
In Silico/In Vitro Strategies Leading to the Discovery of New Nonribosomal Peptide and Polyketide Antibiotics Active against Human Pathogens


Abstract
:1. Introduction and Historical Context
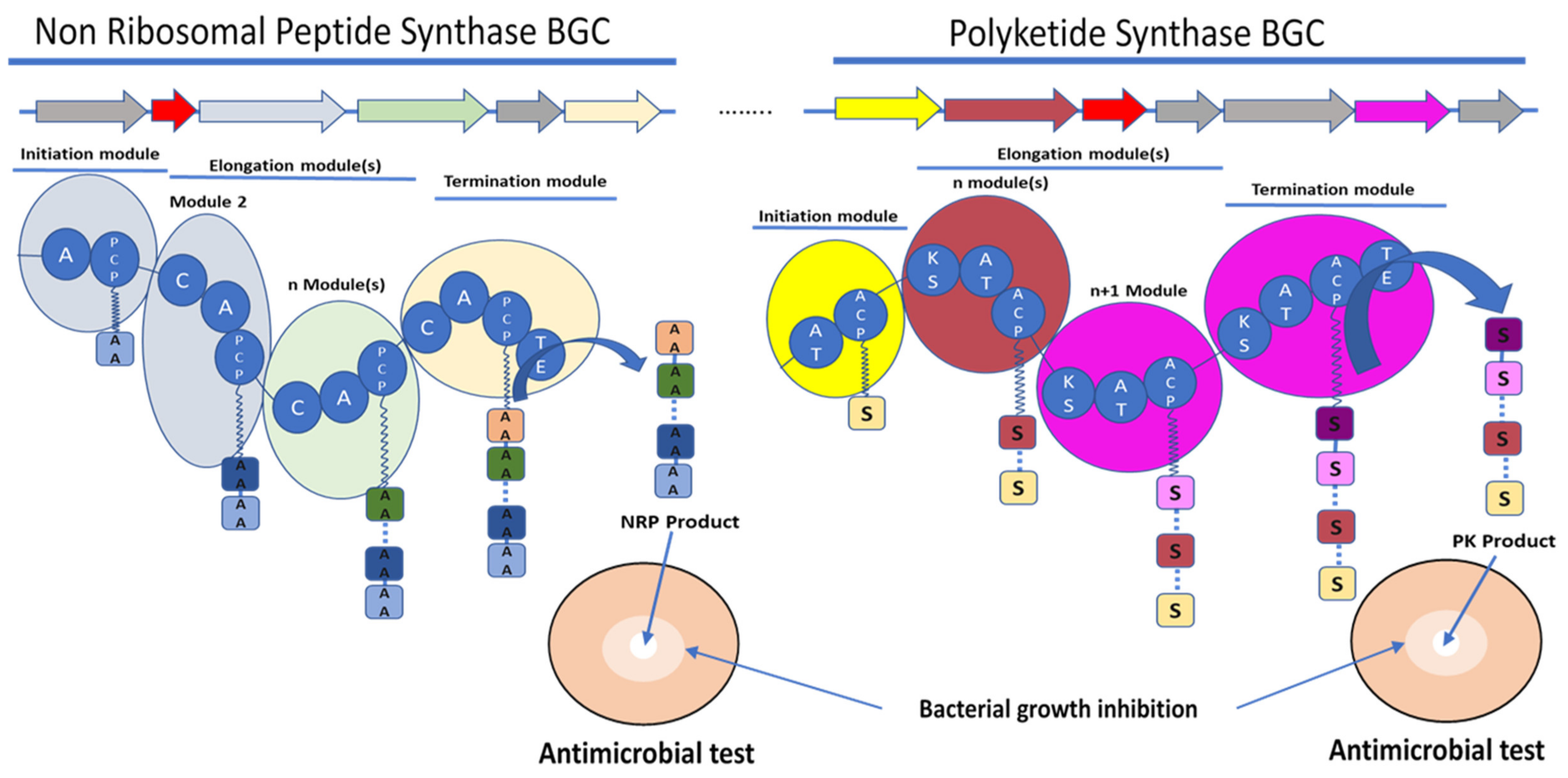
2. Overview of Polyketide Synthase (PKS) and Nonribosomal Peptide Synthase (NRPS)
3. The First Culture-Dependant Discoveries of Cultivable and Uncultivable Micro-Organisms
4. New Discoveries Allowed by Genome-Mining Approaches
5. Antibiotic Production and Resistance
6. Refined Search for New Variants
7. Metagenomic Strategy for Harnessing the Chemical Repertoire in the Environment
8. Synthesis of Natural Antibiotics
9. Dilemma between the Knowledge from In Silico and the Vagaries of In Vitro Methods
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fleming, A. Classics in infectious diseases: On the antibacterial action of cultures of a penicillium, with special reference to their use in the isolation of B. influenzae by Alexander Fleming, Reprinted from the British Journal of Experimental Pathology 10:226-236, 1929. Rev. Infect. Dis. 1980, 2, 129–139. [Google Scholar] [PubMed]
- Gaynes, R. The Discovery of Penicillin—New Insights after More Than 75 Years of Clinical Use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Van Epps, H.L. René Dubos: Unearthing antibiotics. J. Exp. Med. 2006, 203, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiquette, L.; Laupland, K.B. Digging for New Solutions. Can. J. Infect. Dis. Med. Microbiol. 2015, 26, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Lyddiard, D.; Jones, G.L.; Greatrex, B.W. Keeping it simple: Lessons from the golden era of antibiotic discovery. FEMS Microbiol. Lett. 2016, 363, fnw084. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, G.B. Antibacterial Agents from Actinomycetes—A Review. Front. Biosci. 2012, 4, 240–253. [Google Scholar] [CrossRef]
- Lewis, K. Recover the lost art of drug discovery. Nature 2012, 485, 439–440. [Google Scholar] [CrossRef]
- Lewis, K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013, 12, 371–387. [Google Scholar] [CrossRef]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- Pishchany, G.; Kolter, R. On the possible ecological roles of antimicrobials. Mol. Microbiol. 2020, 113, 580–587. [Google Scholar] [CrossRef]
- Van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; Van Wezel, G.P. Ecology and genomics of Actinobacteria: New concepts for natural product discovery. Nat. Rev. Genet. 2020, 18, 1–13. [Google Scholar] [CrossRef]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [CrossRef]
- Felnagle, E.A.; Jackson, E.E.; Chan, Y.A.; Podevels, A.M.; Berti, A.D.; McMahon, M.D.; Thomas, M.G. Nonribosomal Peptide Synthetases Involved in the Production of Medically Relevant Natural Products. Mol. Pharm. 2008, 5, 191–211. [Google Scholar] [CrossRef]
- Walsh, C.T.; Fischbach, M.A. Natural Products Version 2.0: Connecting Genes to Molecules. J. Am. Chem. Soc. 2010, 132, 2469–2493. [Google Scholar] [CrossRef] [Green Version]
- Tracanna, V.; De Jong, A.; Medema, M.H.; Kuipers, O.P. Mining prokaryotes for antimicrobial compounds: From diversity to function. FEMS Microbiol. Rev. 2017, 41, 417–429. [Google Scholar] [CrossRef]
