
Bhalin, an Essential Cytoskeleton-Associated Protein of Trypanosoma brucei Linking TbBILBO1 of the Flagellar Pocket Collar with the Hook Complex

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cell Lines, Culture and Transfection
2.2. Anti-BHALIN Antibody Production
2.3. Reporter Uptake Assays
2.4. Immunofluorescence Assay
2.5. Molecular Biology and Cloning
2.6. Cloning Full-Length BHALIN and Truncations for Transfection into U-2 OS Cells
2.7. Extraction of RNA
2.8. Semi-Quantitative RT-PCR
2.9. Western Blot Analysis
2.10. Yeast Two-Hybrid Assay
3. Results
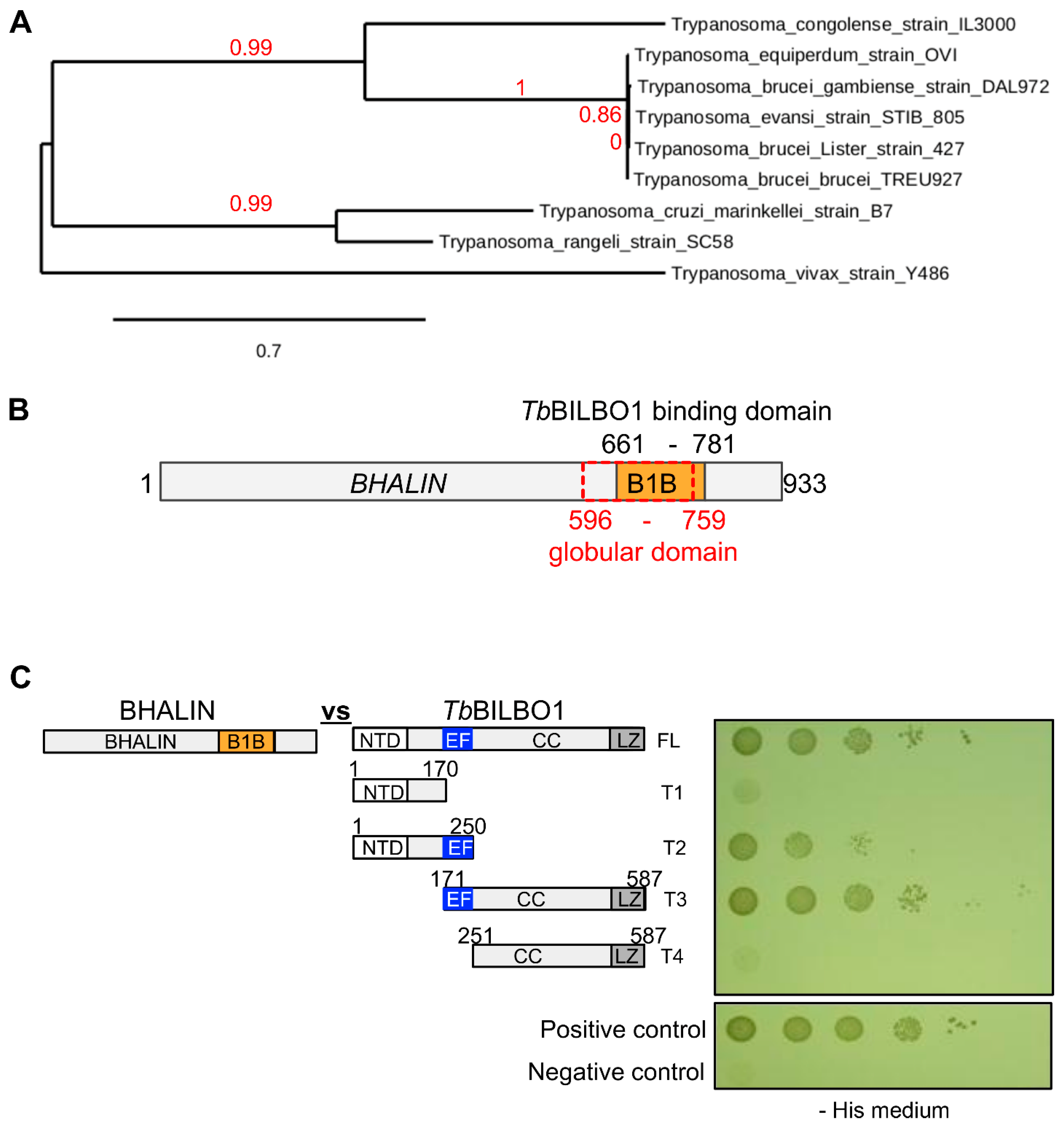
3.1. BHALIN Is a Trypanosomatid Specific Protein
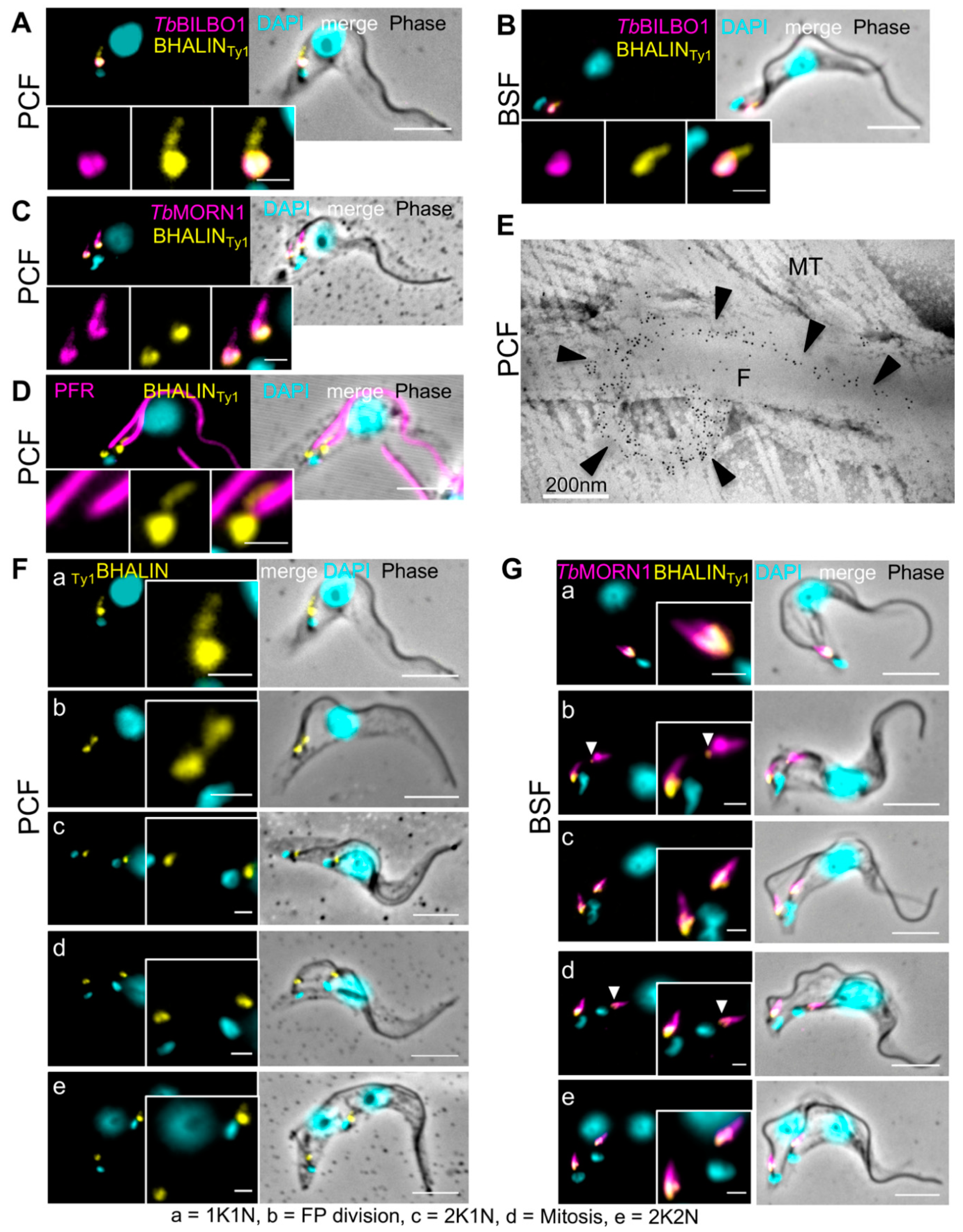
3.2. BHALIN Forms a Hook-Shaped Structure at the Hook Complex
3.3. BHALIN Is Present at the Hook Complex throughout the Cell Cycle
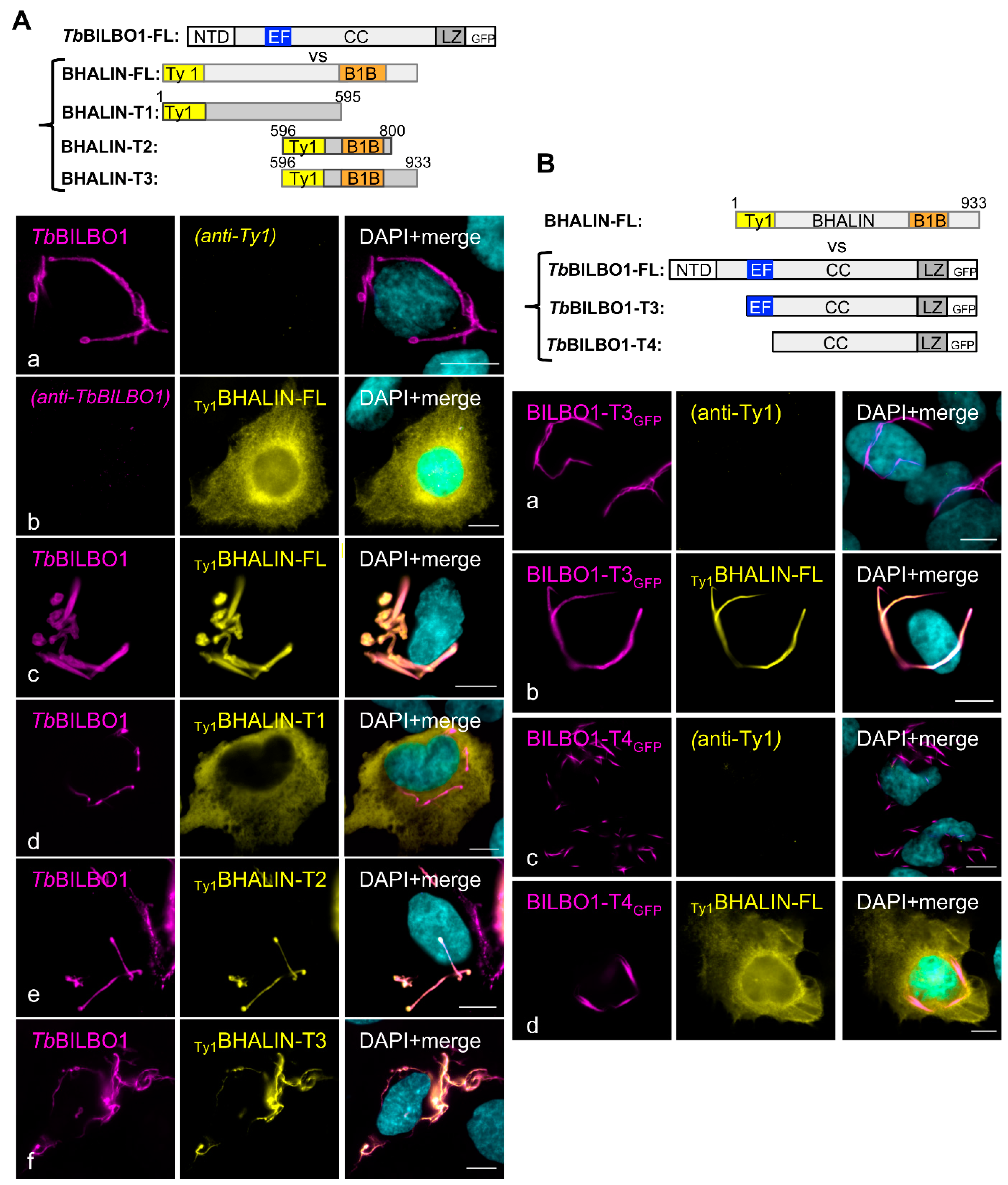
3.4. Binding of BHALIN Modifies the Shape of the TbBILBO1 Filaments and Requires the TbBILBO1 EF-Hand Domain
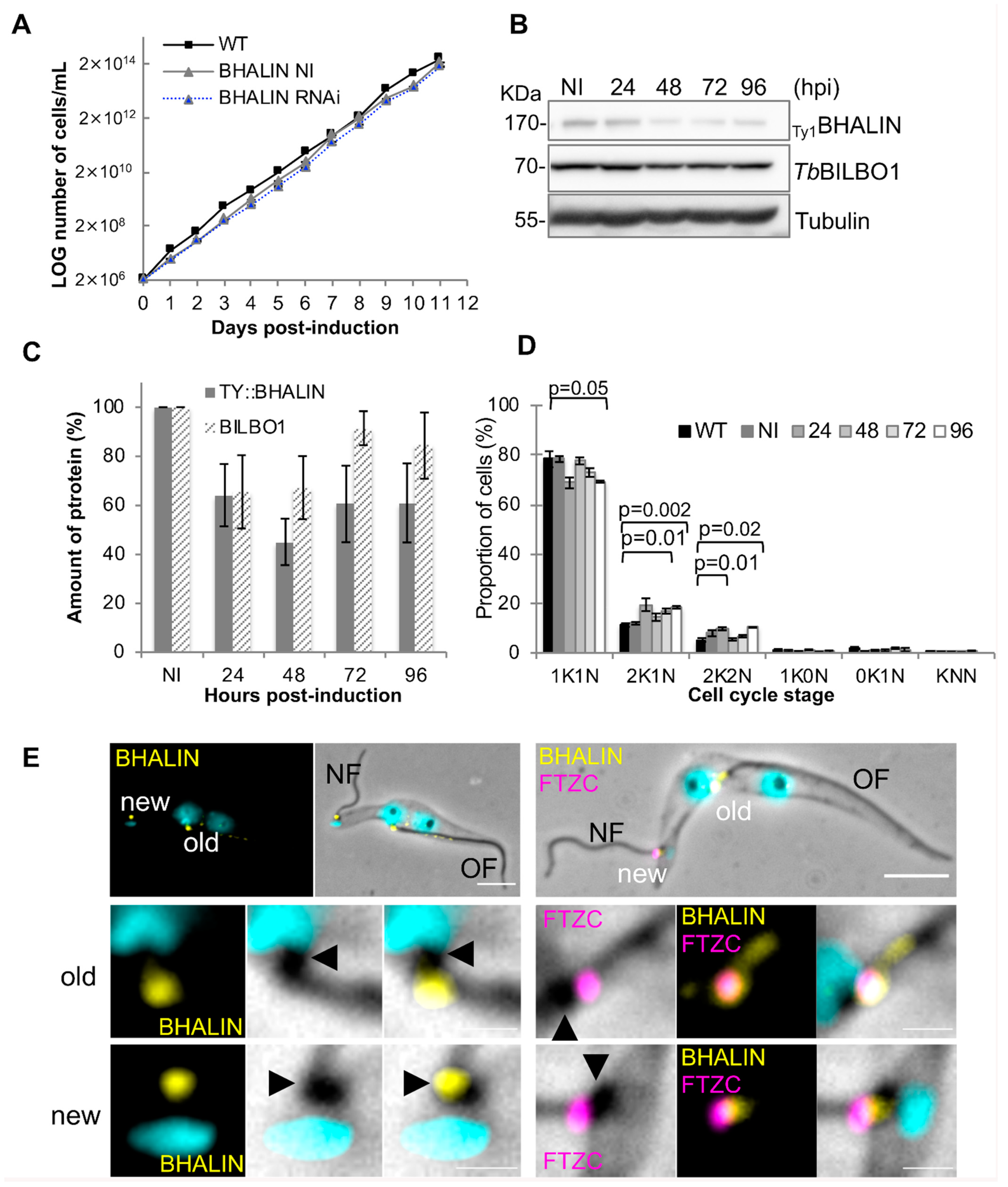
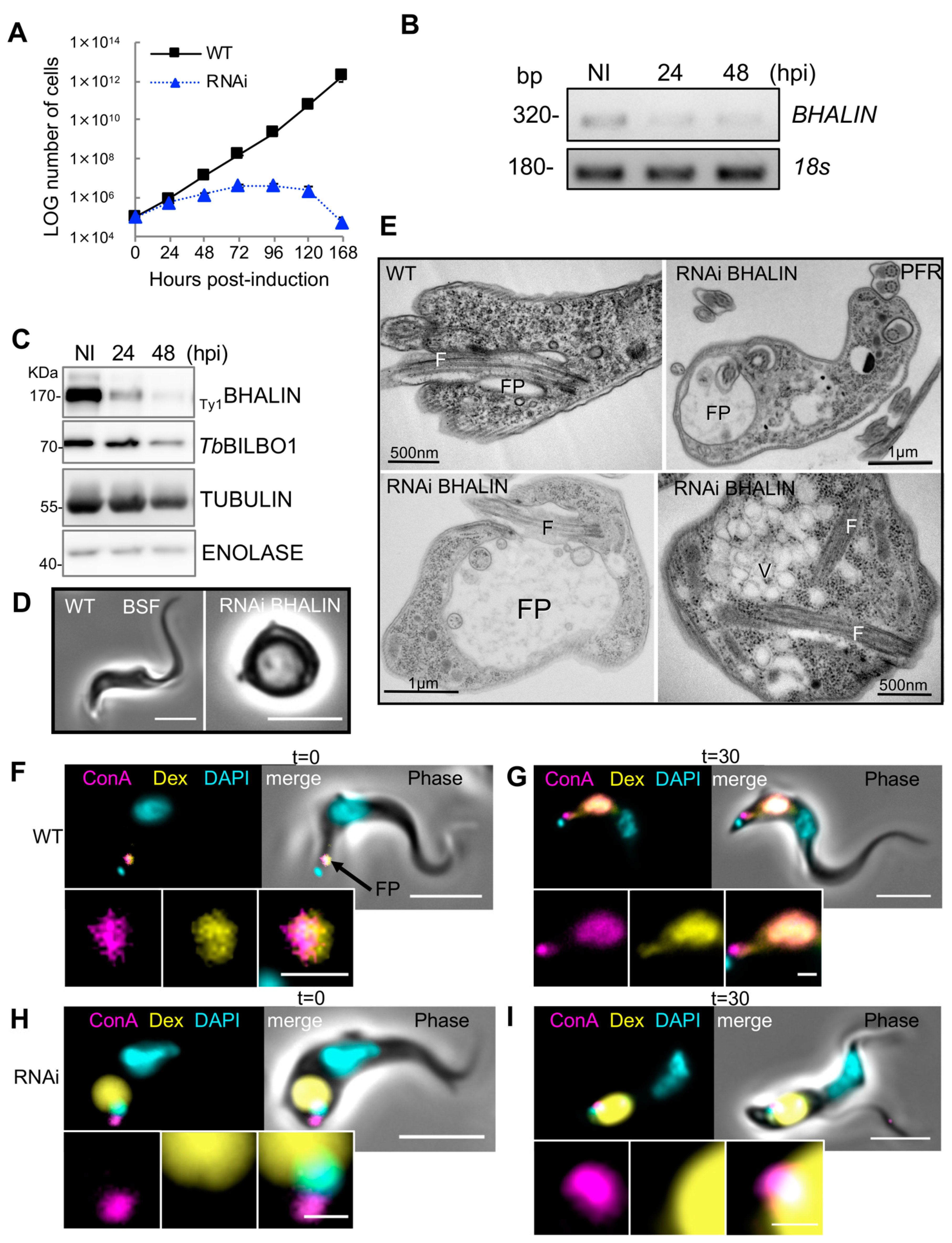
3.5. Knockdown of BHALIN in Procyclic Forms Leads to an Altered Cell Cycle Profile
3.6. Knockdown of BHALIN in Bloodstream Forms Is Lethal and Induces a “BigEye” Phenotype
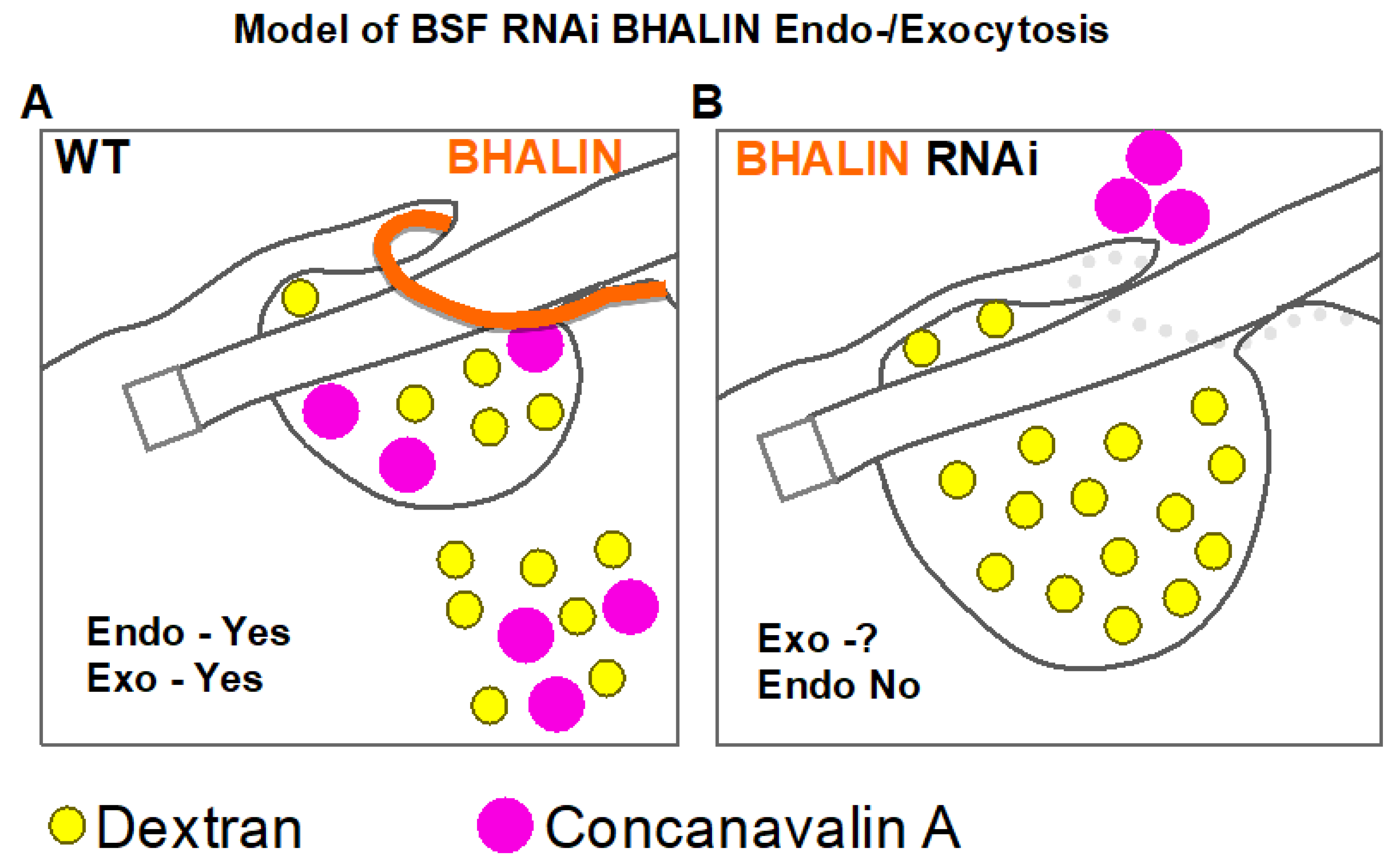
3.7. Endocytosis Is Perturbed in BSF BHALIN RNAi Cells
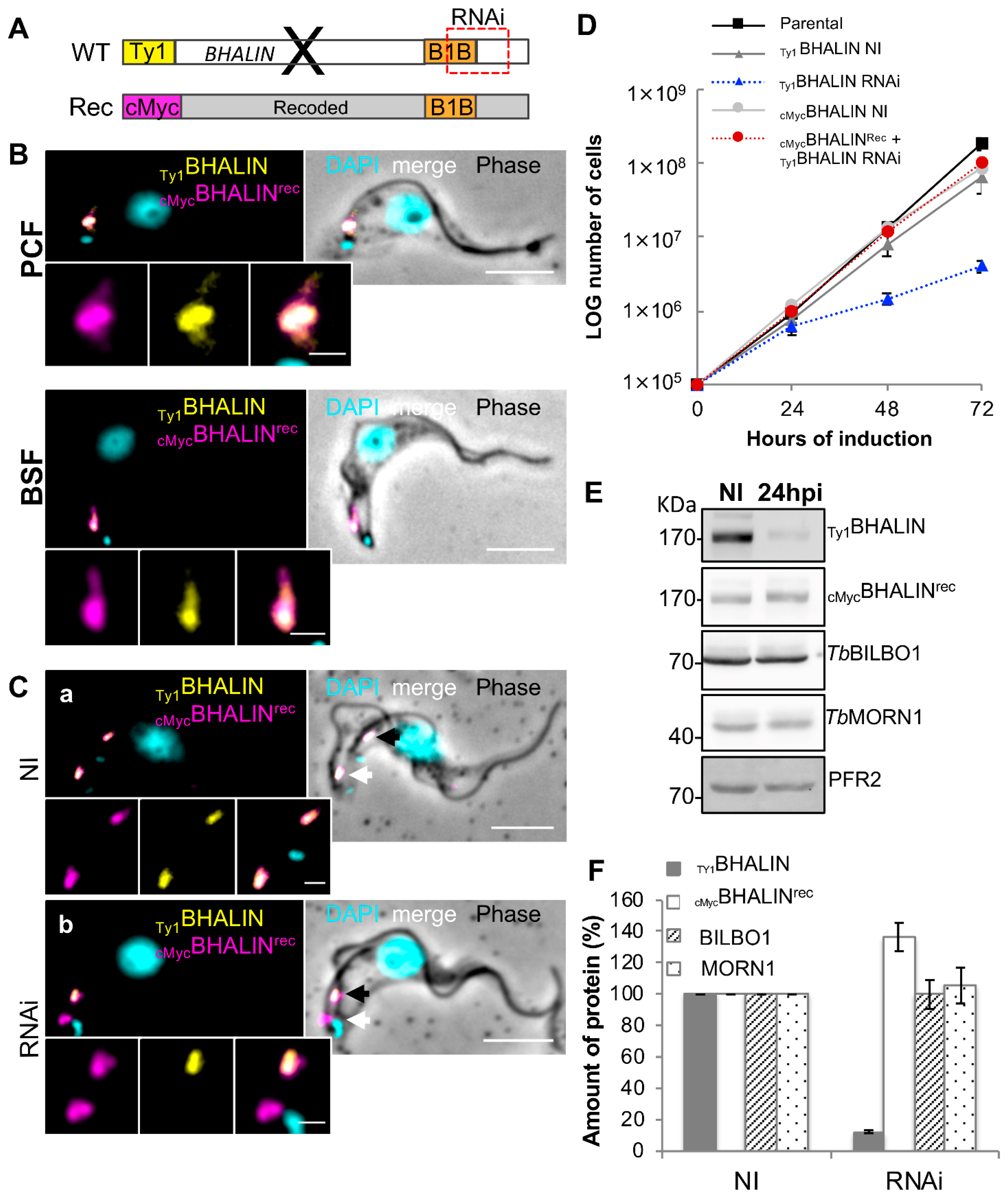
3.8. Expression of RNAi-Resistant BHALIN Rescues the Lethal Phenotype in BSF
3.9. Deletion of the BILBO1-Binding and C-Terminal Domains Relocates BHALIN to the Cytoplasm
3.10. The BHALIN TbBILBO1-Binding Domain (T2) Co-Localises with TbBILBO1 In Vivo
3.11. Expression of BHALIN BILBO1-Binding Domain in BSF Does Not Rescue the RNAi-Induced Lethal Phenotype
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nussbaum, K.; Honek, J.; Cadmus, C.M.C.V.C.; Efferth, T. Trypanosomatid Parasites Causing Neglected Diseases. Curr. Med. Chem. 2010, 17, 1594–1617. [Google Scholar] [CrossRef] [PubMed]
- Radwanska, M.; Vereecke, N.; Deleeuw, V.; Pinto, J.; Magez, S. Salivarian Trypanosomosis: A Review of Parasites Involved, Their Global Distribution and Their Interaction with the Innate and Adaptive Mammalian Host Immune System. Front. Immunol. 2018, 9, 2253. [Google Scholar] [CrossRef] [PubMed]
