
Dietary Microencapsulated Blend of Organic Acids and Plant Essential Oils Affects Intestinal Morphology and Microbiome of Rainbow Trout (Oncorhynchus mykiss)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Facilities
2.2. Diets and Feeding
2.3. Sample Collection
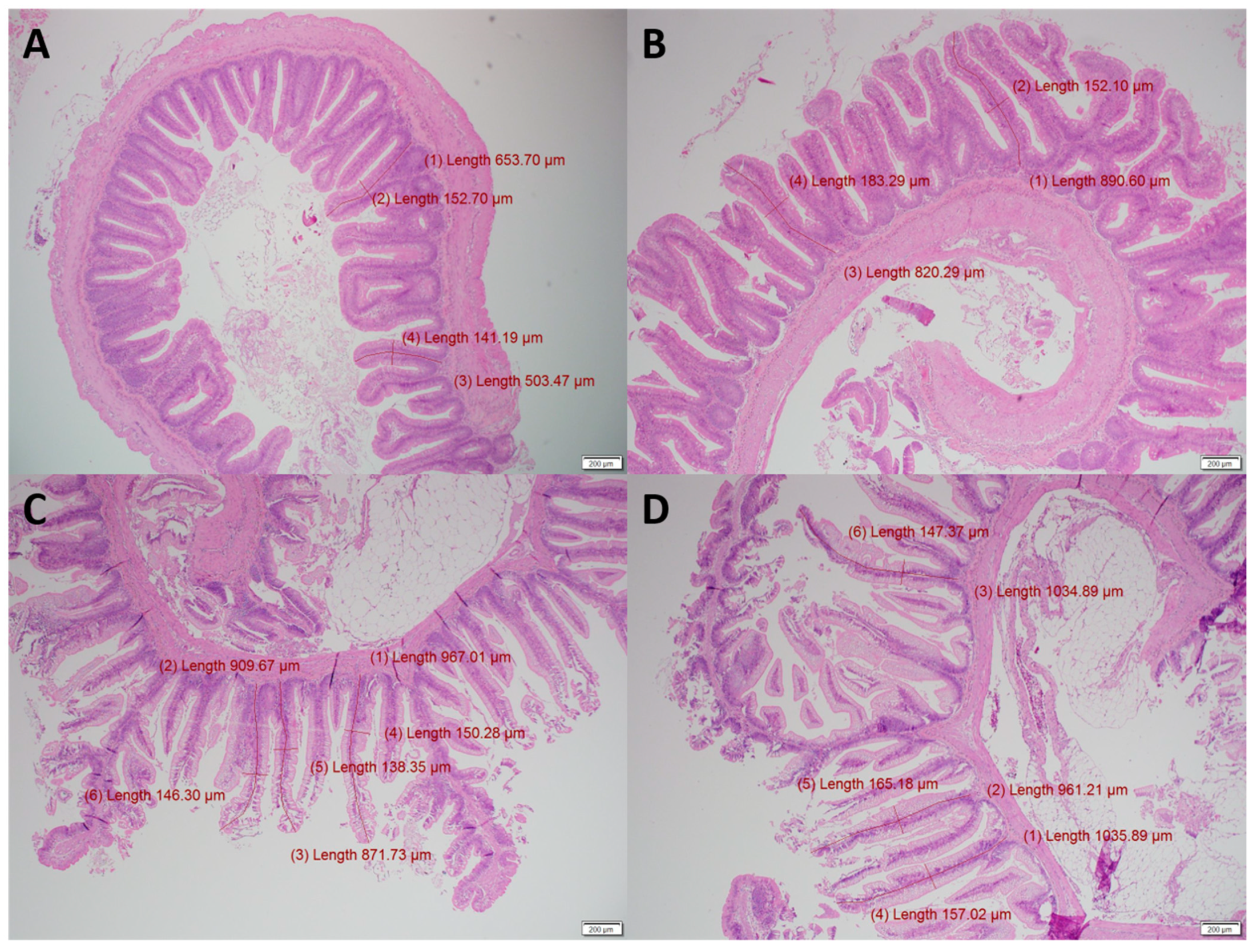
2.4. Histology of Proximal and Distal Intestine