- Malpartida, F.; Hopwood, D.A. Molecular cloning of the whole biosynthetic pathway of a Streptomyces antibiotic and its expression in a heterologous host. Nature 1984, 309, 462–464. [Google Scholar] [CrossRef]
- Cortes, J.; Haydock, S.F.; Roberts, G.A.; Bevitt, D.J.; Leadlay, P. An unusually large multifunctional polypeptide in the erythromycin-producing polyketide synthase of Saccharopolyspora erythraea. Nature 1990, 348, 176–178. [Google Scholar] [CrossRef]
- Stanzak, R.; Matsushima, P.; Baltz, R.H.; Rao, R.N. Cloning and Expression in Streptomyces lividans of Clustered Erythromycin Biosynthesis Genes from Streptomyces erythreus. Nat. Biotechnol. 1986, 4, 229–232. [Google Scholar] [CrossRef]
- Weber, J.; Losick, R. The use of a chromosome integration vector to map erythromycin resistance and production genes in Saccharopolyspora erythraea (Streptomyces erythraeus). Gene 1988, 68, 173–180. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T. Polyketide Synthase Modules Redefined. Angew. Chem. Int. Ed. 2017, 56, 4658–4660. [Google Scholar] [CrossRef]
- Zhang, L.; Hashimoto, T.; Qin, B.; Hashimoto, J.; Kozone, I.; Kawahara, T.; Okada, M.; Awakawa, T.; Ito, T.; Asakawa, Y.; et al. Characterization of Giant Modular PKSs Provides Insight into Genetic Mechanism for Structural Diversification of Aminopolyol Polyketides. Angew. Chem. Int. Ed. 2017, 56, 1740–1745. [Google Scholar] [CrossRef]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Biol. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Zhang, Z.; Pan, H.-X.; Tang, G.-L. New insights into bacterial type II polyketide biosynthesis. F1000Research 2017, 6, 172. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Ogata, H.; Goto, S. Type III Polyketide Synthases: Functional Classification and Phylogenomics. ChemBioChem 2016, 18, 50–65. [Google Scholar] [CrossRef]
- Lim, Y.P.; Go, M.K.; Yew, W.S. Exploiting the Biosynthetic Potential of Type III Polyketide Synthases. Molecules 2016, 21, 806. [Google Scholar] [CrossRef]
- Süssmuth, R.D.; Mainz, A. Nonribosomal Peptide Synthesis-Principles and Prospects. Angew. Chem. Int. Ed. 2017, 56, 3770–3821. [Google Scholar] [CrossRef]
- Strieker, M.; Tanović, A.; Marahiel, M.A. Nonribosomal peptide synthetases: Structures and dynamics. Curr. Opin. Struct. Biol. 2010, 20, 234–240. [Google Scholar] [CrossRef]
- Stanišić, A.; Kries, H. Adenylation Domains in Nonribosomal Peptide Engineering. ChemBioChem 2019, 20, 1347–1356. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Chapple, D.S. Peptide Antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef] [Green Version]
- Stachelhaus, T.; Mootz, H.D.; Marahiel, M.A. The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 1999, 6, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Prieto, C.; Garcia-Estrada, C.; Lorenzana, D.; Martín, J.F. NRPSsp: Non-ribosomal peptide synthase substrate predictor. Bioinformatics 2011, 28, 426–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.V.; Johnson, R.D.; Arcus, V.L.; Lott, J.S. Prediction of the substrate for nonribosomal peptide synthetase (NRPS) adenylation domains by virtual screening. Proteins Struct. Funct. Bioinform. 2015, 83, 2052–2066. [Google Scholar] [CrossRef] [PubMed]
