
Complementary Food Ingredients Alter Infant Gut Microbiome Composition and Metabolism In Vitro

 , , and
, , and
Abstract
:1. Introduction
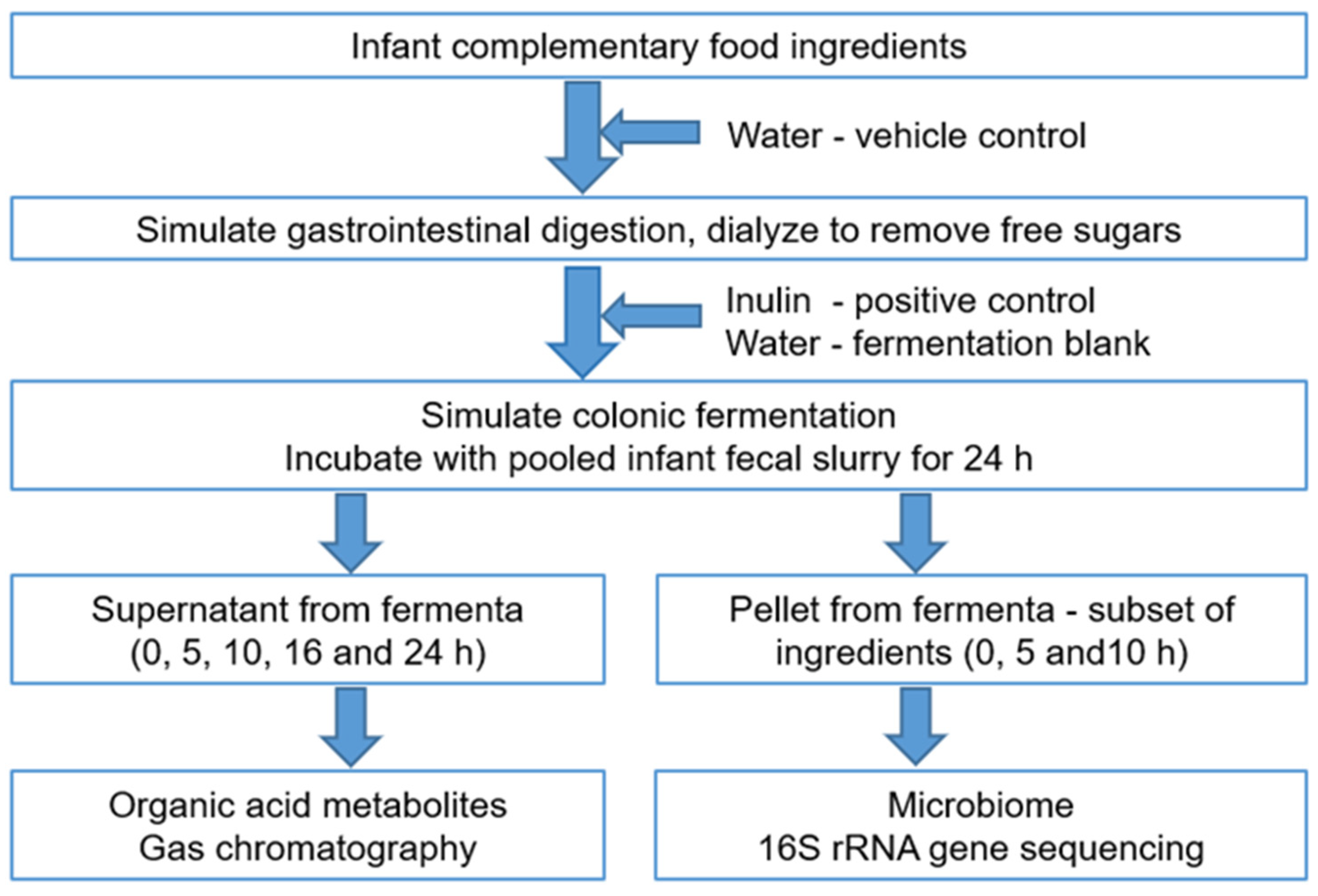
2. Materials and Methods
2.1. Food Ingredients
2.2. In Vitro Simulated Gastroileal Digestion
2.3. Preparation of Fecal Inoculum
2.4. Simulated Colonic Fermentation
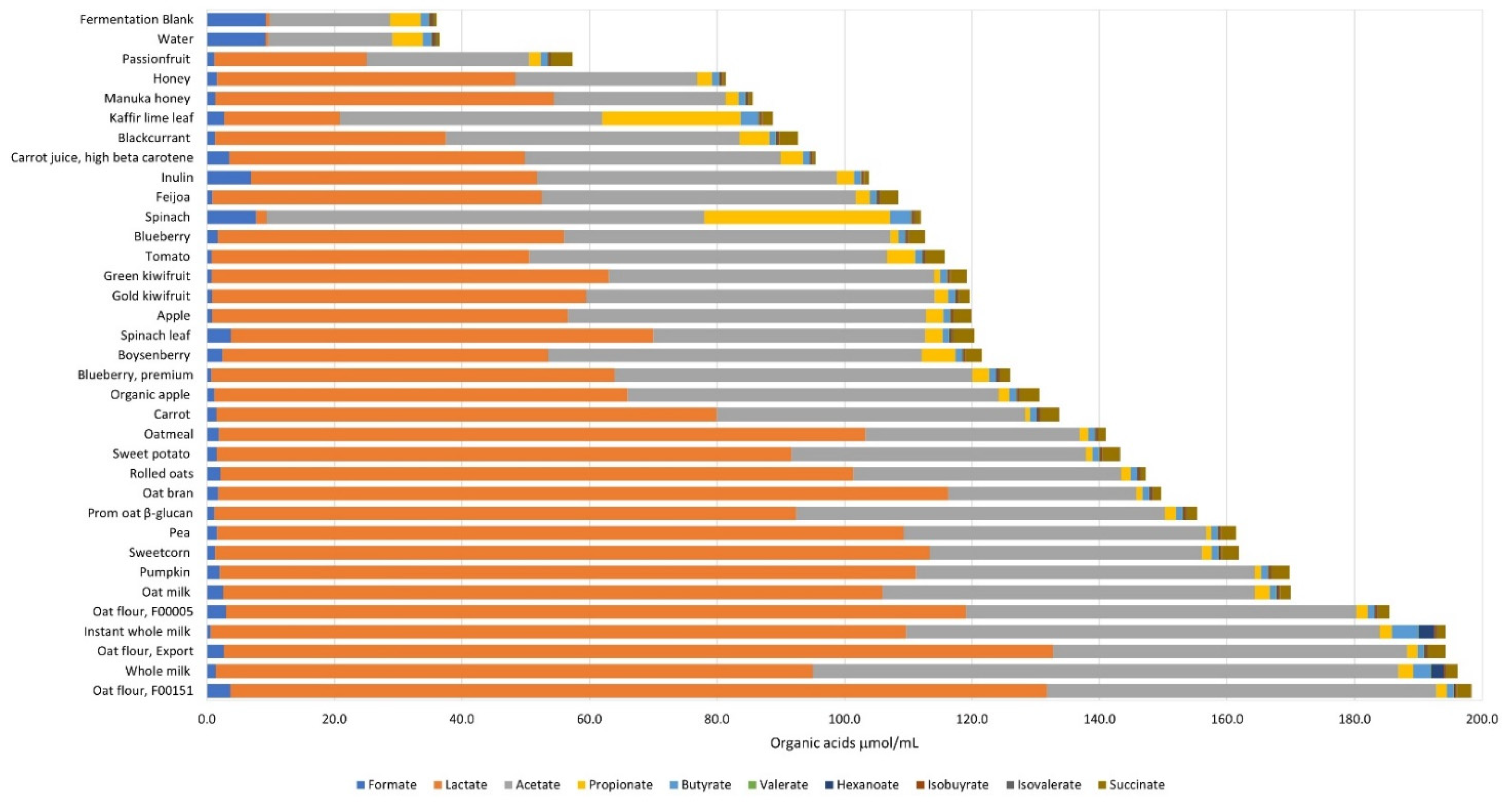
2.5. Analysis of Organic Acid Metabolites
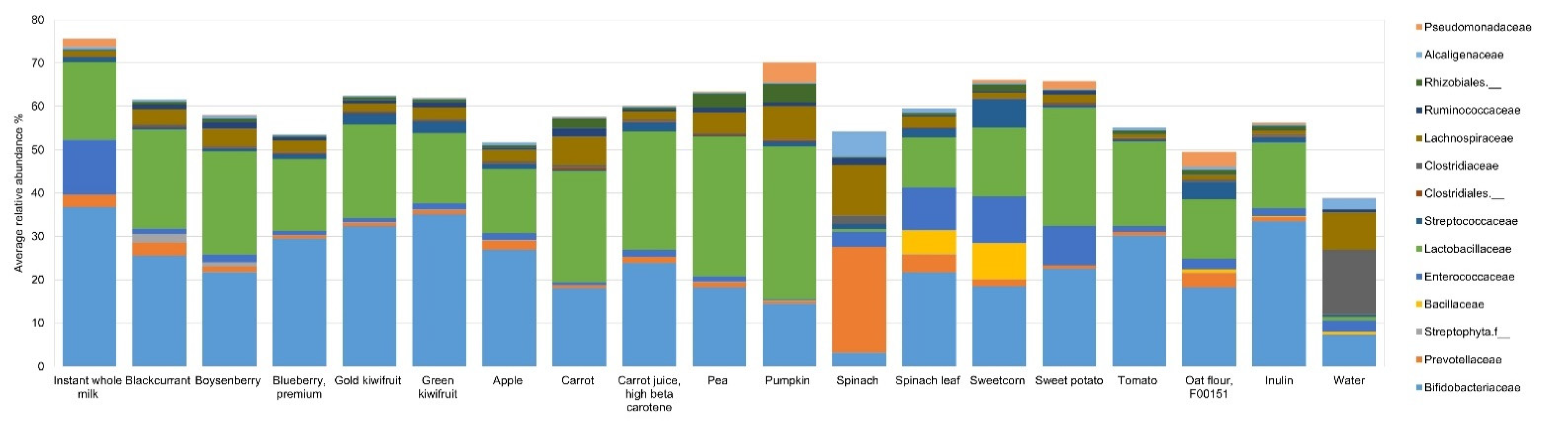
2.6. Characterization of Microbial DNA
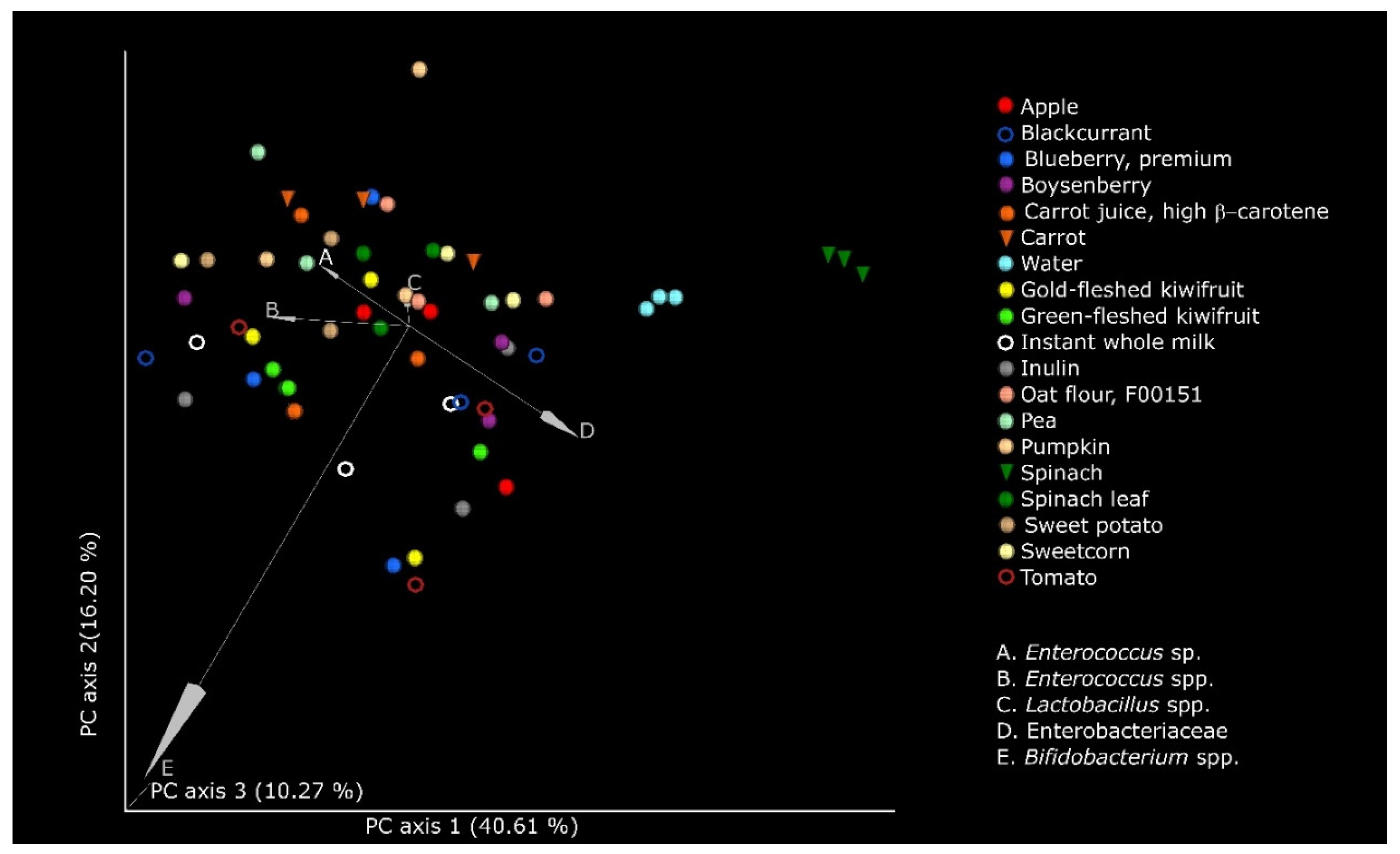
2.7. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laursen, M.F.; Bahl, M.I.; Michaelsen, K.F.; Licht, T.R. First foods and gut microbes. Front. Microbiol. 2017, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koleva, P.T.; Bridgman, S.L.; Kozyrskyj, A.L. The infant gut microbiome: Evidence for obesity risk and dietary intervention. Nutrients 2015, 7, 2237–2260. [Google Scholar] [CrossRef] [Green Version]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Palacio, S.D.; Montes, S.A.; Mancabelli, L.; et al. The first microbial colonizers of the human gut: Composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Solís, G.; Reyes-Gavilan, C.D.L.; Fernández, N.; Margolles, A.; Gueimonde, M. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe 2010, 16, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Tannock, G.W.; Lawley, B.; Munro, K.; Pathmanathan, S.G.; Zhou, S.J.; Makrides, M.; Gibson, R.A.; Sullivan, T.; Prosser, C.G.; Lowry, D.; et al. Comparison of the compositions of the stool microbiotas of infants fed goat milk formula, cow milk-based formula, or breast milk. Appl. Environ. Microbiol. 2013, 79, 3040–3048. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, H.J.M.; Wildeboer-Veloo, A.C.M.; Raangs, G.C.; Wagendorp, A.A.; Klijn, N.; Bindels, J.G.; Welling, G.W. