol and TG analysis by fast protein liquid chromatography (FPLC) gel filtration technique, fresh plasma was transferred to Agilent autosample vials and sent for analysis to the University of Alberta Lipidomics core facility with post-column detection of both cholesterol and TG.
2.8. Apolipoprotein B48 (apoB48) Quantification
Intestinally derived chylomicron (CM) particle concentration was determined by using an adapted immune Western blotting procedure [
18]. Lymph and plasma proteins were separated by SDS-PAGE on a 3–8% Tris-acetate polyacrylamide NuPage gel (Invitrogen, Camarillo, CA, USA). Separated proteins were then transferred to a polyvinylidene fluoride membrane (0.45 μm, ImmobilonTM, Millipore, Billerica, MA, USA). A goat polyclonal antibody specific for apoB (Santa Cruz Biotech, Santa Cruz, CA, USA) was incubated with the membrane overnight at 4 deg, which has specificity for both apoB100 and apoB48 isoforms. An anti-goat secondary antibody tagged with hydrogen peroxidise (Santa Cruz Biotech, Santa Cruz, CA, USA) was used (incubation time 1 h at room temperature) for detection purposes and Enhanced Chemiluminescence (ECL) (Amersham Biosciences Little Chalfont, Bucks, UK) intensity was used to quantify and compare with a known mass of purified rodent apoB48 protein.
2.9. Muscle, Liver and Intestinal Lipid Analysis by High Performance Liquid Chromatography (HPLC)
Tissue TG content was assessed by high-pressure liquid chromatography (HPLC). Tissue samples were homogenized using a standard buffer of 250 mM sucrose, 50 mM Tris, 1 mM EDTA with a pH of 7.4 and an added protease inhibitor cocktail tablet (Roche Diagnostics, Germany) using a polytron. Lipids were extracted using the modified Folch method [
19] and quantified by HPLC using phosphatidyldimethylethanolamine (PDME) as an internal standard at the Faculty of Medicine and Dentistry Lipid Analysis Core as previously described [
20].
2.10. Cecal Bacterial Community Characterization
Total DNA was extracted from cecal digesta using the QIAamp DNA Stool Mini Kit (Qiagen, Inc. Germantown, MD, USA) according to the manufacturer’s instructions, with the addition of a bead-beating step to lyse gram positive bacteria (FastPrep instrument, MP Biomedicals, Solon, OH, USA). DNA concentration was measured using a Quant-It™ PicoGreen
® dsDNA Assay Kit (Thermo Fischer Scientific, Waltham, MA, USA). Extracted DNA was amplified targeting the V3-V4 regions of the bacterial 16Sr RNA genes using KAPA HiFidelity Hot Start Polymerase (Kapa Biosystems, Inc., Wilmington, MA, USA) with the following conditions: 95 °C for 3 min, followed by 25 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, and 72 °C for 5 min with universal primers (Forward 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′ and Reverse 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-3′) for amplification [
21]. After amplification, AMPure XP beads were used to purify the 16S amplicon to remove primers and primer dimer species. Dual indices and Illumina sequencing adapters were attached using the Nextera XT Index Kit, followed by a second PCR cleanup and quantification. Samples were diluted to 4 nM and 5 μL aliquots of each diluted DNA sample were pooled and size-selected and denatured with NaOH, diluted to 4pM in Illumina HT1 buffer, spiked with 2-PhiX and heat denatured at 96 °C for 2 min. The library was sequenced using a MiSeq 600 cycle v3 kit on an Illumina MiSeq platform (San Diego, IL, USA), according to the manufacturer’s instructions.