- Egeru, A.; Opio, J.; Siya, A.; Barasa, B.; Magaya, J.P.; Namaalwa, J.J. Tsetse Invasion as an Emerging Threat to Socioecological Resilience of Pastoral Communities in Karamoja, Uganda. Sustainability 2020, 12, 1599. [Google Scholar] [CrossRef] [Green Version]
- Grunfelder, C.G.; Engstler, M.; Weise, F.; Schwarz, H.; Stierhof, Y.D.; Morgan, G.W.; Field, M.C.; Overath, P. Endocytosis of a Glycosylphosphatidylinositol-Anchored Protein via Clathrin-Coated Vesicles, Sorting by Default in Endosomes, and Exocytosis via RAB11-Positive Carriers. Mol. Biol. Cell. 2003, 14, 2029–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.L.; Goulding, D.; Field, M.C. Clathrin-Mediated Endocytosis Is Essential in Trypanosoma brucei. EMBO J. 2003, 22, 4991–5002. [Google Scholar] [CrossRef] [Green Version]
- Overath, P.; Engstler, M. Endocytosis, Membrane Recycling and Sorting of GPI-Anchored Proteins: Trypanosoma brucei as a Model System. Mol. Microbiol. 2004, 53, 735–744. [Google Scholar] [CrossRef]
- Engstler, M.; Thilo, L.; Weise, F.; Grunfelder, C.G.; Schwarz, H.; Boshart, M.; Overath, P. Kinetics of Endocytosis and Recycling of the GPI-Anchored Variant Surface Glycoprotein in Trypanosoma brucei. J. Cell Sci. 2004, 117, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Gadelha, C.; Rothery, S.; Morphew, M.; McIntosh, J.R.; Severs, N.J.; Gull, K. Membrane Domains and Flagellar Pocket Boundaries Are Influenced by the Cytoskeleton in African Trypanosomes. Proc. Natl. Acad. Sci. USA 2009, 106, 17425–17430. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Salcedo, J.A.; Perez-Morga, D.; Gijon, P.; Dilbeck, V.; Pays, E.; Nolan, D.P. A Differential Role for Actin during the Life Cycle of Trypanosoma brucei. EMBO J. 2004, 23, 780–789. [Google Scholar] [CrossRef] [Green Version]
- Henley, G.L.; Lee, C.M.; Takeuchi, A. Electron Microscopy Observations on Trypanosoma brucei: Freeze-Cleaving and Thin-Sectioning Study of the Apical Part of the Flagellar Pocket. Z. Parasitenkd. 1978, 55, 181–187. [Google Scholar] [CrossRef]