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatric Gastroenterol. Nutr. 2000, 30, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Hove, H.; Nørgaard, H.; Mortensen, P.B. Lactic acid bacteria and the human gastrointestinal tract. Eur. J. Clin. Nutr. 1999, 53, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Hibberd, M.L.; Pettersson, S.; Lee, Y.K. Enterococcus faecalis from healthy infants modulates inflammation through MAPK signaling pathways. PLoS ONE 2014, 9, e97523. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.; Morgan, X.; Wang, X.; Wickens, K.; Purdie, G.; Fitzharris, P.; Otal, A.; Lawley, B.; Stanley, T.; Barthow, C.; et al. Eczema-protective probiotic alters infant gut microbiome functional capacity but not composition: Sub-sample analysis from a RCT. Benef. Microbes 2019, 10, 5–17. [Google Scholar] [CrossRef]
- Gotoh, A.; Katoh, T.; Sakanaka, M.; Ling, Y.; Yamada, C.; Asakuma, S.; Urashima, T.; Tomabechi, Y.; Katayama-Ikegami, A.; Kurihara, S.; et al. Sharing of human milk oligosaccharides degradants within bifidobacterial communities in faecal cultures supplemented with Bifidobacterium bifidum. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Turroni, F.; Duranti, S.; Milani, C.; Lugli, G.A.; Van Sinderen, D.; Ventura, M. Bifidobacterium bifidum: A key member of the early human gut microbiota. Microorganisms 2019, 7, 544. [Google Scholar] [CrossRef] [Green Version]
- Reuter, G. The Lactobacillus and Bifidobacterium microflora of the human intestine: Composition and succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar]
- O’Callaghan, A.; Van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunesova, V.; Lacroix, C.; Schwab, C. Mucin cross-feeding of infant Bifidobacteria and Eubacterium hallii. Microb. Ecol. 2018, 75, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Liu, Q.; Wang, W. Bifidobacterium bifidum TMC3115 ameliorates milk protein allergy in by affecting gut microbiota: A randomized double-blind control trial. J. Food Biochem. 