2.11. Sequence Data Analysis
Sequence data were analyzed using a QIIME2 pipeline (MacQIIME v2019.4) [
22]. The Dada2-plugin was used for quality filtration and to generate feature tables; reads were truncated where average quality fell below 20 [
23]. Amplicon sequence variants were aligned with MAFFT to construct a phylogenetic tree [
24]. Greengenes reference database was used for taxonomic classification of bacterial 16S rRNA gene sequences [
25], with amplicons for the domain of interest extracted using primer sequences targeting the V3-V4 regions of the 16S rRNA gene [
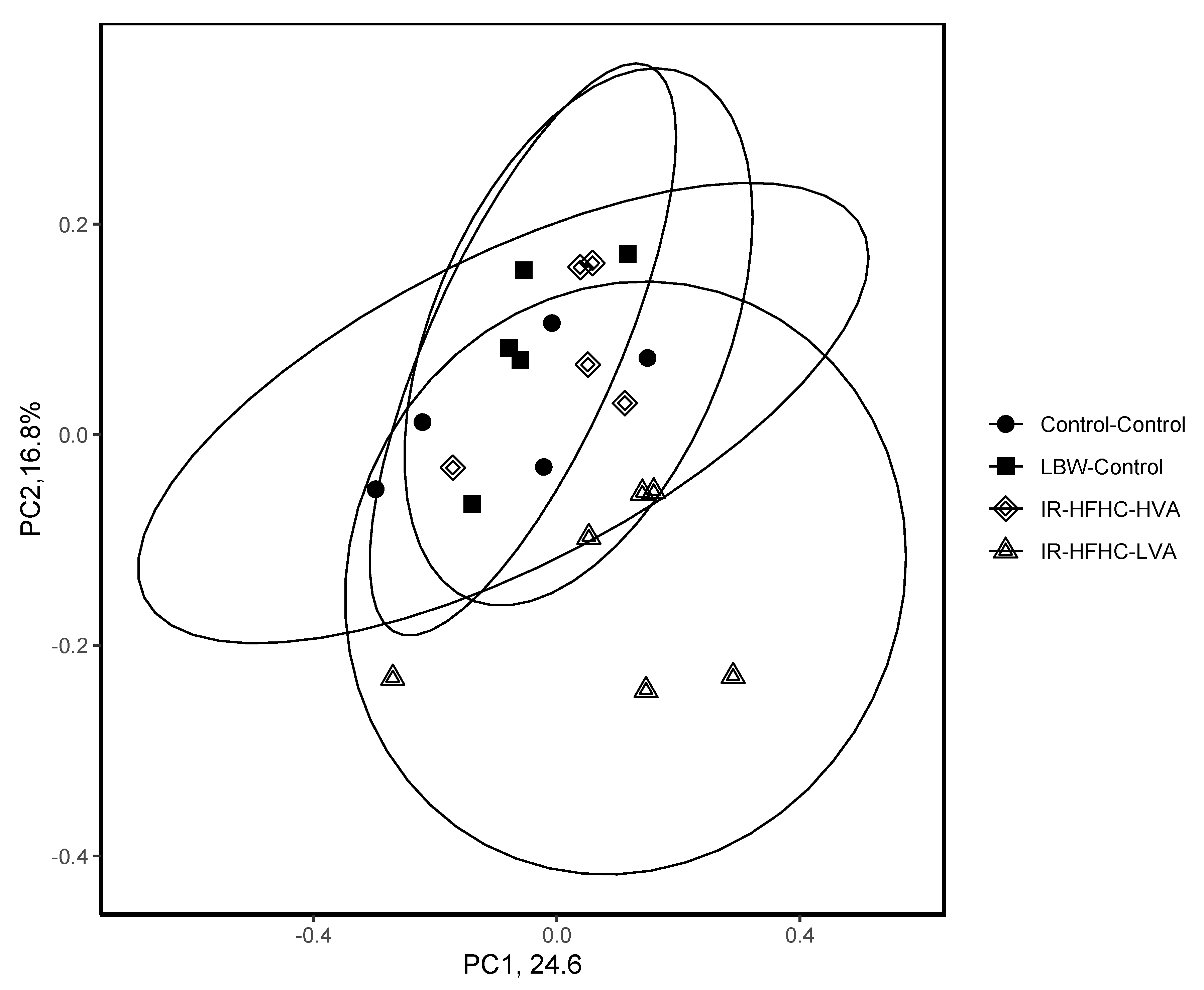
26]. The R package, phyloseq, was used to visualize the changes in microbial community alpha diversity using Chao1, Shannon, and Simpson indices and the overall microbial community structure using the Bray–Curtis dissimilarly and principal coordinate analysis (PCoA) (R, v3.6.1).
2.12. Circulating Endotoxin (LPS) Assay
Serum LPS concentrations were measured using PYROGENT-5000 kit as per manufacturer’s instructions (Lonza, Mississauga, ON, Canada). The absorbance was measured at 340 nm per minute for 1 h at 37 °C using a SpectraMax® M3 Microplate Reader (Molecular Devices, LLC. Sunnyvale, CA, USA). Reaction time was defined as the time required for the absorbance to increase 0.03 absorbance units.
2.13. Statistical Analysis
Statistical analysis was performed using the GraphPad Prism 7.02 software. Data were tested for normal distribution using the Shapiro–Wilk test due to its ability to test normality with a lower sample size. One-way ANOVA was used to detect differences between Control-CD and IR-CD, IR-CD and IR-HFHC, as well as IR-HFHC and IR-HFHC + VA treatment groups. Multiple comparisons were tested for using Fisher’s least significant difference (LSD) post-hoc analysis. Pair-matched values of each parameter at each time point for the postprandial curve was also analyzed for significant differences. Results are expressed as means ± SEM. Significance was set using a level of p value < 0.05 for all analyses.
Differences in bacterial community structure between treatments was compared using ADONIS. Alpha-diversity measures and relative abundances were compared using nonparametric Wilcoxon rank sum test with npar1way with pairwise multiple comparison analysis achieved using Dwass, Steel, Critchlow-Flingner multiple comparison procedure (SAS Studio University Edition, SAS Institute Inc., Cary, NC, USA).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}