- Chevrette, M.G.; Aicheler, F.; Kohlbacher, O.; Currie, C.R.; Medema, M.H. SANDPUMA: Ensemble predictions of nonribosomal peptide chemistry reveal biosynthetic diversity across Actinobacteria. Bioinformatics 2017, 33, 3202–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachelhaus, T.; Mootz, H.D.; Bergendahl, V.; Marahiel, M.A. Peptide Bond Formation in Nonribosomal Peptide Biosynthesis. J. Biol. Chem. 1998, 273, 22773–22781. [Google Scholar] [CrossRef] [Green Version]
- Bloudoff, K.; Schmeing, T.M. Structural and functional aspects of the nonribosomal peptide synthetase condensation domain superfamily: Discovery, dissection and diversity. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2017, 1865, 1587–1604. [Google Scholar] [CrossRef]
- Owen, J.G.; Calcott, M.J.; Robins, K.J.; Ackerley, D.F. Generating Functional Recombinant NRPS Enzymes in the Laboratory Setting via Peptidyl Carrier Protein Engineering. Cell Chem. Biol. 2016, 23, 1395–1406. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Cobb, R.E.; Zhao, H. Recent advances in natural product discovery. Curr. Opin. Biotechnol. 2014, 30, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Zipperer, A.; Konnerth, M.C.; Laux, C.; Berscheid, A.; Janek, D.; Weidenmaier, C.; Burian, M.; Schilling, N.A.; Slavetinsky, C.; Marschal, M.; et al. Human commensals producing a novel antibiotic impair pathogen colonization. Nature 2016, 535, 511–516. [Google Scholar] [CrossRef]
- Wilson, M.R.; Zha, L.; Balskus, E.P.; Wilson, M.R.; Zha, L.; Balskus, E.P. Natural product discovery from the human microbiome. J. Biol. Chem. 2017, 292, 8546–8552. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S.S.; et al. A new antibiotic kills pathogens without detectable resistance. Nat. Cell Biol. 2015, 517, 455–459. [Google Scholar] [CrossRef]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.M.; Pham, L.; Mehta, A.; Belanger, A.; Kanigan, T.; Lewis, K.; Epstein, S.S. Use of Ichip for High-Throughput In Situ Cultivation of “Uncultivable” Microbial Species. Appl. Environ. Microbiol. 2010, 76, 2445–2450. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Fritz, S.; Rajaonison, A.; Chabrol, O.; Raoult, D.; Rolain, J.-M.; Merhej, V. Full-length title: NRPPUR database search and in vitro analysis identify an NRPS-PKS biosynthetic gene cluster with a potential antibiotic effect. BMC Bioinform. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.; Fedorova, N.D. SMURF: Genomic mapping of fungal secondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef] [Green Version]
- Ziemert, N.; Podell, S.; Penn, K.; Badger, J.H.; Allen, E.; Jensen, P.R. The Natural Product Domain Seeker NaPDoS: A Phylogeny Based Bioinformatic Tool to Classify Secondary Metabolite Gene Diversity. PLoS ONE 2012, 7, e34064. [Google Scholar] [CrossRef] [Green Version]
- Alanjary, M.; Kronmiller, B.; Adamek, M.; Blin, K.; Weber, T.; Huson, D.H.; Philmus, B.; Ziemert, N. The Antibiotic Resistant Target Seeker (ARTS), an exploration engine for antibiotic cluster prioritization and novel drug target discovery. Nucleic Acids Res. 2017, 45, W42–W48. [Google Scholar] [CrossRef]