2020, 44, e13489. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Bottacini, F.; Egan, M.; Chamignon, C.; Tondereau, V.; Moriez, R.; Knol, J.; Langella, P.; Eutamene, H.; Smokvina, T.; et al. The infant-derived Bifidobacterium bifidum strain CNCM I-4319 strengthens gut functionality. Microorganisms 2020, 8, 1313. [Google Scholar] [CrossRef]
- Turroni, F.; Ventura, M.; Buttó, L.F.; Duranti, S.; O’Toole, P.; Motherway, M.O.; Van Sinderen, D. Molecular dialogue between the human gut microbiota and the host: A Lactobacillus and Bifidobacterium perspective. Cell. Mol. Life Sci. 2014, 71, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; Bertalan, M.; Borruel, N.; Casellas, F.; Fernandez, L.; Gautier, L.; Hansen, T.; et al. Enterotypes of the human gut microbiome. Nat. Cell Biol. 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Verkhnyatskaya, S.; Ferrari, M.; de Vos, P.; Walvoort, M.T.C. Shaping the infant microbiome with non-digestible carbohydrates. Front. Microbiol. 2019, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Binns, C.W.; Lee, A.H. The early introduction of complementary (solid) foods: A prospective cohort study of infants in Chengdu, China. Nutrients 2019, 11, 760. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.C.; Parkar, S.G.; Gopal, P.K. Developing infant gut microflora and complementary nutrition. J. R. Soc. N. Z. 2020, 50, 1–13. [Google Scholar] [CrossRef]
- Scott, K.P.; Martin, J.C.; Duncan, S.; Flint, H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and Bifidobacteria, In Vitro. FEMS Microbiol. Ecol. 2013, 87, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Makras, L.; De Vuyst, L. The In Vitro inhibition of gram-negative pathogenic bacteria by Bifidobacteria is caused by the production of organic acids. Int. Dairy J. 2006, 16, 1049–1057. