
Characterization of Clinical Escherichia coli Strains Producing a Novel Shiga Toxin 2 Subtype in Sweden and Denmark

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Collection of STEC Strains and Clinical Data
2.3. Whole-Genome Sequencing (WGS) and Genome Assembly
2.4. WGS-Based Molecular Characterization
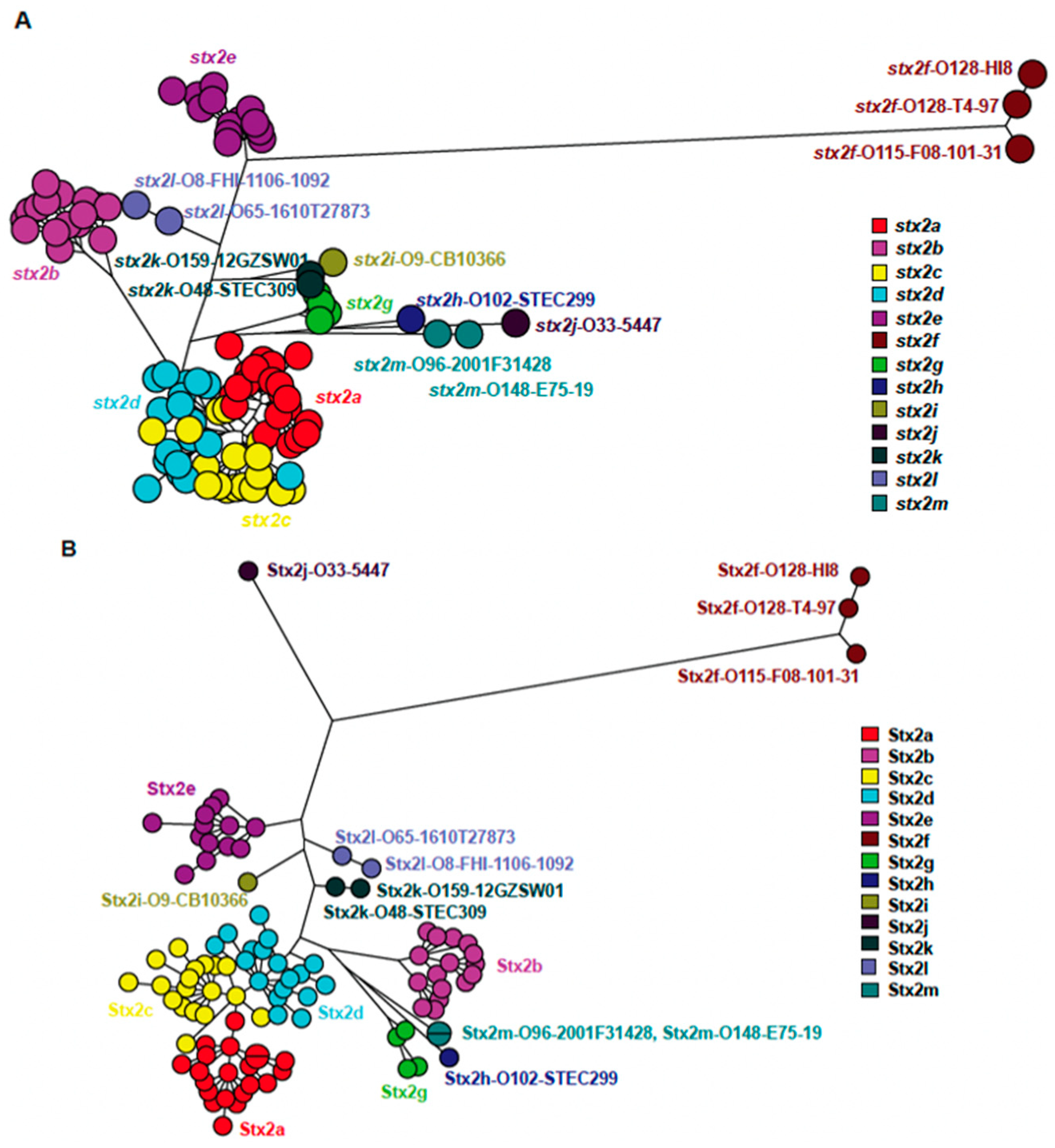
2.5. Stx Subtyping
2.6. Detection of Shiga Toxin Production
2.7. Antimicrobial Susceptibility Testing
2.8. Data Availability
3. Results
3.1. Identification of a Novel Stx2 Subtype in Clinical STEC Strains
3.2. Genetic Features of Stx2m-Producing Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar] [CrossRef] [PubMed]
- Obrig, T.G. Escherichia coli Shiga Toxin Mechanisms of Action in Renal Disease. Toxins 2010, 2, 2769–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, C.A.; Pellino, C.A.; Flagler, M.J.; Strasser, J.E.; Weiss, A.A. Shiga toxin subtypes display dramatic differences in potency. Infect Immun. 2011, 79, 1329–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasel, D.; Mellmann, A.; Cernela, N.; Hachler, H.; Fruth, A.; Khanna, N.; Egli, A.; Beckmann, C.; Hirsch, H.H.; Goldenberger, D.; et al. Hemolytic uremic syndrome in a 65-Year-old male linked to a very unusual type of stx2e- and eae-harboring O51:H49 shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2014, 52, 1301–1303. [Google Scholar] [CrossRef] [Green Version]
- De Rauw, K.; Jacobs, S.; Pierard, D. Twenty-seven years of screening for Shiga toxin-producing Escherichia coli in a university hospital. Brussels, Belgium, 1987–2014. PLoS ONE 2018, 13, e0199968. [Google Scholar] [CrossRef] [PubMed]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Fu, S.; Zhang, J.; Fan, R.; Xu, Y.; Sun, H.; He, X.; Xu, J.; Xiong, Y. Identification and pathogenomic analysis of an Escherichia coli strain producing a novel Shiga toxin 2 subtype. Sci. Rep. 2018, 8, 6756. [Google Scholar] [CrossRef] [PubMed]
- Lacher, D.W.; Gangiredla, J.; Patel, I.; Elkins, C.A.; Feng, P.C. Use of the Escherichia coli Identification Microarray for Characterizing the Health Risks of Shiga Toxin-Producing Escherichia coli Isolated from Foods. J. Food Prot. 2016, 79, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Bai, X.; Zhang, J.; Sun, H.; Fu, S.; Fan, R.; He, X.; Scheutz, F.; Matussek, A.; Xiong, Y. Escherichia coli strains producing a novel Shiga toxin 2 subtype circulate in China. Int. J. Med. Microbiol. 2020, 310, 151377. [Google Scholar] [CrossRef] [PubMed]
