
Molecular Epidemiology of Multidrug-Resistant Uropathogenic Escherichia coli O25b Strains Associated with Complicated Urinary Tract Infection in Children

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Strains of UPEC
2.2. Typing of UPEC O25b Clinical Strains
2.3. Antimicrobial Susceptibility Profile of UPEC O25b Strains
2.4. Phenotypic Determination of ESBL and MBL Production in Clinical Strains of UPEC O25b
2.5. Typing of Phylogenetic Groups, Virulence Genes, and Integrons Associated with UPEC O25b Strains
2.6. Pulsed-Field Gel Electrophoresis (PFGE) of Clinical UPEC O25b Strains
2.7. Typing of UPEC O25b Clinical Strains Using MLST
2.8. Statistical Analysis
3. Results
3.1. UPEC O25b Strains Associated with cUTIs
3.2. Multidrug Resistance Profile of UPEC O25b Clinical Strains
3.3. Resistance Associated with the Phenotypic Expression of ESBLs and Integrons
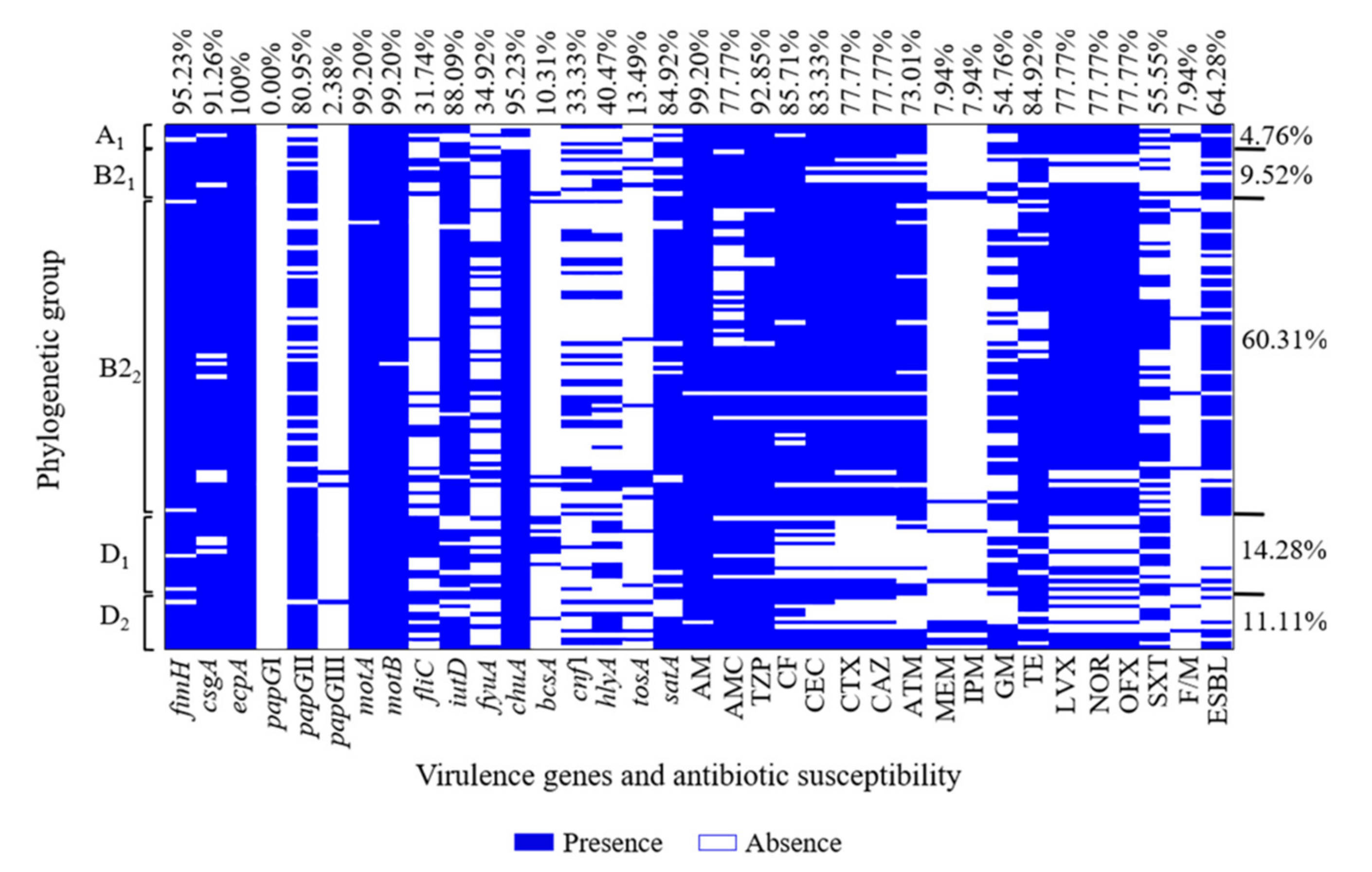
3.4. Virulence Gene and Phylogenetic Group Associations of UPEC O25b Clinical Strains
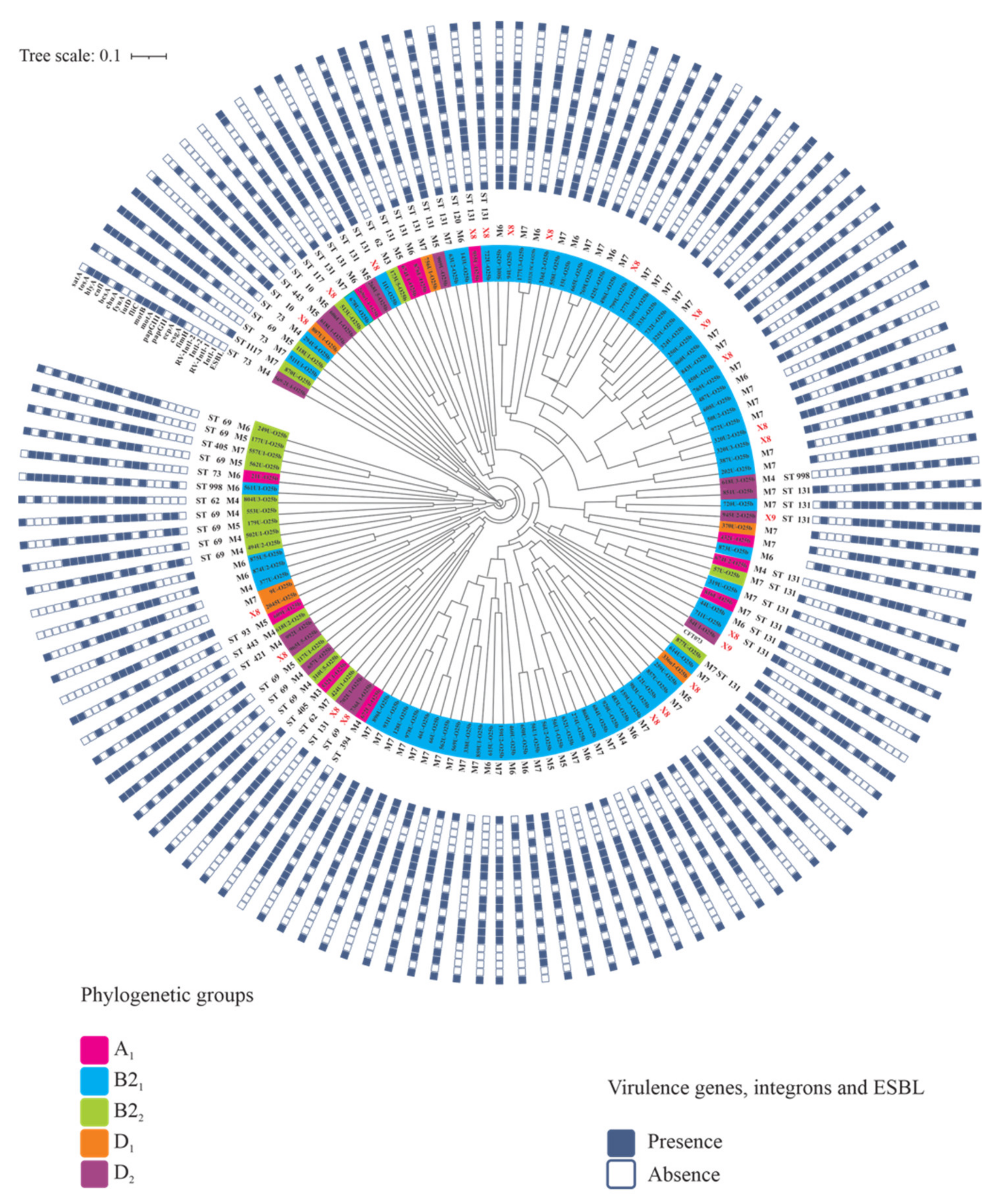
3.5. PFGE Genetic Diversity of UPEC O25b Strains
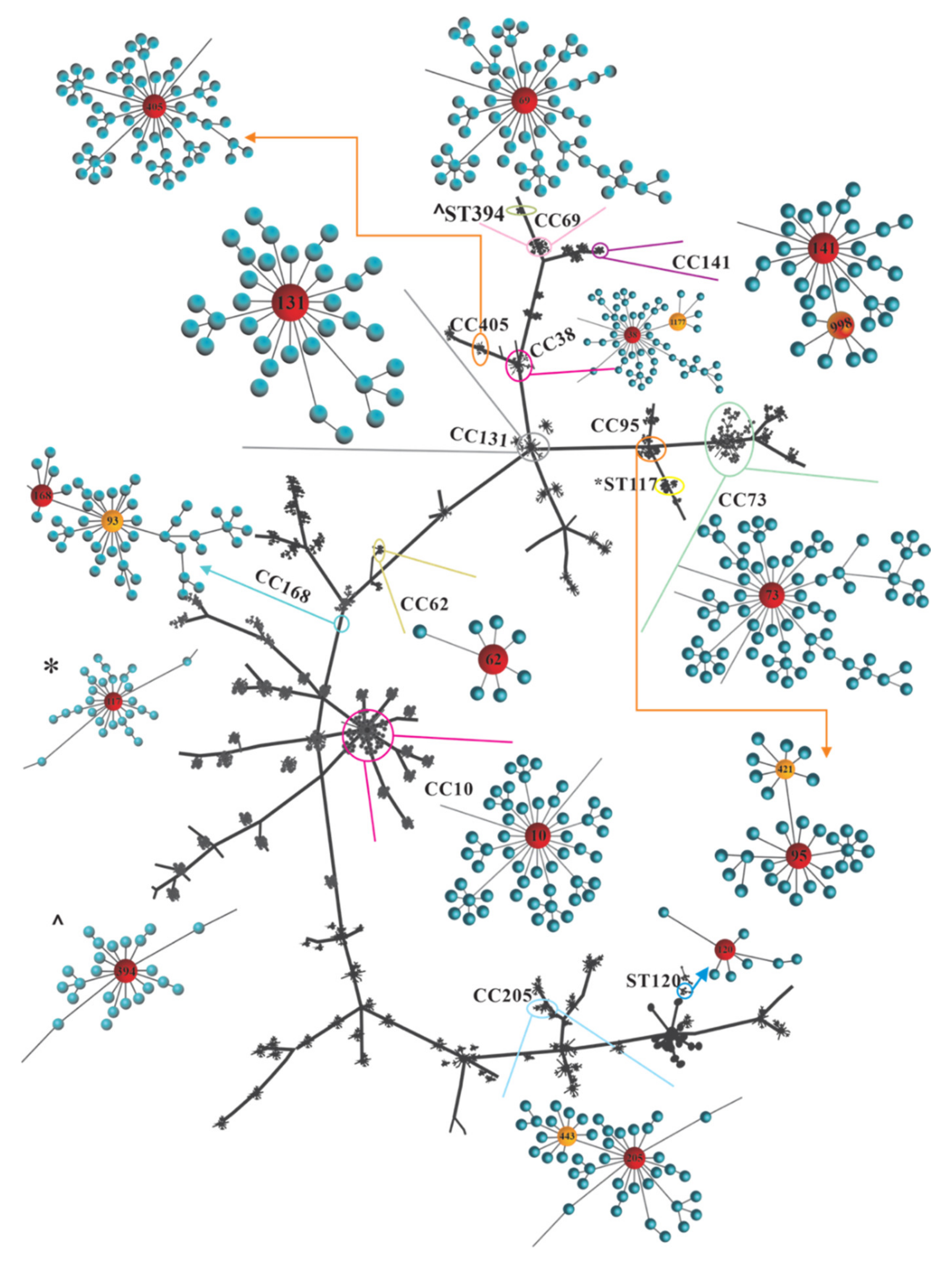
3.6. STs and CCs Maintain an Association with UPEC O25b Clinical Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
| AM | Ampicillin. |
| AMC | Amoxicillin–clavulanate. |
| ATM | Aztreonam. |
| CAZ | Ceftazidime. |
| CCs | Clonal complexes. |
| CEC | Cefaclor. |
| CF | Cephalothin. |
| CLSI | Clinical and Laboratory Standard Institute. |
| CRO | Ceftriaxone. |
| CTX-M | Cefotaximase-M. |
| CTX | Cefotaxime. |
| cUTIs | Complicated urinary tract infections. |
| ESBLs | Extended-spectrum β-lactamases. |
| F/M | Nitrofurantoin. |
| GM | Gentamicin. |
| IPM | Imipenem. |
| LVX | Levofloxacin. |
| MBLs | Metallo-β-lactamases. |
| MDR | Multidrug-resistant. |
| MEM | Meropenem. |
| MLST | Multilocus Sequence Typing. |
| NOR | Norfloxacin. |
| OFX | Ofloxacin. |
| PFGE | Pulsed-field gel Electrophoresis. |
| PLSD | ANOVA Fisher’s protected least significant difference. |
| SNP | Single-nucleotide polymorphism. |
| STs | Sequence types. |
| SXT | Trimethoprim–sulfamethoxazole. |
| TE | Tetracycline. |
| TZP | Piperacillin–tazobactam. |
| UPEC | Uropathogenic Escherichia coli. |
| UPGMA | Unweighted Pair-Cluster Method using Arithmetic Averages. |
| UTIs | Urinary tract infections. |
| XDR | Extremely drug-resistant. |
References
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Luna-Pineda, V.; Ochoa, S.; Cruz Córdova, A.; Cázares, V.; Vélez, F.; Hernández-Castro, R.; Xicohtencatl-Cortes, J. Infecciones del tracto urinario, inmunidad y vacunación. Bol. Méd. Hosp. Infant. Méx. 2018, 75, 67–78. [Google Scholar] [CrossRef]