- Bonhivers, M.; Nowacki, S.; Landrein, N.; Robinson, D.R. Biogenesis of the Trypanosome Endo-Exocytotic Organelle Is Cytoskeleton Mediated. PLoS Biol. 2008, 6, e105. [Google Scholar] [CrossRef] [PubMed]
- Lacomble, S.; Vaughan, S.; Gadelha, C.; Morphew, M.K.; Shaw, M.K.; McIntosh, J.R.; Gull, K. Three-Dimensional Cellular Architecture of the Flagellar Pocket and Associated Cytoskeleton in Trypanosomes Revealed by Electron Microscope Tomography. J. Cell Sci. 2009, 122, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Albisetti, A.; Florimond, C.; Landrein, N.; Vidilaseris, K.; Eggenspieler, M.; Lesigang, J.; Dong, G.; Robinson, D.R.; Bonhivers, M. Interaction between the Flagellar Pocket Collar and the Hook Complex via a Novel Microtubule-Binding Protein in Trypanosoma brucei. PLoS Pathog. 2017, 13, e1006710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florimond, C.; Sahin, A.; Vidilaseris, K.; Dong, G.; Landrein, N.; Dacheux, D.; Albisetti, A.; Byard, E.H.; Bonhivers, M.; Robinson, D.R. BILBO1 is a Scaffold Protein of the Flagellar Pocket Collar in the Pathogen Trypanosoma Brucei. PLoS Pathog. 2015, 11, e1004654. [Google Scholar] [CrossRef] [Green Version]
- Vidilaseris, K.; Shimanovskaya, E.; Esson, H.J.; Morriswood, B.; Dong, G. Assembly Mechanism of Trypanosoma brucei BILBO1, a Multidomain Cytoskeletal Protein. J. Biol. Chem. 2014, 289, 23870–23881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidilaseris, K.; Morriswood, B.; Kontaxis, G.; Dong, G. Structure of the TbBILBO1 Protein N-Terminal Domain from Trypanosoma brucei Reveals an Essential Requirement for a Conserved Surface Patch. J. Biol. Chem. 2014, 289, 3724–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.Y.; Pypaert, M.; Warren, G. Golgi Duplication in Trypanosoma brucei Requires Centrin2. Science 2005, 310, 1196–1198. [Google Scholar] [CrossRef]
- Shi, J.; Franklin, J.B.; Yelinek, J.T.; Ebersberger, I.; Warren, G.; He, C.Y. Centrin4 Coordinates Cell and Nuclear Division in T. brucei. J. Cell Sci. 2008, 121, 3062–3070. [Google Scholar] [CrossRef] [Green Version]