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Kelly, C.J.; Battista, K.D.; Schaefer, R.; Lanis, J.M.; Alexeev, E.E.; Wang, R.X.; Onyiah, J.C.; Kominsky, D.J.; Colgan, S.P. Microbial-derived butyrate promotes epithelial barrier function through IL-10 receptor–dependent repression of claudin-2. J. Immunol. 2017, 199, 2976–2984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nat. Cell Biol. 2018, 562, 583–588. [Google Scholar] [CrossRef]
- Sjögren, Y.M.; Tomicic, S.; Lundberg, A.; Böttcher, M.F.; Björkstén, B.; Sverremark-Ekström, E.; Jenmalm, M. Influence of early gut microbiota on the maturation of childhood mucosal and systemic immune responses. Clin. Exp. Allergy 2009, 39, 1842–1851. [Google Scholar] [CrossRef] [Green Version]
- Braegger, C.; Chmielewska, A.; Decsi, T.; Kolacek, S.; Mihatsch, W.; Moreno, L.; Pieścik, M.; Puntis, J.; Shamir, R.; Szajewska, H.; et al. Supplementation of infant formula with probiotics and/or prebiotics: A Systematic review and comment by the ESPGHAN committee on nutrition. J. Pediatric Gastroenterol. Nutr. 2011, 52, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Scholtens, P.; Alles, M.S.; Bindels, J.G.; van der Linde, E.G.M.; Tolboom, J.J.M.; Knol, J. Bifidogenic effects of solid weaning foods with added prebiotic oligosaccharides: A randomised controlled clinical trial. J. Pediatric Gastroenterol. Nutr. 2006, 42, 553–559. [Google Scholar] [CrossRef]
- Stiverson, J.; Williams, T.; Chen, J.; Adams, S.; Hustead, D.; Price, P.; Guerrieri, J.; Deacon, J.; Yu, Z. Prebiotic oligosaccharides: Comparative evaluation using In Vitro cultures of infants’ fecal microbiomes. Appl. Environ. Microbiol. 