- Carroll, L.M.; Larralde, M.; Fleck, J.S.; Ponnudurai, R.; Milanese, A.; Barazzone, E.C.; Zeller, G. Accurate de novo identification of biosynthetic gene clusters with GECCO. bioRxiv 2021. [Google Scholar] [CrossRef]
- Li, J.W.-H.; Vederas, J.C. Drug Discovery and Natural Products: End of an Era or an Endless Frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Pidot, S.; Coyne, S.; Kloss, F.; Hertweck, C. Antibiotics from neglected bacterial sources. Int. J. Med. Microbiol. 2014, 304, 14–22. [Google Scholar] [CrossRef]
- Panthee, S.; Hamamoto, H.; Paudel, A.; Sekimizu, K. Lysobacter species: A potential source of novel antibiotics. Arch. Microbiol. 2016, 198, 839–845. [Google Scholar] [CrossRef]
- Sang, M.; Wang, H.; Shen, Y.; De Almeida, N.R.; Conda-Sheridan, M.; Li, S.; Li, Y.; Du, L. Identification of an Anti-MRSA Cyclic Lipodepsipeptide, WBP-29479A1, by Genome Mining of Lysobacter antibioticus. Org. Lett. 2019, 21, 6432–6436. [Google Scholar] [CrossRef]
- Dahal, R.H.; Chaudhary, D.K.; Kim, D.-U.; Pandey, R.P.; Kim, J. Chryseobacterium antibioticum sp. nov. with antimicrobial activity against Gram-negative bacteria, isolated from Arctic soil. J. Antibiot. 2020, 74, 115–123. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Armougom, F.; Million, M.; Hugon, P.; Pagnier, I.; Robert, C.; Bittar, F.; Fournous, G.; Gimenez, G.; Maraninchi, M.; et al. Microbial culturomics: Paradigm shift in the human gut microbiome study. Clin. Microbiol. Infect. 2012, 18, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.-C.; Khelaifia, S.; Alou, M.T.; Ndongo, S.; Dione, N.; Hugon, P.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat. Microbiol. 2016, 1, 16203. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.-M.; Fournier, P.-E.; Raoult, D. Culturing the human microbiota and culturomics. Nat. Rev. Genet. 2018, 16, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Munnoch, J.T.; Devine, R.; Holmes, N.A.; Seipke, R.F.; Wilkinson, K.A.; Wilkinson, B.; Hutchings, M.I. Formicamycins, antibacterial polyketides produced by Streptomyces formicae isolated from African Tetraponera Plant-Ants. Chem. Sci. 2017, 8, 3218–3227. [Google Scholar] [CrossRef] [Green Version]
- Cobb, R.E.; Wang, Y.; Zhao, H. High-Efficiency Multiplex Genome Editing of Streptomyces Species Using an Engineered CRISPR/Cas System. ACS Synth. Biol. 2014, 4, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Liu, N.; Tang, Y. Recent developments in self-resistance gene directed natural product discovery. Nat. Prod. Rep. 2020, 37, 879–892. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.C.; Schorn, M.; Larson, C.B.; Millán-Aguiñaga, N. Targeted antibiotic discovery through biosynthesis-associated resistance determinants: Target directed genome mining. Crit. Rev. Microbiol. 2019, 45, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Diene, S.M.; Pinault, L.; Baron, S.A.; Azza, S.; Armstrong, N.; Hadjadj, L.; Chabrière, E.; Rolain, J.-M.; Pontarotti, P.; Raoult, D. A metallo-β-lactamase enzyme for internal detoxification of the antibiotic thienamycin. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hobson, C.; Chan, A.N.; Wright, G.D. The Antibiotic Resistome: A Guide for the Discovery of Natural Products as Antimicrobial Agents. Chem. Rev. 2021, 121, 3464–3494. [Google Scholar] [CrossRef]