- Morriswood, B.; Havlicek, K.; Demmel, L.; Yavuz, S.; Sealey-Cardona, M.; Vidilaseris, K.; Anrather, D.; Kostan, J.; Djinović-Carugo, K.; Roux, K.J.; et al. Novel Bilobe Components in Trypanosoma Brucei Identified Using Proximity-Dependent Biotinylation. Eukaryot. Cell 2013, 12, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Pham, K.T.M.; Hu, H.; Li, Z. Maintenance of Hook Complex Integrity and Centrin Arm Assembly Facilitates Flagellum Inheritance in Trypanosoma brucei. J. Biol. Chem. 2020, 295, 12962–12974. [Google Scholar] [CrossRef]
- Perry, J.A.; Sinclair-Davis, A.N.; McAllaster, M.R.; de Graffenried, C.L. TbSmee1 Regulates Hook Complex Morphology and the Rate of Flagellar Pocket Uptake in Trypanosoma brucei. Mol. Microbiol. 2018, 107, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Gheiratmand, L.; Chen, Y.; Lim, T.K.; Zhang, J.; Li, S.; Xia, N.; Liu, B.; Lin, Q.; He, C.Y. A Comparative Proteomic Analysis Reveals a New Bi-Lobe Protein Required for Bi-Lobe Duplication and Cell Division in Trypanosoma brucei. PLoS ONE 2010, 5, e9660. [Google Scholar] [CrossRef] [Green Version]
- Morriswood, B.; He, C.Y.; Sealey-Cardona, M.; Yelinek, J.; Pypaert, M.; Warren, G. The Bilobe Structure of Trypanosoma brucei Contains a MORN-Repeat Protein. Mol. Biochem. Parasitol. 2009, 167, 95–103. [Google Scholar] [CrossRef] [PubMed]
- de Graffenried, C.L.; Ho, H.H.; Warren, G. Polo-like Kinase Is Required for Golgi and Bilobe Biogenesis in Trypanosoma brucei. J. Cell Biol. 2008, 181, 431–438. [Google Scholar] [CrossRef]
- Morriswood, B.; Schmidt, K. A MORN Repeat Protein Facilitates Protein Entry into the Flagellar Pocket of Trypanosoma brucei. Eukaryot. Cell 2015, 14, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Pham, K.T.M.; Zhou, Q.; Kurasawa, Y.; Li, Z. BOH1 Cooperates with Polo-like Kinase to Regulate Flagellum Inheritance and Cytokinesis Initiation in Trypanosoma brucei. J. Cell Sci. 2019, 132, jcs230581. [Google Scholar] [CrossRef] [Green Version]