- Li, D.; Liu, B.; Chen, M.; Guo, D.; Guo, X.; Liu, F.; Feng, L.; Wang, L. A multiplex PCR method to detect 14 Escherichia coli serogroups associated with urinary tract infections. J. Microbiol. Methods 2010, 82, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Momtaz, H.; Karimian, A.; Madani, M.; Dehkordi, F.S.; Ranjbar, R.; Sarshar, M.; Souod, N. Uropathogenic Escherichia coli in Iran: Serogroup distributions, virulence factors and antimicrobial resistance properties. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Rodríguez-Moctezuma, J.R.; Domínguez-Trejo, P.; Vaca-Paniagua, F.; Vaca, S. Virulence factors, antibiotic resistance phenotypes and O-serogroups of Escherichia coli strains isolated from community-acquired urinary tract infection patients in Mexico. J. Microbiol. Immunol. Infect. 2017, 50, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Oskouie, A.N.; Hasani, A.; Rezaee, M.A.; Soroush Bar Haghi, M.H.; Hasani, A.; Soltani, E. A relationship between O-serotype, antibiotic susceptibility and biofilm formation in uropathogenic Escherichia coli. Microb. Drug Resist. 2019, 25, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Prakapaite, R.; Saab, F.; Planciuniene, R.; Petraitis, V.; Walsh, T.J.; Petraitiene, R.; Semoskaite, R.; Baneviciene, R.; Kalediene, L.; Kavaliauskas, P. Molecular characterization of uropathogenic Escherichia coli reveals emergence of drug resistant O15, O22 and O25 serogroups. Medicina 2019, 55, 733. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Chiñas, U.; Pérez-Ramos, A.; Belmont-Monroy, L.; Chávez-Berrocal, M.E.; González-Villalobos, E.; Navarro-Ocaña, A.; Eslava, C.A.; Molina-Lopez, J. Characterization of auto-agglutinating and non-typeable uropathogenic Escherichia coli strains. J. Infect. Dev. Ctries. 2019, 13, 465–472. [Google Scholar] [CrossRef]
- Mostafavi, S.K.S.; Najar-Peerayeh, S.; Mobarez, A.M.; Parizi, M.K. Serogroup distribution, diversity of exotoxin gene profiles, and phylogenetic grouping of CTX-M-1-producing uropathogenic Escherichia coli. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 148–153. [Google Scholar] [CrossRef]
- Nielsen, K.L.; Stegger, M.; Kiil, K.; Godfrey, P.A.; Feldgarden, M.; Lilje, B.; Andersen, P.S.; Frimodt-Møller, N. Whole-genome comparison of urinary pathogenic Escherichia coli and faecal isolates of UTI patients and healthy controls. Int. J. Med. Microbiol. 2017, 307, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.; Meng, T.; Guastalli, E.A.; Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; De Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef] [PubMed]
- Braga, J.F.V.; Chanteloup, N.K.; Trotereau, A.; Baucheron, S.; Guabiraba, R.; Ecco, R.; Schouler, C. Diversity of Escherichia coli strains involved in vertebral osteomyelitis and arthritis in broilers in Brazil. BMC Vet. Res. 2016, 12, 140. [Google Scholar] [CrossRef]