- Kling, A.; Lukat, P.; Almeida, D.V.; Bauer, A.; Fontaine, E.; Sordello, S.; Zaburannyi, N.; Herrmann, J.; Wenzel, S.C.; König, C.; et al. Targeting DnaN for tuberculosis therapy using novel griselimycins. Science 2015, 348, 1106–1112. [Google Scholar] [CrossRef]
- Yarlagadda, V.; Medina, R.; Johnson, T.A.; Koteva, K.P.; Cox, G.; Thaker, M.N.; Wright, G.D. Resistance-Guided Discovery of Elfamycin Antibiotic Producers with Antigonococcal Activity. ACS Infect. Dis. 2020, 6, 3163–3173. [Google Scholar] [CrossRef]
- Panter, F.; Krug, D.; Baumann, S.; Müller, R. Self-resistance guided genome mining uncovers new topoisomerase inhibitors from myxobacteria. Chem. Sci. 2018, 9, 4898–4908. [Google Scholar] [CrossRef] [Green Version]
- Adamek, M.; Alanjary, M.; Ziemert, N. Applied evolution: Phylogeny-based approaches in natural products research. Nat. Prod. Rep. 2019, 36, 1295–1312. [Google Scholar] [CrossRef]
- Kang, H.-S. Phylogeny-guided (meta)genome mining approach for the targeted discovery of new microbial natural products. J. Ind. Microbiol. Biotechnol. 2016, 44, 285–293. [Google Scholar] [CrossRef]
- Amin, D.H.; Borsetto, C.; Tolba, S.; Abolmaaty, A.; Abdallah, N.A.; Wellington, E.M.H. Phylogenic Analysis of NRPS and PKS Genes Associated with Antagonistic Micromonospora Rc5 and Streptomyces Ru87 Isolates. J. Adv. Biol. Biotechnol. 2017, 16, 1–22. [Google Scholar] [CrossRef]
- Guo, J.; Ran, H.; Zeng, J.; Liu, N.; Xin, Z. Tafuketide, a phylogeny-guided discovery of a new polyketide from Talaromyces funiculosus Salicorn 58. Appl. Microbiol. Biotechnol. 2016, 100, 5323–5338. [Google Scholar] [CrossRef]
- Li, Y.-X.; Zhong, Z.; Zhang, W.-P.; Qian, P.-Y. Discovery of cationic nonribosomal peptides as Gram-negative antibiotics through global genome mining. Nat. Commun. 2018, 9, 3273. [Google Scholar] [CrossRef]
- Thaker, M.N.; Wang, W.; Spanogiannopoulos, P.; Waglechner, N.; King, A.M.; Medina, R.; Wright, G. Identifying producers of antibacterial compounds by screening for antibiotic resistance. Nat. Biotechnol. 2013, 31, 922–927. [Google Scholar] [CrossRef]
- Culp, E.J.; Waglechner, N.; Wang, W.; Fiebig-Comyn, A.A.; Hsu, Y.-P.; Koteva, K.; Sychantha, D.; Coombes, B.K.; Van Nieuwenhze, M.S.; Brun, Y.V.; et al. Evolution-guided discovery of antibiotics that inhibit peptidoglycan remodelling. Nature 2020, 578, 582–587. [Google Scholar] [CrossRef]
- Lukoseviciute, L.; Lebedeva, J.; Kuisiene, N. Diversity of Polyketide Synthases and Nonribosomal Peptide Synthetases Revealed Through Metagenomic Analysis of a Deep Oligotrophic Cave. Microb. Ecol. 2020, 81, 110–121. [Google Scholar] [CrossRef]
- Elsaeed, E.; Enany, S.; Hanora, A.; Fahmy, N. Comparative Metagenomic Screening of Aromatic Hydrocarbon Degradation and Secondary Metabolite-Producing Genes in the Red Sea, the Suez Canal, and the Mediterranean Sea. OMICS A J. Integr. Biol. 2020, 24, 541–550. [Google Scholar] [CrossRef]