- Poon, S.K.; Peacock, L.; Gibson, W.; Gull, K.; Kelly, S. A Modular and Optimized Single Marker System for Generating Trypanosoma brucei Cell Lines Expressing T7 RNA Polymerase and the Tetracycline Repressor. Open Biol. 2012, 2, 110037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibu, V.P.; Storm, L.; Haile, S.; Clayton, C.; Horn, D. A Doubly Inducible System for RNA Interference and Rapid RNAi Plasmid Construction in Trypanosoma brucei. Mol. Biochem. Parasitol. 2005, 139, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Dean, S.; Sunter, J.; Wheeler, R.J.; Hodkinson, I.; Gluenz, E.; Gull, K. A Toolkit Enabling Efficient, Scalable and Reproducible Gene Tagging in Trypanosomatids. Open Biol. 2015, 5, 140197. [Google Scholar] [CrossRef] [Green Version]
- Hirumi, H.; Hirumi, K. Continuous Cultivation of Trypanosoma brucei Blood Stream Forms in a Medium Containing a Low Concentration of Serum Protein without Feeder Cell Layers. J. Parasitol. 1989, 75, 985–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirtz, E.; Leal, S.; Ochatt, C.; Cross, G.A.M. A Tightly Regulated Inducible Expression System for Conditional Gene Knock-Outs and Dominant-Negative Genetics in Trypanosoma Brucei. Mol. Biochem. Parasitol. 1999, 99, 89–101. [Google Scholar] [CrossRef]
- Schumann Burkard, G.; Jutzi, P.; Roditi, I. Genome-Wide RNAi Screens in Bloodstream Form Trypanosomes Identify Drug Transporters. Mol. Biochem. Parasitol. 2011, 175, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H.; Johnsson, A.; Wennergren, S.; Wernstedt, C.; Betsholtz, C.; Westermark, B. A Human Osteosarcoma Cell Line Secretes a Growth Factor Structurally Related to a Homodimer of PDGF A-Chains. Nature 1986, 319, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Bastin, P.; Bagherzadeh, A.; Matthews, K.R.; Gull, K. A Novel Epitope Tag System to Study Protein Targeting and Organelle Biogenesis in Trypanosoma brucei. Mol. Biochem. Parasitol. 1996, 77, 235–239. [Google Scholar] [CrossRef]