- Bozcal, E.; Eldem, V.; Aydemir, S.; Skurnik, M. The relationship between phylogenetic classification, virulence and antibiotic resistance of extraintestinal pathogenic Escherichia coli in İzmir province, Turkey. PeerJ 2018, 6, e5470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas-Chanoine, M.H.; Blanco, J.; Leflon-Guibout, V.; Demarty, R.; Alonso, M.P.; Caniça, M.M.; Park, Y.J.; Lavigne, J.P.; Pitout, J.; Johnson, J.R. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 2008, 61, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa, S.A.; Cruz-Córdova, A.; Luna-Pineda, V.M.; Reyes-Grajeda, J.P.; Cázares-Domínguez, V.; Escalona, G.; Sepúlveda-González, M.E.; López-Montiel, F.; Arellano-Galindo, J.; López-Martínez, B.; et al. Multidrug- and extensively drug-resistant uropathogenic Escherichia coli clinical strains: Phylogenetic groups widely associated with integrons maintain high genetic diversity. Front. Microbiol. 2016, 7, 2042. [Google Scholar] [CrossRef] [Green Version]
- Luna-Pineda, V.M.; Ochoa, S.A.; Cruz-Córdova, A.; Cázares-Domínguez, V.; Reyes-Grajeda, J.P.; Flores-Oropeza, M.A.; Arellano-Galindo, J.; Hernández-Castro, R.; Flores-Encarnación, M.; Ramírez-Vargas, A.; et al. Correction: Features of urinary Escherichia coli isolated from children with complicated and uncomplicated urinary tract infections in Mexico. PLoS ONE 2018, 13, e0208285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppé, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa, S.A.; Cruz-Córdova, A.; Rodea, G.E.; Cázares-Domínguez, V.; Escalona, G.; Arellano-Galindo, J.; Hernández-Castro, R.; Reyes-López, A.; Xicohtencatl-Cortes, J. Phenotypic characterization of multidrug-resistant Pseudomonas aeruginosa strains isolated from pediatric patients associated to biofilm formation. Microbiol. Res. 2015, 172, 68–78. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Mancilla-Rojano, J.; Ochoa, S.A.; Reyes-Grajeda, J.P.; Flores, V.; Medina-Contreras, O.; Espinosa-Mazariego, K.; Parra-Ortega, I.; Rosa-Zamboni, D.D.L.; Castellanos-Cruz, M.D.C.; Arellano-Galindo, J.; et al. Molecular epidemiology of acinetobacter calcoaceticus-acinetobacter baumannii complex isolated from children at the hospital Infantil de México Federico Gómez. Front. Microbiol. 2020, 11, 576673. [Google Scholar] [CrossRef] [PubMed]
- Landau, S.; Rabe-Hesketh, S. StatView for Windows, Version 5.0. Stat. Methods Med. Res. 1999, 8, 337–341. [Google Scholar] [CrossRef]