- Brady, S.F.; Simmons, L.; Kim, J.H.; Schmidt, E.W. Metagenomic approaches to natural products from free-living and symbiotic organisms. Nat. Prod. Rep. 2009, 26, 1488–1503. [Google Scholar] [CrossRef]
- Hug, J.J.; Bader, C.D.; Remškar, M.; Cirnski, K.; Müller, R. Concepts and Methods to Access Novel Antibiotics from Actinomycetes. Antibiotics 2018, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Hover, B.M.; Kim, S.-H.; Katz, M.; Charlop-Powers, Z.; Owen, J.G.; Ternei, M.A.; Maniko, J.; Estrela, A.B.; Molina, H.; Park, S.; et al. Culture-independent discovery of the malacidins as calcium-dependent antibiotics with activity against multidrug-resistant Gram-positive pathogens. Nat. Microbiol. 2018, 3, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Shang, Z.; Lemetre, C.; Ternei, M.A.; Brady, S.F. Cadasides, Calcium-Dependent Acidic Lipopeptides from the Soil Metagenome That Are Active against Multidrug-Resistant Bacteria. J. Am. Chem. Soc. 2019, 141, 3910–3919. [Google Scholar] [CrossRef]
- Peek, J.; Lilic, M.; Montiel, D.; Milshteyn, A.; Woodworth, I.; Biggins, J.B.; Ternei, M.A.; Calle, P.Y.; Danziger, M.; Warrier, T.; et al. Rifamycin congeners kanglemycins are active against rifampicin-resistant bacteria via a distinct mechanism. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Minowa, Y.; Araki, M.; Kanehisa, M. Comprehensive Analysis of Distinctive Polyketide and Nonribosomal Peptide Structural Motifs Encoded in Microbial Genomes. J. Mol. Biol. 2007, 368, 1500–1517. [Google Scholar] [CrossRef]
- Chu, J.; Vila-Farres, X.; Inoyama, D.; Ternei, M.; Cohen, L.J.; Gordon, E.A.; Reddy, B.V.B.; Charlop-Powers, Z.; Zebroski, H.A.; Gallardo-Macias, R.; et al. Discovery of MRSA active antibiotics using primary sequence from the human microbiome. Nat. Chem. Biol. 2016, 12, 1004–1006. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Koirala, B.; Forelli, N.; Vila-Farres, X.; Ternei, M.A.; Ali, T.; Colosimo, D.A.; Brady, S.F. Synthetic-Bioinformatic Natural Product Antibiotics with Diverse Modes of Action. J. Am. Chem. Soc. 2020, 142, 14158–14168. [Google Scholar] [CrossRef] [PubMed]
- Vila-Farres, X.; Chu, J.; Inoyama, D.; Ternei, M.A.; Lemetre, C.; Cohen, L.J.; Cho, W.; Reddy, B.V.B.; Zebroski, H.A.; Freundlich, J.S.; et al. Antimicrobials Inspired by Nonribosomal Peptide Synthetase Gene Clusters. J. Am. Chem. Soc. 2017, 139, 1404–1407. [Google Scholar] [CrossRef] [PubMed]
- Vila-Farres, X.; Chu, J.; Ternei, M.A.; Lemetre, C.; Park, S.; Perlin, D.S.; Brady, S.F. An Optimized Synthetic-Bioinformatic Natural Product Antibiotic Sterilizes Multidrug-Resistant Acinetobacter baumannii-Infected Wounds. mSphere 2018, 3, e00528-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoskisson, P.A.; Seipke, R.F. Cryptic or Silent? The Known Unknowns, Unknown Knowns, and Unknown Unknowns of Secondary Metabolism. mBio 2020, 11, e02642-20. [Google Scholar] [CrossRef]
- Son, S.; Ko, S.-K.; Jang, M.; Lee, J.K.; Ryoo, I.-J.; Lee, J.-S.; Lee, K.H.; Soung, N.-K.; Oh, H.; Hong, Y.-S.; et al. Ulleungamides A and B, Modified α,β-Dehydropipecolic Acid Containing Cyclic Depsipeptides from Streptomyces sp. KCB13F003. Org. Lett. 2015, 17, 4046–4049. [Google Scholar] [CrossRef]