- Chan, K.Y.; Matthews, K.R.; Ersfeld, K. Functional Characterisation and Drug Target Validation of a Mitotic Kinesin-13 in Trypanosoma brucei. PLoS Pathog. 2010, 6, e1001050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bringaud, F.; Robinson, D.R.; Barradeau, S.; Biteau, N.; Baltz, D.; Baltz, T. Characterization and Disruption of a New Trypanosoma brucei Repetitive Flagellum Protein, Using Double-Stranded RNA Inhibition. Mol. Biochem. Parasitol. 2000, 111, 283–297. [Google Scholar] [CrossRef]
- Pradel, L.C. NIMA-Related Kinase TbNRKC Is Involved in Basal Body Separation in Trypanosoma brucei. J. Cell Sci. 2006, 119, 1852–1863. [Google Scholar] [CrossRef] [Green Version]
- Beyer, H.M.; Gonschorek, P.; Samodelov, S.L.; Meier, M.; Weber, W.; Zurbriggen, M.D. AQUA Cloning: A Versatile and Simple Enzyme-Free Cloning Approach. PLoS ONE 2015, 10, e0137652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacheux, D.; Landrein, N.; Thonnus, M.; Gilbert, G.; Sahin, A.; Wodrich, H.; Robinson, D.R.; Bonhivers, M. A MAP6-Related Protein Is Present in Protozoa and Is Involved in Flagellum Motility. PLoS ONE 2012, 7, e31344. [Google Scholar] [CrossRef] [Green Version]
- Hannaert, V.; Albert, M.-A.; Rigden, D.J.; da Silva Giotto, M.T.; Thiemann, O.; Garratt, R.C.; Van Roy, J.; Opperdoes, F.R.; Michels, P.A.M. Kinetic Characterization, Structure Modelling Studies and Crystallization of Trypanosoma brucei Enolase. Eur. J. Biochem. 2003, 270, 3205–3213. [Google Scholar] [CrossRef]
- Woods, A.; Sherwin, T.; Sasse, R.; MacRae, T.H.; Baines, A.J.; Gull, K. Definition of Individual Components within the Cytoskeieton of Trypanosoma brucei by a Library of Monoclonal Antibodies. J. Cell Sci. 1989, 93, 491–500. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Halliday, C.; de Castro-Neto, A.; Alcantara, C.L.; Cunha-E.-Silva, N.L.; Vaughan, S.; Sunter, J.D. Trypanosomatid Flagellar Pocket from Structure to Function. Trends Parasitol. 2021, 37, 317–329. [Google Scholar] [CrossRef]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The Genome of the African Trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.Fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Audic, S.; Claverie, J.