- Feil, E.J.; Li, B.C.; Aanensen, D.M.; Hanage, W.P.; Spratt, B.G. eBURST: Inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J. Bacteriol. 2004, 186, 1518–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyna-Flores, F.; Barrios, H.; Garza-Ramos, U.; Sánchez-Pérez, A.; Rojas-Moreno, T.; Uribe-Salas, F.J.; Fagundo-Sierra, R.; Silva-Sanchez, J. Molecular epidemiology of Escherichia coli O25b-ST131 isolates causing community-acquired UTIs in Mexico. Diagn. Microbiol. Infect. Dis. 2013, 76, 396–398. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Díaz-Velásquez, C.E.; Uribe-García, A.; Labastida, A.; Peñaloza-Figueroa, F.; Domínguez-Trejo, P.; García, L.R.; Vaca-Paniagua, F.; Vaca, S. Whole-genome sequence analysis of multidrug-resistant uropathogenic strains of Escherichia coli from Mexico. Infect. Drug Resist. 2019, 12, 2363–2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashti, A.A.; Vali, L.; El-Shazly, S.; Jadaon, M.M. The characterization and antibiotic resistance profiles of clinical Escherichia coli O25b-B2-ST131 isolates in Kuwait. BMC Microbiol. 2014, 14, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlieghe, E.R.; Huang, T.D.; Phe, T.; Bogaerts, P.; Berhin, C.; De Smet, B.; Peetermans, W.E.; Jacobs, J.A.; Glupczynski, Y. Prevalence and distribution of beta-lactamase coding genes in third-generation cephalosporin-resistant Enterobacteriaceae from bloodstream infections in Cambodia. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1223–1229. [Google Scholar] [CrossRef]
- El-Badawy, M.F.; Tawakol, W.M.; Maghrabi, I.A.; Mansy, M.S.; Shohayeb, M.M.; Ashour, M.S. Iodometric and molecular detection of ESBL production among clinical isolates of E. coli fingerprinted by ERIC-PCR: The first egyptian report declares the emergence of E. coli O25b-ST131clone Harboring bla(GES). Microb. Drug Resist. 2017, 23, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Demirci, M.; Ünlü, Ö.; Tosun, A.I. Detection of O25b-ST131 clone, CTX-M-1 and CTX-M-15 genes via real-time PCR in Escherichia coli strains in patients with UTIs obtained from a university hospital in Istanbul. J. Infect. Public Health 2019, 12, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.F.; Chen, W.L.; Hung, W.Y.; Huang, I.F.; Chiou, Y.H.; Chen, Y.S.; Lee, S.S.; Hung, C.H.; Wang, J.L. Emergence of extended spectrum-β-lactamase-producing Escherichia coli O25b-ST131: A major community-acquired uropathogen in infants. Pediatr. Infect. Dis. J. 2015, 34, 469–475. [Google Scholar] [CrossRef]
- Aquino-Andrade, A.; Merida-Vieyra, J.; Garza, E.A.D.; Arzate-Barbosa, P.; De Colsa Ranero, A. Carbapenemase-producing Enterobacteriaceae in Mexico: Report of seven non-clonal cases in a pediatric hospital. BMC Microbiol. 2018, 18, 38. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Romero, A.L.; Silva-Sanchez, J.; Garza-Ramos, U.; Barrios, H.; Sánchez-Pérez, A.; Reyna-Flores, F. Molecular characterization of ESBL-producing Escherichia coli isolates from hospital- and community-acquired infections in NW Mexico. Diagn. Microbiol. Infect. Dis. 2017, 87, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Demir, M.; Kazanasmaz, H. Uropathogens and antibiotic resistance in the community and hospital-induced urinary tract infected children. J. Glob. Antimicrob. Resist. 2020, 20, 68–73. [Google Scholar] [CrossRef]