2014, 80, 7388–7397. [Google Scholar] [CrossRef] [Green Version]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maalouf, J.; Cogswell, M.E.; Bates, M.; Yuan, K.; Scanlon, K.S.; Pehrsson, P.; Gunn, J.P.; Merritt, R.K. Sodium, sugar, and fat content of complementary infant and toddler foods sold in the United States, 2015. Am. J. Clin. Nutr. 2017, 105, 1443–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, R.; Ferguson, E.; Lehrfeld, J. Complementary foods for infant feeding in developing countries: Their nutrient adequacy and improvement. Eur. J. Clin. Nutr. 1998, 52, 764–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslin, K.; Venter, C. Nutritional aspects of commercially prepared infant foods in developed countries: A narrative review. Nutr. Res. Rev. 2017, 30, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Moumin, N.A.; Green, T.J.; Golley, R.K.; Netting, M.J. Are the nutrient and textural properties of Australian commercial infant and toddler foods consistent with infant feeding advice? Br. J. Nutr. 2020, 124, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic-Malinovska, R.; Kuzmanova, S.; Winkelhausen, E. Oligosaccharide profile in fruits and vegetables as sources of prebiotics and functional foods. Int. J. Food Prop. 2014, 17, 949–965. [Google Scholar] [CrossRef]
- Parkar, S.G.; Frost, J.K.T.; Rosendale, D.; Stoklosinski, H.M.; Jobsis, C.M.H.; Hedderley, D.I.; Gopal, P. The sugar composition of the fibre in selected plant foods modulates weaning infants’ gut microbiome composition and fermentation metabolites in vitro. Sci. Rep. 2021, 11, 9292. [Google Scholar] [CrossRef]
- Larsen, N.; de Souza, C.B.; Krych, L.; Cahú, T.B.; Wiese, M.; Kot, W.; Hansen, K.M.; Blennow, A.; Venema, K.; Jespersen, L. Potential of pectins to beneficially modulate the gut microbiota depends on their structural properties. Front. Microbiol. 2019, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- USDA Food & Nutrition Service. Available online: https://www.fns.usda.gov/ (accessed on 8 February 2021).