-M.; Blanc, G. BLAST-EXPLORER Helps You Building Datasets for Phylogenetic Analysis. BMC Evol. Biol. 2010, 10, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davaasuren, B.; Yamagishi, J.; Mizushima, D.; Narantsatsral, S.; Otgonsuren, D.; Myagmarsuren, P.; Battsetseg, B.; Battur, B.; Inoue, N.; Suganuma, K. Draft Genome Sequence of Trypanosoma equiperdum Strain IVM-T1. Microbiol. Resour. Announc. 2019, 8, e01119-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrenfeltz, S.; Basenko, E.Y.; Crouch, K.; Harb, O.S.; Kissinger, J.C.; Roos, D.S.; Shanmugasundram, A.; Silva-Franco, F. EuPathDB: The Eukaryotic Pathogen Genomics Database Resource. Methods Mol. Biol. 2018, 1757, 69–113. [Google Scholar] [CrossRef] [Green Version]
- Kohl, L.; Sherwin, T.; Gull, K. Assembly of the Paraflagellar Rod and the Flagellum Attachment Zone Complex During the Trypanosoma brucei Cell Cycle. J. Eukaryot. Microbiol. 1999, 46, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Ooi, C.; Bastin, P. More than Meets the Eye: Understanding Trypanosoma brucei Morphology in the Tsetse. Front. Cell. Infect. Microbiol. 2013, 3, 71. [Google Scholar] [CrossRef]
- Armstrong, J.K.; Wenby, R.B.; Meiselman, H.J.; Fisher, T.C. The Hydrodynamic Radii of Macromolecules and Their Effect on Red Blood Cell Aggregation. Biophys. J. 2004, 87, 4259–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brickman, M.J.; Cook, J.M.; Balber, A.E. Low Temperature Inhibits Transport from Tubular Endosomes to a Perinuclear, Acidic Compartment in African Trypanosomes. J. Cell Sci. 1995, 108, 3611–3621. [Google Scholar] [CrossRef] [PubMed]
- Sinclair-Davis, A.N.; McAllaster, M.R.; de Graffenried, C.L. A Functional Analysis of TOEFAZ1 Uncovers Protein Domains Essential for Cytokinesis in Trypanosoma brucei. J. Cell Sci. 2017, 130, 3918–3932. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, D.; Bonhivers, M.; Robinson, D. The Trypanosome Flagellar Pocket Collar and Its Ring Forming Protein—TbBILBO1. Cells 2016, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Isch, C.; Majneri, P.; Landrein, N.; Pivovarova, Y.; Lesigang, J.; Lauruol, F.; Robinson, D.R.; Dong, G.; Bonhivers, M. Structural and Functional Studies of the First Tripartite Protein Complex at the Trypanosoma brucei Flagellar Pocket Collar. PLoS Pathog. 2021, 17, e1009329. [Google Scholar] [CrossRef]