- Kara, A.; Gurgoze, M.K. The use of nitrofurantoin for children with acute cystitis caused by extended-spectrum Β-lactamase-producing Escherichia coli. J. Pediatr. Urol. 2019, 15, 378.e1–378.e5. [Google Scholar] [CrossRef]
- Vazouras, K.; Velali, K.; Tassiou, I.; Anastasiou-Katsiardani, A.; Athanasopoulou, K.; Barbouni, A.; Jackson, C.; Folgori, L.; Zaoutis, T.; Basmaci, R.; et al. Antibiotic treatment and antimicrobial resistance in children with urinary tract infections. J. Glob. Antimicrob. Resist. 2020, 20, 4–10. [Google Scholar] [CrossRef]
- World Health Organization. WHO Model List of Essential Medicines for Children: 6th List; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Grabe, M.; Bartoletti, R.; Johansen, T.E.B.; Cai, T.; Çek, M.; Köves, B.; Naber, K.G.; Pickard, R.S.; Tenke, P.; Wagenlehner, F.; et al. Guidelines on Urological Infections; EAU: Arnhem, The Netherlands, 2015. [Google Scholar]
- Farra, A.; Frank, T.; Tondeur, L.; Bata, P.; Gody, J.C.; Onambele, M.; Rafaï, C.; Vray, M.; Breurec, S. High rate of faecal carriage of extended-spectrum β-lactamase-producing Enterobacteriaceae in healthy children in Bangui, Central African Republic. Clin. Microbiol. Infect. 2016, 22, 891.e1–891.e4. [Google Scholar] [CrossRef] [Green Version]
- Huang, I.F.; Lee, W.Y.; Wang, J.L.; Hung, C.H.; Hu, H.H.; Hung, W.Y.; Hung, Y.J.; Chen, W.C.; Shen, Y.T.; Cheng, M.F. Fecal carriage of multidrug-resistant Escherichia coli by community children in southern Taiwan. BMC Gastroenterol. 2018, 18, 86. [Google Scholar] [CrossRef]
- Flament-Simon, S.C.; Nicolas-Chanoine, M.H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal structure, virulence factor-encoding genes and antibiotic resistance of Escherichia coli, causing urinary tract infections and other extraintestinal infections in humans in Spain and France during 2016. Antibiotics 2020, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- Karami, N.; Wold, A.E.; Adlerberth, I. Antibiotic resistance is linked to carriage of papC and iutA virulence genes and phylogenetic group D background in commensal and uropathogenic Escherichia coli from infants and young children. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 721–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Guranie, D.; Al-Mayahie, S. Prevalence of E. coli ST131 among Uropathogenic E. coli Isolates from Iraqi Patients in Wasit Province, Iraq. Int. J. Microbiol. 2020, 2020, 8840561. [Google Scholar] [CrossRef]