- Parkar, S.G.; Simmons, L.; Herath, T.D.; Phipps, J.E.; Trower, T.; Hedderley, D.; McGhie, T.K.; Blatchford, P.; Ansell, J.; Sutton, K.H.; et al. Evaluation of the prebiotic potential of five kiwifruit cultivars after simulated gastrointestinal digestion and fermentation with human Faecal bacteria. Int. J. Food Sci. Technol. 2017, 53, 1203–1210. [Google Scholar] [CrossRef]
- Parkar, S.; Jobsis, C.M.; Herath, T.D.; Stoklosinski, H.; van Klink, J.; Sansom, C.E.; Sims, I.M.; Hedderley, D. Metabolic and microbial responses to the complexation of manuka honey with α-cyclodextrin after simulated gastrointestinal digestion and fermentation. J. Funct. Foods 2017, 31, 266–273. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Pollock, J.; Glendinning, L.; Wisedchanwet, T.; Watson, M. The madness of microbiome: Attempting to find consensus “best practice” for 16s microbiome studies. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Flickinger, E.A.; Hatch, T.F.; Wofford, R.C.; Grieshop, C.M.; Murray, S.M.; Fahey, G.C. In Vitro fermentation properties of selected fructooligosaccharide-containing vegetables and In Vivo colonic microbial populations are affected by the diets of healthy human infants. J. Nutr. 2002, 132, 2188–2194. [Google Scholar] [CrossRef] [Green Version]
- Korpela, K.; de Vos, W.M. Early life colonization of the human gut: Microbes matter everywhere. Curr. Opin. Microbiol. 2018, 44, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Biesalski, H.K. Nutrition meets the microbiome: Micronutrients and the microbiota. Ann. N. Y. Acad. Sci. 2016, 1372, 53–64. [Google Scholar] [CrossRef]
- Tuohy, K.M.; Conterno, L.; Gasperotti, M.; Viola, R. Up-regulating the human intestinal microbiome using whole plant foods, polyphenols, and/or fiber. J. Agric. Food Chem. 2012, 60, 8776–8782. [Google Scholar] [CrossRef]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the effects of diet on bacterial metabolism in the large intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef]
- Gomez, J.A.L.; Mukhopadhya, I.; Duncan, S.; Louis, P.; Shaw, S.; Collie-Duguid, E.; Crost, E.; Juge, N.; Flint, H.J. Formate cross-feeding and cooperative metabolic interactions revealed by transcriptomics in co-cultures of acetogenic and amylolytic human colonic bacteria. Environ. Microbiol. 2019, 21, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.A.; Grant, L.J.; Gidley, M.J.; Mikkelsen, D. Gut fermentation of dietary fibres: Physico-chemistry of plant cell walls and implications for health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef] [Green Version]
- Shirai, N.; Homma, C.T.; Kon, C.; Imura, T.; Wada, Y. Japanese toddlers prefer the scent of soy sauce to that of honey with a sweet drink. Food Qual. Prefer. 2020, 86, 104024. [Google Scholar] [CrossRef]
- Sanz, M.L.; Polemis, N.; Morales, V.; Corzo, N.; Drakoularakou, A.; Gibson, A.G.R.; Rastall, R.A. In Vitro investigation into the potential prebiotic activity of honey oligosaccharides. J. Agric. Food Chem. 2005, 53, 2914–2921. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.T.; Steiner, H. The bowel habit of young children. Arch. Dis. Child. 1984, 59, 649–652. [Google Scholar] [CrossRef]
- Palframan, R.J.; Gibson, G.R.; Rastall, R.A. Effect of pH and dose on the growth of gut bacteria on prebiotic carbohydrates In Vitro. Anaerobe 2002, 8, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, L.M.A.; Sundekilde, U.K.; Andersen, H.J.; Nielsen, D.S.; Bertram, H.C. Lactose and bovine milk oligosaccharides synergistically stimulate B. longum subsp. longum growth in a simplified model of the infant gut microbiome. J. Proteome Res. 2019, 18, 3086–3098. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Roy, N.C.; Guo, Y.; Jia, H.; Ryan, L.; Samuelsson, L.; Thomas, A.; Plowman, J.; Clerens, S.; Day, L.; et al. Human breast milk and infant formulas differentially modify the intestinal microbiota in human infants and host physiology in rats. J. Nutr. 2016, 146, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oki, K.; Akiyama, T.; Matsuda, K.; Gawad, A.; Makino, H.; Ishikawa, E.; Oishi, K.; Kushiro, A.; Fujimoto, J. Long-term colonization exceeding six years from early infancy of Bifidobacterium longum subsp. longum in human gut. BMC Microbiol. 2018, 18, 209. [Google Scholar] [CrossRef]