- Broadhead, R.; Dawe, H.R.; Farr, H.; Griffiths, S.; Hart, S.R.; Portman, N.; Shaw, M.K.; Ginger, M.L.; Gaskell, S.J.; McKean, P.G.; et al. Flagellar Motility Is Required for the Viability of the Bloodstream Trypanosome. Nature 2006, 440, 224–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmel, L.; Schmidt, K.; Lucast, L.; Havlicek, K.; Zankel, A.; Koestler, T.; Reithofer, V.; de Camilli, P.; Warren, G. The Endocytic Activity of the Flagellar Pocket in Trypanosoma brucei Is Regulated by an Adjacent Phosphatidylinositol Phosphate Kinase. J. Cell Sci. 2016, 129, 2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, P.T.; Field, M.C. Phosphoinositides, Kinases and Adaptors Coordinating Endocytosis in Trypanosoma brucei. Commun. Integr. Biol. 2015, 8, e1082691. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broster Reix, C.E.; Florimond, C.; Cayrel, A.; Mailhé, A.; Agnero-Rigot, C.; Landrein, N.; Dacheux, D.; Havlicek, K.; Bonhivers, M.; Morriswood, B.; et al. Bhalin, an Essential Cytoskeleton-Associated Protein of Trypanosoma brucei Linking TbBILBO1 of the Flagellar Pocket Collar with the Hook Complex. Microorganisms 2021, 9, 2334. https://doi.org/10.3390/microorganisms9112334
Broster Reix CE, Florimond C, Cayrel A, Mailhé A, Agnero-Rigot C, Landrein N, Dacheux D, Havlicek K, Bonhivers M, Morriswood B, et al. Bhalin, an Essential Cytoskeleton-Associated Protein of Trypanosoma brucei Linking TbBILBO1 of the Flagellar Pocket Collar with the Hook Complex. Microorganisms. 2021; 9(11):2334. https://doi.org/10.3390/microorganisms9112334
Chicago/Turabian StyleBroster Reix, Christine E., Célia Florimond, Anne Cayrel, Amélie Mailhé, Corentin Agnero-Rigot, Nicolas Landrein, Denis Dacheux, Katharina Havlicek, Mélanie Bonhivers, Brooke Morriswood, and et al. 2021. "Bhalin, an Essential Cytoskeleton-Associated Protein of Trypanosoma brucei Linking TbBILBO1 of the Flagellar Pocket Collar with the Hook Complex" Microorganisms 9, no. 11: 2334. https://doi.org/10.3390/microorganisms9112334