- Dziri, O.; Dziri, R.; Maraoub, A.; Chouchani, C. Characterization of O25b-ST131 Escherichia coli clone producing CTX-M-15, DHA-4, and CMY-42 in urinary tract infections in a Tunisian Island. Microb. Drug Resist. 2020, 26, 741–746. [Google Scholar] [CrossRef]
- Lane, M.C.; Alteri, C.J.; Smith, S.N.; Mobley, H.L. Expression of flagella is coincident with uropathogenic Escherichia coli ascension to the upper urinary tract. Proc. Natl. Acad. Sci. USA 2007, 104, 16669–16674. [Google Scholar] [CrossRef] [Green Version]
- Engstrom, M.D.; Alteri, C.J.; Mobley, H.L. A conserved PapB family member, TosR, regulates expression of the uropathogenic Escherichia coli RTX nonfimbrial adhesin TosA while conserved LuxR family members TosE and TosF suppress motility. Infect. Immun. 2014, 82, 3644–3656. [Google Scholar] [CrossRef] [Green Version]
- Vigil, P.D.; Stapleton, A.E.; Johnson, J.R.; Hooton, T.M.; Hodges, A.P.; He, Y.; Mobley, H.L. Presence of putative repeat-in-toxin gene tosA in Escherichia coli predicts successful colonization of the urinary tract. MBio 2011, 2, e00066-11. [Google Scholar] [CrossRef] [Green Version]
- Xicohtencatl-Cortes, J.; Cruz-Córdova, A.; Cázares-Domínguez, V.; Escalona-Venegas, G.; Zavala-Vega, S.; Arellano-Galindo, J.; Romo-Castillo, M.; Hernández-Castro, R.; Ochoa, S.A.; Luna-Pineda, V.M. Uropathogenic Escherichia coli strains harboring tosA gene were associated to high virulence genes and a multidrug-resistant profile. Microb. Pathog. 2019, 134, 103593. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, M.B.; Zamora, E.; Diaz, P.; Tollefson, L.; Fedorka-Cray, P.J.; Headrick, M.L. Risk factors for fecal quinolone-resistant Escherichia coli in Mexican children. Antimicrob. Agents Chemother. 2003, 47, 1999–2001. [Google Scholar] [CrossRef] [Green Version]
- Dyar, O.J.; Hoa, N.Q.; Trung, N.V.; Phuc, H.D.; Larsson, M.; Chuc, N.T.; Lundborg, C.S. High prevalence of antibiotic resistance in commensal Escherichia coli among children in rural Vietnam. BMC Infect. Dis. 2012, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, V.; Leflon-Guibout, V.; Blanco, J.; Haenni, M.; Madec, J.Y.; Rafignon, G.; Bruno, P.; Mora, A.; Lopez, C.; Dahbi, G.; et al. Prevalence of day-care centre children (France) with faecal CTX-M-producing Escherichia coli comprising O25b:H4 and O16:H5 ST131 strains. J. Antimicrob. Chemother. 