- EFSA Biohaz Panel; Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020, 18, e05967. [Google Scholar] [CrossRef]
- Lagerqvist, N.; Lof, E.; Enkirch, T.; Nilsson, P.; Roth, A.; Jernberg, C. Outbreak of gastroenteritis highlighting the diagnostic and epidemiological challenges of enteroinvasive Escherichia coli, County of Halland, Sweden, November 2017. Eurosurveillance 2020, 25, 1900466. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malberg Tetzschner, A.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico Genotyping of Escherichia coli Isolates for Extraintestinal Virulence Genes by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Souvorov, A.; Agarwala, R.; Lipman, D.J. SKESA: Strategic k-mer extension for scrupulous assemblies. Genome Biol. 2018, 19, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, A.; Lucchesi, P.M. Shiga toxins and stx phages: Highly diverse entities. Microbiology 2015, 161, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Scheutz, F. Vero Cytotoxin Producing Escherichia coli (VTEC) Isolated from Danish Patients. Ph.D. Thesis, Statens Serum Institut, Copenhagen, Denmark, 1997. [Google Scholar]
- Agger, M.; Scheutz, F.; Villumsen, S.; Molbak, K.; Petersen, A.M. Antibiotic treatment of verocytotoxin-producing Escherichia coli (VTEC) infection: A systematic review and a proposal. J. Antimicrob. Chemother. 2015, 70, 2440–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| E75_19 | E79_19 | 2001F31428 | |
|---|---|---|---|
| Clinical information | |||
| Source | Diarrheal patient | Healthy contact | Diarrheal patient |
| Infected region | Sweden | Sweden | Denmark |
| Isolation year | 2019 | 2019 | 2020 |
| Genetic characteristics | |||
| Serotype | O148:H39 | O148:H39 | O96:H19 |
| Stx2 subtype | Stx2m | Stx2m | Stx2m |
| Sequence type | 5825 | 5825 | 99 |
| Genome size (bp) | 5,514,783 | 5,565,491 | 4,948,862 |
| CG % | 50.39 | 50.43 | 50.94 |
| Accession number | SAMEA7019263 | SAMEA7019264 | SAMEA6873236 |
| Nucleotide\ Amino Acids | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Stx2a | 92.6 | 98.8 | 97.5 | 92.6 | 72.4 | 95.6 | 92.4 | 93.6 | 90.1 | 95.6 | 95.1 | 93.3 | |
| 2. Stx2b | 91.3 | 92.9 | 93.6 | 90.6 | 70.9 | 92.1 | 92.6 | 89.9 | 89.6 | 92.6 | 90.6 | 92.1 | |
| 3. Stx2c | 98.4 | 91.6 | 97.8 | 92.1 | 71.7 | 94.9 | 92.1 | 92.9 | 90.3 | 95.8 | 94.4 | 92.6 | |
| 4. Stx2d | 96.8 | 92.9 | 97.4 | 93.1 | 72.4 | 95.6 | 92.9 | 93.9 | 91.1 | 97.1 | 95.3 | 92.9 | |
| 5. Stx2e | 91.5 | 88.5 | 91.1 | 91.3 | 75.6 | 93.6 | 91.1 | 95.8 | 88.6 | 95.6 | 95.8 | 90.9 | |