- Casterline, J.J.L.; Oles, A.C.J.; Ku, Y. In Vitro fermentation of various food fiber fractions. J. Agric. Food Chem. 1997, 45, 2463–2467. [Google Scholar] [CrossRef]
- Jonathan, M.C.; van den Borne, J.J.G.C.; Van Wiechen, P.; Da Silva, C.S.; Schols, H.A.; Gruppen, H. In Vitro fermentation of 12 dietary fibres by faecal inoculum from pigs and humans. Food Chem. 2012, 133, 889–897. [Google Scholar] [CrossRef]
- Lunn, J.; Buttriss, J.L. Carbohydrates and dietary fibre. Nutr. Bull. 2007, 32, 21–64. [Google Scholar] [CrossRef]
- Gawkowska, D.; Cybulska, J.; Zdunek, A. Structure-related gelling of pectins and linking with other natural compounds: A review. Polymers 2018, 10, 762. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Heath, A.-L.; Galland, B.; Rehrer, N.; Drummond, L.; Wu, X.-Y.; Bell, T.J.; Lawley, B.; Sims, I.M.; Tannock, G.W. Substrate use prioritization by a coculture of five species of gut bacteria fed mixtures of arabinoxylan, xyloglucan, β-glucan, and pectin. Appl. Environ. Microbiol. 2020, 86, 86. [Google Scholar] [CrossRef]
- Falony, G.; Calmeyn, T.; Leroy, F.; De Vuyst, L. Coculture fermentations of Bifidobacterium species and Bacteroides thetaiotaomicron reveal a mechanistic insight into the prebiotic effect of inulin-type fructans. Appl. Environ. Microbiol. 2009, 75, 2312–2319. [Google Scholar] [CrossRef] [Green Version]
- Fry, S.C. Phenolic components of the primary cell wall. Feruloylated disaccharides of d-galactose and l-arabinose from spinach polysaccharide. Biochem. J. 1982, 203, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Dziedzic, K.; Górecka, D.; Szwengiel, A.; Michniewicz, J.; Drożdżyńska, A.; Walkowiak, J. Interactions between fecal bacteria, bile acids and components of tomato pomace. Food Sci. Biotechnol. 2019, 28, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Franke, T.; Deppenmeier, U. Physiology and central carbon metabolism of the gut bacterium Prevotella copri. Mol. Microbiol. 2018, 109, 528–540. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Martínez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. In Vitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordlund, E.; Aura, A.-M.; Mattila, I.; Kössö, T.; Rouau, X.; Poutanen, K. Formation of phenolic microbial metabolites and short-chain fatty acids from rye, wheat, and oat bran and their fractions in the metabolical in vitro colon model. J. Agric. Food Chem. 2012, 60, 8134–8145. [Google Scholar] [CrossRef] [PubMed]
- Day, L.; Gomez, J.; Øiseth, S.K.; Gidley, M.J.; Williams, B.A. Faster fermentation of cooked carrot cell clusters compared to cell wall fragments In Vitro by porcine feces. J. Agric. Food Chem. 2012, 60, 3282–3290. [Google Scholar] [CrossRef] [PubMed]
- Bakke, A.J.; Carney, E.M.; Higgins, M.; Moding, K.; Johnson, S.L.; Hayes, J.E. Blending dark green vegetables with fruits in commercially available infant foods makes them taste like fruit. Appetite 2020, 150, 104652. [Google Scholar] [CrossRef] [PubMed]
- Moding, K.J.; Ferrante, M.J.; Bellows, L.; Bakke, A.J.; Hayes, J.; Johnson, S.L. Variety and content of commercial infant and toddler vegetable products manufactured and sold in the United States. Am. J. Clin. Nutr. 2018, 107, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gibson, G.R.; Sailer, M.; Theis, S.; Rastall, R.A. Prebiotics inhibit proteolysis by gut bacteria in a host diet-dependent manner: A three-stage continuous in vitro gut model experiment. Appl. Environ. Microbiol. 2020, 86, e02730-19. [Google Scholar] [CrossRef]
- Heredia, N.L.; Labbé, R.G. Clostridium perfringens. In Guide to Foodborne Pathogens; John Wiley and Sons: Hoboken, NJ, USA, 2013; pp. 82–90. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Brief Description | Source |
|---|---|---|
| Instant whole milk | Ingredients: whole milk (cow’s), skim milk (cow’s), lactose, lecithin, vitamins A and D3 | Miraka Limited, Taupo, New Zealand |