- Son, S.; Hong, Y.-S.; Jang, M.; Heo, K.T.; Lee, B.; Jang, J.-P.; Kim, J.-W.; Ryoo, I.-J.; Kim, W.-G.; Ko, S.-K.; et al. Genomics-Driven Discovery of Chlorinated Cyclic Hexapeptides Ulleungmycins A and B from a Streptomyces Species. J. Nat. Prod. 2017, 80, 3025–3031. [Google Scholar] [CrossRef]
- Zhu, J.-W.; Zhang, S.-J.; Wang, W.-G.; Jiang, H. Strategies for Discovering New Antibiotics from Bacteria in the Post-Genomic Era. Curr. Microbiol. 2020, 77, 3213–3223. [Google Scholar] [CrossRef]
- Lim, Y.H.; Wong, F.T.; Yeo, W.L.; Ching, K.C.; Lim, Y.W.; Heng, E.; Chen, S.; Tsai, D.-J.; Lauderdale, T.-L.; Shia, K.-S.; et al. Auroramycin: A Potent Antibiotic from Streptomyces roseosporus by CRISPR-Cas9 Activation. ChemBioChem 2018, 19, 1716–1719. [Google Scholar] [CrossRef]
- Liu, W.-T.; Lamsa, A.; Wong, W.R.; Boudreau, P.D.; Kersten, R.D.; Peng, Y.; Moree, W.J.; Duggan, B.; Moore, B.; Gerwick, W.H.; et al. MS/MS-based networking and peptidogenomics guided genome mining revealed the stenothricin gene cluster in Streptomyces roseosporus. J. Antibiot. 2013, 67, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Plackett, B. Why big pharma has abandoned antibiotics. Nature 2020, 586, S50–S52. [Google Scholar] [CrossRef]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial resistance in ESKAPE pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Sokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184–193. [Google Scholar] [CrossRef]
- Borsetto, C.; Amos, G.C.A.; Da Rocha, U.N.; Mitchell, A.L.; Finn, R.D.; Laidi, R.F.; Vallin, C.; Pearce, D.A.; Newsham, K.K.; Wellington, E.M.H. Microbial community drivers of PK/NRP gene diversity in selected global soils. Microbiome 2019, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Synthesis Mode | Class | Antibiotics | Organism | Discovery | Spectrum |
|---|---|---|---|---|---|
| RPS | β-Lactams | Penicillin | Penicillium notatum, Penicillium chrysogenum | 1928 | Broad |
| Cephalosporin | Cephalosporium acremonium | 1948 | Broad | ||
| Carbapenem | Streptomyces cattleya | 1976 | Broad | ||
| Monobactam | Chromobacterium violaceum | 1981 | Aerobic Gram-negative bacilli | ||
| Glycopeptides | Vancomycin Teicoplanin | Amycolatopsis orientalis | 1953 | Gram-positive | |
| Actinoplanes teichomyceticus | 1978 | Gram-positive | |||
| Polypeptides | Polymyxin | Paenibacillus polymyxa | 1947 | Gram-negative | |
| Streptogramins | Streptogramin B | Streptomyces graminofaciens | 1953 | Gram-positive | |
| Pristinamycin | Streptomyces pristinaespiralis | 1961 | Gram-positive | ||
| Lincosamides | Lincomycin | Streptomyces lincolnensis | 1963 | Gram-positive and some anaerobic bacteria | |
| Lipopeptides | Daptomycin | Streptomyces roseosporus | 1986 | Gram-positive | |
| PKS | Macrolides | Erythromycin | Streptomyces erythraeus | 1948 | Broad |
| Josamycin | Streptomyces narbonensis var. josamyceticus | 1967 | Broad | ||
| Midecamycin | Streptomyces mycarofaciens | 1975 | Broad | ||
| Spiramycin | Streptomyces ambofaciens | 1952 | Broad | ||
| Fidaxomicin | Dactylosporangium aurantiacum subsp. hamdenesis | 1975 | Broad | ||