2014, 69, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Guo, S.; Seow, K.L.G.; Ming, G.O.H.; Schlundt, J. Characterization of extended-spectrum beta-lactamase-producing Escherichia coli isolates from Jurong Lake, Singapore with whole-genome-sequencing. Int. J. Environ. Res. Public Health 2021, 18, 937. [Google Scholar] [CrossRef] [PubMed]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Blanco, J.; Mora, A.; Mamani, R.; López, C.; Blanco, M.; Dahbi, G.; Herrera, A.; Blanco, J.E.; Alonso, M.P.; García-Garrote, F.; et al. National survey of Escherichia coli causing extraintestinal infections reveals the spread of drug-resistant clonal groups O25b:H4-B2-ST131, O15:H1-D-ST393 and CGA-D-ST69 with high virulence gene content in Spain. J. Antimicrob. Chemother. 2011, 66, 2011–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trong, L.N.; Viet, H.D.; Quoc, D.T.; Tuan, L.A.; Raal, A.; Usai, D.; Madeddu, S.; Marchetti, M.; Usai, M.; Rappelli, P.; et al. In Vitro Antimicrobial Activity of Essential Oil Extracted from Leaves of Leoheo domatiophorus Chaowasku, D.T. Ngo and H.T. Le in Vietnam. Plants 2020, 9, 453. [Google Scholar] [CrossRef] [Green Version]
- Trong, L.N.; Viet, H.D.; Quoc, D.T.; Tuan, L.A.; Raal, A.; Usai, D.; Sanna, G.; Carta, A.; Rappelli, P.; Diaz, N.; et al. Biological Activities of Essential Oils from Leaves of Paramignya trimera (Oliv.) Guillaum and Limnocitrus littoralis (Miq.) Swingle. Antibiotics 2020, 9, 207. [Google Scholar] [CrossRef] [PubMed]
- Donadu, M.G.; Trong, L.N.; Viet, H.D.; Quoc, D.T.; Tuan, L.A.; Raal, A.; Usai, M.; Marchetti, M.; Sanna, G.; Madeddu, S.; et al. Phytochemical Compositions and Biological Activities of Essential Oils from the Leaves, Rhizomes and Whole Plant of Hornstedtia bella Škorničk. Antibiotics 2020, 9, 334. [Google Scholar] [CrossRef] [PubMed]
- Le, N.T.; Donadu, M.G.; Ho, D.V.; Doan, T.Q.; Le, A.T.; Raal, A.; Usai, D.; Sanna, G.; Marchetti, M.; Usai, M.; et al. Biological activities of essential oil extracted from leaves of Atalantia sessiflora Guillauminin Vietnam. J. Infect. Dev. Ctries. 2020, 14, 1054–1064. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antibiotic (µg) | Phylogenetic Group n (%) | TR (n = 126) | ||||
|---|---|---|---|---|---|---|
| A1 (n = 6) | B21 (n = 12) | B22 (n = 76) | D1 (n = 18) | D2 (n = 14) | ||
| Penicillins | ||||||
| AM-10 | 6 (100) | 12 (100) | 76 (100) | 18 (100) | 13 (92.86) | 125 (99.20) |
| β-Lactam/combination agents | ||||||
| AMC-20/10 | 6 (100) | 11 (91.7) | 52 (68.42) | 15 (83.33) | 14 (100) | 98 (77.77) |
| TZP-100/10 | 6 (100) | 12 (100) | 70 (92.11) | 15 (83.33) | 14 (100) | 117 (92.85) |
| Cephems 1st and 2rd | ||||||
| CF-30 | 6 (100) | 12 (100) | 71 (93.42) | 8 (44.44) | 11 (78.57) | 108 (85.71) |
| CEC-30 | 6 (100) | 9 (75) | 73 (96.05) | 8 (44.44) | 9 (64.29) | 105 (83.33) |
| Cephems 3nd | ||||||
| CTX-30 | 6 (100) | 8 (66.7) | 72 (94.74) | 4 (22.22) | 8 (57.14) | 98 (77.77) |
| CAZ-30 | 6 (100) | 8 (66.7) | 72 (94.74) | 4 (22.22) | 8 (57.14) | 98 (77.77) |