| 6. Stx2f | 61.7 | 62.6 | 61.2 | 61.2 | 69.1 | 72.9 | 71.9 | 72.9 | 73.2 | 72.7 | 72.9 | 72.4 | |
| 7. Stx2g | 93.9 | 90.7 | 92.7 | 93.8 | 91.7 | 63.5 | 92.9 | 94.6 | 88.8 | 95.1 | 94.1 | 93.8 | |
| 8. Stx2h | 91.2 | 91.8 | 91.1 | 91.7 | 89.5 | 63.6 | 91.4 | 92.6 | 90.1 | 93.3 | 91.6 | 91.9 | |
| 9. Stx2i | 92.9 | 88.3 | 91.9 | 92.5 | 94.4 | 63.5 | 94.3 | 91.5 | 88.3 | 96.8 | 95.8 | 91.4 | |
| 10. Stx2j | 88.3 | 88.2 | 88.4 | 88.8 | 87.2 | 63.0 | 87.2 | 91.6 | 87.6 | 90.8 | 90.6 | 87.8 | |
| 11. Stx2k | 94.2 | 90.5 | 94.5 | 96.0 | 93.2 | 62.0 | 92.5 | 92.4 | 96.4 | 89.3 | 96.6 | 92.1 | |
| 12. Stx2l | 95.3 | 89.1 | 94.6 | 94.6 | 94.3 | 62.0 | 92.4 | 90.5 | 94.9 | 88.2 | 95.3 | 90.1 | |
| 13. Stx2m | 92.6 | 89.8 | 91.5 | 90.7 | 87.9 | 63.4 | 91.0 | 91.5 | 89.7 | 88.2 | 90.0 | 89.0 |
| Genus | Function | E75_19 | E79_19 | 2001F31428 |
|---|---|---|---|---|
| stx2 | Shiga toxin 2 | + | + | + |
| iha | Adherence protein | + | + | - |
| astA | Heat-stable enterotoxin 1 | + | + | + |
| lpfA | Long polar fimbriae | - | - | + |
| iss | Increased serum survival | - | - | + |
| celb | Endonuclease colicin E2 | - | - | + |
| ompT | Outer membrane protease | - | - | + |
| terC | Tellurium ion resistance protein | + | + | + |
| hra | Heat-resistant agglutinin | + | + | - |
| chuA | Outer membrane hemin receptor | + | + | - |
| eilA | HilA (an invasion protein) homolog | + | + | - |
| traT | Outer membrane protein complement resistance | + | + | - |
| kpsMII_K5 | Polysialic acid transport protein | + | + | - |
| mchB | Microcin H47 part of colicin H | + | + | - |
| mchC | Microcin H47 biosynthesis protein | + | + | - |
| mchF | ABC transporter protein MchF | + | + | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Scheutz, F.; Dahlgren, H.M.; Hedenström, I.; Jernberg, C. Characterization of Clinical Escherichia coli Strains Producing a Novel Shiga Toxin 2 Subtype in Sweden and Denmark. Microorganisms 2021, 9, 2374. https://doi.org/10.3390/microorganisms9112374
Bai X, Scheutz F, Dahlgren HM, Hedenström I, Jernberg C. Characterization of Clinical Escherichia coli Strains Producing a Novel Shiga Toxin 2 Subtype in Sweden and Denmark. Microorganisms. 2021; 9(11):2374. https://doi.org/10.3390/microorganisms9112374
Chicago/Turabian StyleBai, Xiangning, Flemming Scheutz, Henrik Mellström Dahlgren, Ingela Hedenström, and Cecilia Jernberg. 2021. "Characterization of Clinical Escherichia coli Strains Producing a Novel Shiga Toxin 2 Subtype in Sweden and Denmark" Microorganisms 9, no. 11: 2374. https://doi.org/10.3390/microorganisms9112374