| Tetracyclines | Chlortetracycline | Streptomyces aureofaciens, Streptomyces rimosus | 1948 | Broad | |
| Carboxylic acid | Mupirocin | Pseudomonas fluorescens | 1971 | Aerobic Gram-positive and negative | |
| Hybrid NRPS/PKS | Rifamycins | Rifampicin | Streptomyces mediterranei | 1957 | Broad |
| Discovery Approach | Authors Target | Antibiotics | Activity | Synthesis | |
|---|---|---|---|---|---|
| Culture-dependant first discoveries | Ecology guided | Lugdunin | S. aureus (Mrsa, GISA), Enterococcus (VRE) | NRPS | |
| Teixobactin | Gram-positive bacteria, Mycobacterium tuberculosis | NRPS | |||
| In silico/In vitro | Classical screen | Formicamycins | Staphylococcus aureus (MRSA), various pathogenic drug resistant bacteria and fungi | PKS | |
| WBP-29479A1 | Gram positive bacteria including MRSA | NRPS | |||
| Self-resistance guided | Topoisomerase inhibitors | Pyxidilicyline | Antibacterial and anticancerous effect | PKS | |
| Specific interaction modes | Cationic nonribosomal peptides | Brevicidine | Antibiotic resistant gram-negative pathogen and colistin resistant Escherichia coli | NRPS | |
| Cationic nonribosomal peptides | Laterocidine | Antibiotic resistant gram-negative pathogen and colistin resistant Escherichia coli | NRPS | ||
| Calcium-dependent antibiotic | Malacidin | Multiple drug-resistant Gram-positive pathogens | NRPS | ||
| Calcium-dependent antibiotic | Cadasides | Multiple drug-resistant Gram-positive pathogens | NRPS | ||
| Phylogeny guided | Talafun | Escherichia coli | PKS | ||
| Glycopeptide related with new mode of action | Carbomycin | Gram+ bacteria, Staphylococcus aureus (MRSA). | NRPS | ||
| Silent BGC:diverse culture condition | Ulleungmycins A and B | Gram +, quinolone-resistant and methicillin-resistant Staphylococcus aureus. | NRPS | ||
| Silent BGC:Crisp cas9 activation | Auroramycin | Gram-positive bacteria including methicillin resistant Staphylococcus aureus (MRSA) | PKS | ||
| Syn-BNP | Humimycin | Gram + | NRPS | ||
| Paenimucillin A | Gram +/− | NRPS | |||
| Paenimucillin C | Gram - Acinetobacter baumanii | NRPS | |||
| Gladiosyn | Gram +/− | NRPS | |||
| Thurinsyn | Gram +/− Mycobacterium tuberculosis | NRPS | |||
| Mucilasyn | Gram +/− Acinetobacter baumanii | NRPS | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khabthani, S.; Rolain, J.-M.; Merhej, V. In Silico/In Vitro Strategies Leading to the Discovery of New Nonribosomal Peptide and Polyketide Antibiotics Active against Human Pathogens. Microorganisms 2021, 9, 2297. https://doi.org/10.3390/microorganisms9112297
Khabthani S, Rolain J-M, Merhej V. In Silico/In Vitro Strategies Leading to the Discovery of New Nonribosomal Peptide and Polyketide Antibiotics Active against Human Pathogens. Microorganisms. 2021; 9(11):2297. https://doi.org/10.3390/microorganisms9112297
Chicago/Turabian StyleKhabthani, Sami, Jean-Marc Rolain, and Vicky Merhej. 2021. "In Silico/In Vitro Strategies Leading to the Discovery of New Nonribosomal Peptide and Polyketide Antibiotics Active against Human Pathogens" Microorganisms 9, no. 11: 2297. https://doi.org/10.3390/microorganisms9112297