| Monobactams | ||||||
| ATM-30 | 6 (100) | 7 (58.33) | 68 (89.47) | 5 (27.78) | 6 (42.86) | 92 (73.01) |
| Carbapenems | ||||||
| MEM-10 | 0 (0) | 2 (16.7) | 1 (1.31) | 2(11.11) | 5 (35.71) | 10 (7.94) |
| IPM-10 | 0 (0) | 2 (16.7) | 1 (1.31) | 2 (11.11) | 5 (35.71) | 10 (7.94) |
| Aminoglycosides | ||||||
| GM-10 | 4 (66.7) | 5 (41.7) | 42 (55.26) | 11 (61.11) | 7 (50) | 69 (54.76) |
| Tetracyclines | ||||||
| TE-30 | 6 (100) | 10 (83.33) | 65 (85.52) | 15 (83.33) | 11 (78.57) | 107 (84.92) |
| Fluoroquinolones | ||||||
| LVX-5 | 6 (100) | 6 (50) | 73 (96.05) | 6 (33.33) | 7 (50) | 98 (77.77) |
| NOR-10 | 6 (100) | 6 (50) | 73 (96.05) | 6 (33.33) | 7 (50) | 98 (77.77) |
| OFX-5 | 6 (100) | 6 (50) | 73 (96.05) | 6 (33.33) | 7 (50) | 98 (77.77) |
| Folate pathway antagonists | ||||||
| SXT-1.25/23.75 | 3 (50) | 3 (25) | 44 (57.87) | 13 (72.22) | 7 (50) | 70 (55.55) |
| Nitrofurans | ||||||
| F/M-300 | 2 (33.3) | 1 (8.33) | 4 (5.26) | 1 (5.56) | 2 (14.29) | 10 (7.94) |
| ESBLs | ||||||
| 5 (83.33) | 8 (66.66) * | 59 (77.63) ^ + | 3 (16.66) *^ | 6 (42.85) + | 81 (64.28) | |
| Integrons | ||||||
| Int-1 (intl1) | 4 (66.66) | 2 (16.66) °ꝏ | 37 (48.68) ° | 10 (55.55) ꝏ | 7 (50.00) | 60 (47.61) |
| Int-2 (intl2) | 1 (16.66) | 1 (8.33) | 2 (2.63) | 0 (0.00) | 0 (0.00) | 4 (3.17) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras-Alvarado, L.M.; Zavala-Vega, S.; Cruz-Córdova, A.; Reyes-Grajeda, J.P.; Escalona-Venegas, G.; Flores, V.; Alcázar-López, V.; Arellano-Galindo, J.; Hernández-Castro, R.; Castro-Escarpulli, G.; et al. Molecular Epidemiology of Multidrug-Resistant Uropathogenic Escherichia coli O25b Strains Associated with Complicated Urinary Tract Infection in Children. Microorganisms 2021, 9, 2299. https://doi.org/10.3390/microorganisms9112299
Contreras-Alvarado LM, Zavala-Vega S, Cruz-Córdova A, Reyes-Grajeda JP, Escalona-Venegas G, Flores V, Alcázar-López V, Arellano-Galindo J, Hernández-Castro R, Castro-Escarpulli G, et al. Molecular Epidemiology of Multidrug-Resistant Uropathogenic Escherichia coli O25b Strains Associated with Complicated Urinary Tract Infection in Children. Microorganisms. 2021; 9(11):2299. https://doi.org/10.3390/microorganisms9112299
Chicago/Turabian StyleContreras-Alvarado, Laura M., Sergio Zavala-Vega, Ariadnna Cruz-Córdova, Juan Pablo Reyes-Grajeda, Gerardo Escalona-Venegas, Víctor Flores, Virginia Alcázar-López, José Arellano-Galindo, Rigoberto Hernández-Castro, Graciela Castro-Escarpulli, and et al. 2021. "Molecular Epidemiology of Multidrug-Resistant Uropathogenic Escherichia coli O25b Strains Associated with Complicated Urinary Tract Infection in Children" Microorganisms 9, no. 11: 2299. https://doi.org/10.3390/microorganisms9112299