| Whole milk | Ingredients: whole milk (cow’s), skim milk (cow’s), lactose | Miraka Limited, Taupo, New Zealand |
| Blackcurrant * | Freshly harvested blackcurrants, destrigged, freeze dried, milled | Sujon, Gibb Holdings (Nelson) Ltd., Nelson, New Zealand |
| Boysenberry * | Freshly harvested Boysenberries, freeze dried, milled | Sujon, Gibb Holdings (Nelson) Ltd., Nelson, New Zealand |
| Blueberry | Freshly harvested blueberries, freeze dried, milled | Sujon, Gibb Holdings (Nelson) Ltd., Nelson, New Zealand |
| Gold-fleshed kiwifruit * | Fruit freeze-dried, milled | Fresh As, Auckland, New Zealand |
| Apple * | ’Braeburn’ apple, freeze-dried, milled; ascorbic acid added as preservative | Fresh As, Auckland, New Zealand |
| Feijoa | Fresh feijoa, freeze-dried, milled | Fresh As, Auckland, New Zealand |
| Passionfruit | Deseeded passionfruit pulp, freeze-dried, milled | Fresh As, Auckland, New Zealand |
| Carrot * | Fresh carrot, freeze-dried, milled | Fresh As, Auckland, New Zealand |
| Spinach* | NZ-grown spinach, freeze-dried, milled | Fresh As, Auckland, New Zealand |
| Kaffir lime leaf | Kaffir lime leaves, freeze-dried, milled | Fresh As, Auckland, New Zealand |
| Blueberry, premium * | Whole blueberries, freeze-dried, milled | NutraDry, Hendra, Australia |
| Green-fleshed kiwifruit * | Fruit puree, freeze-dried, milled | NutraDry, Hendra, Australia |
| Organic apple | Fruit puree, freeze-dried, milled | NutraDry, Hendra, Australia |
| Carrot juice, high beta-carotene * | 100% carrot juice (minus fiber), freeze-dried, milled | NutraDry, Hendra, Australia |
| Spinach leaf * | Whole spinach leaves, pureed, freeze-dried, milled | NutraDry, Hendra, Australia |
| Sweet potato * | Whole orange-fleshed sweet potato, pureed, freeze-dried, milled | NutraDry, Hendra, Australia |
| Sweetcorn | Mature super sweet sweetcorn kernels, pureed, drum-dried, milled | Cedenco Foods New Zealand Ltd., Gisborne, New Zealand |
| Pea * | Peas pureed, drum-dried, milled | Cedenco Foods New Zealand Ltd., Gisborne, New Zealand |
| Pumpkin * | Pumpkin peeled, pureed, drum-dried, milled | Cedenco Foods New Zealand Ltd., Gisborne, New Zealand |
| Tomato * | Tomatoes pureed, concentrated, mixed with starch carrier, drum-dried and milled | Cedenco Foods New Zealand Ltd., Gisborne, New Zealand |
| Oat flour, F00151 * | Milled oat flour | Harraways, Dunedin, New Zealand |
| Oat flour, F00005 | Finely milled oat flour | Harraways, Dunedin, New Zealand |
| Oat flour, Export | Low moisture and water activity | Harraways, Dunedin, New Zealand |
| Rolled oats | Starting material for oat flour | Harraways, Dunedin, New Zealand |
| Oatmeal | Coarser particles than oat flour | Harraways, Dunedin, New Zealand |
| Oat bran | Coarse particles, higher source of fiber than oats | Harraways, Dunedin, New Zealand |
| Oat milk | Dehydrated oat milk | Harraways, Dunedin, New Zealand |
| PromOat® Beta Glucan | Fine oat powder with 35% β-glucan | Tate & Lyle ANZ Pty Ltd., Auckland, New Zealand |
| Honey | 70% honeydew honey, 30% maltodextrin | G & S Foods, Canvastown, New Zealand |
| Mānuka honey | 70% mānuka honey, 30% maltodextrin | G & S Foods, Canvastown, New Zealand |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parkar, S.G.; Rosendale, D.I.; Stoklosinski, H.M.; Jobsis, C.M.H.; Hedderley, D.I.; Gopal, P. Complementary Food Ingredients Alter Infant Gut Microbiome Composition and Metabolism In Vitro. Microorganisms 2021, 9, 2089. https://doi.org/10.3390/microorganisms9102089
Parkar SG, Rosendale DI, Stoklosinski HM, Jobsis CMH, Hedderley DI, Gopal P. Complementary Food Ingredients Alter Infant Gut Microbiome Composition and Metabolism In Vitro. Microorganisms. 2021; 9(10):2089. https://doi.org/10.3390/microorganisms9102089
Chicago/Turabian StyleParkar, Shanthi G., Doug I. Rosendale, Halina M. Stoklosinski, Carel M. H. Jobsis, Duncan I. Hedderley, and Pramod Gopal. 2021. "Complementary Food Ingredients Alter Infant Gut Microbiome Composition and Metabolism In Vitro" Microorganisms 9, no. 10: 2089. https://doi.org/10.3390/microorganisms9102089