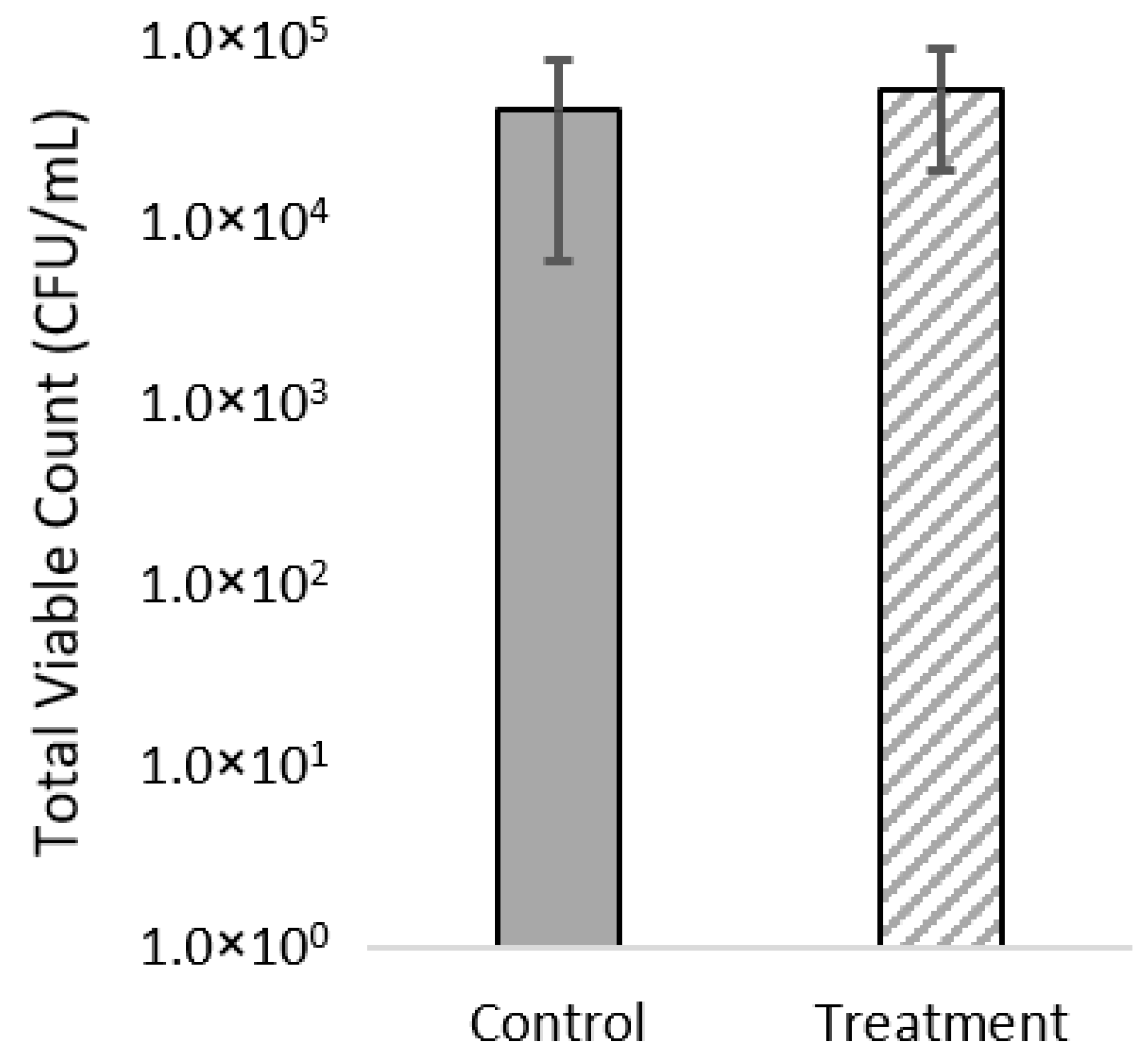
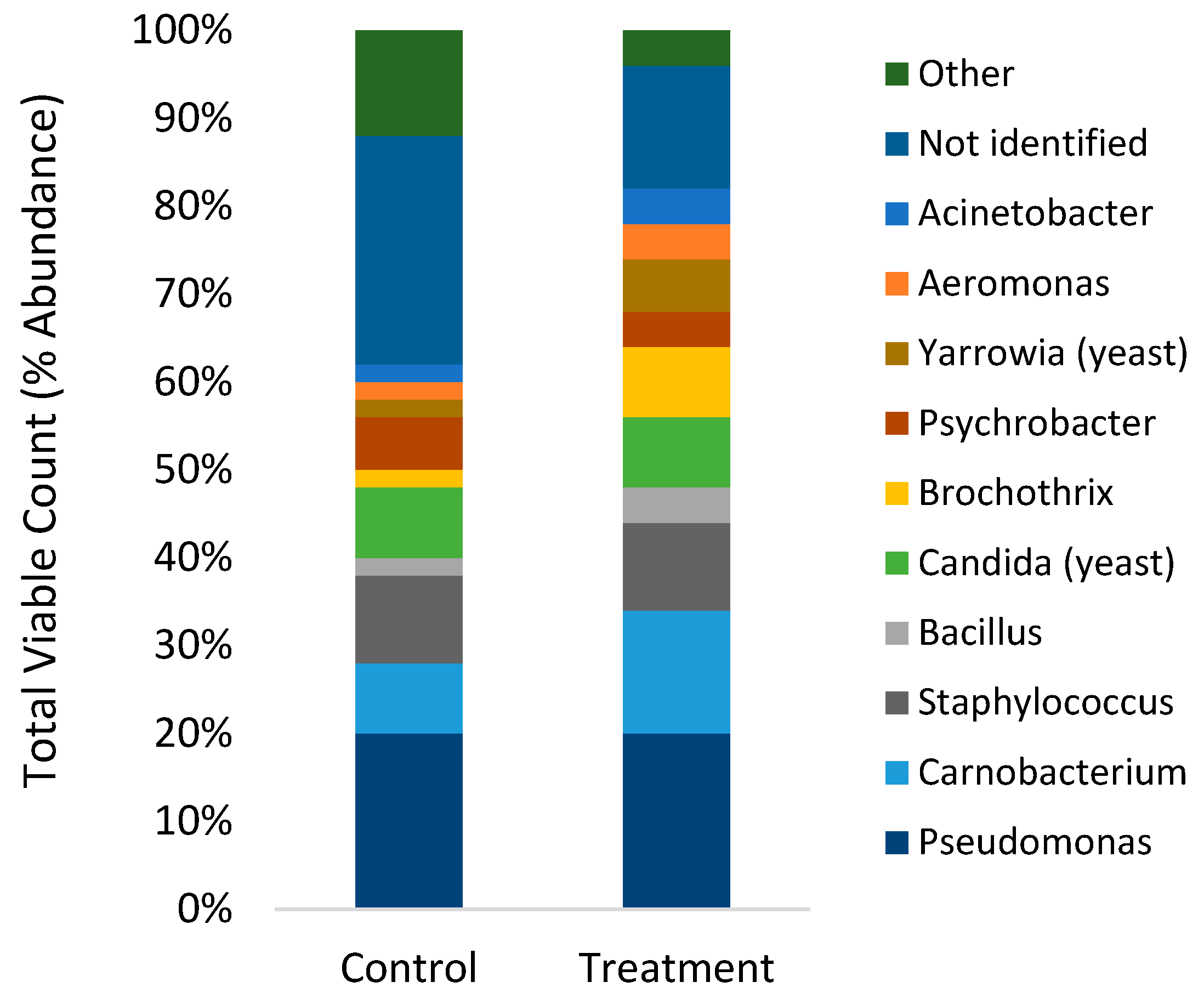
2.5. Viable Plate Counts of Bacteria
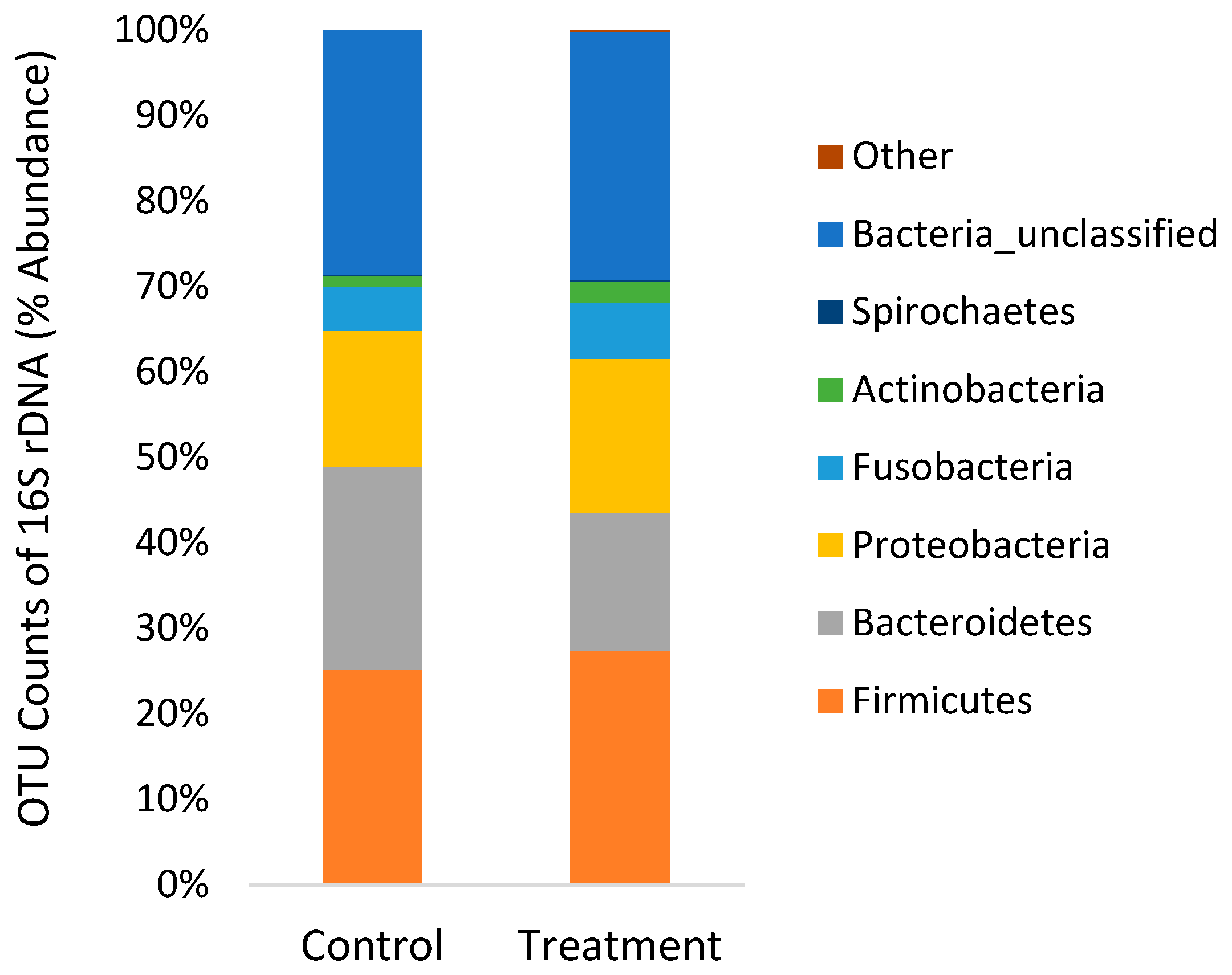
2.6. Extraction and Sequencing of 16S rDNA Bacteria
2.7. Bioinformatics of 16S rDNA Bacteria
2.8. Statistical Analysis
3. Results
3.1. Gut Histology
3.2. Viable Plate Counts and MALDI-TOF of Gut Microbiome
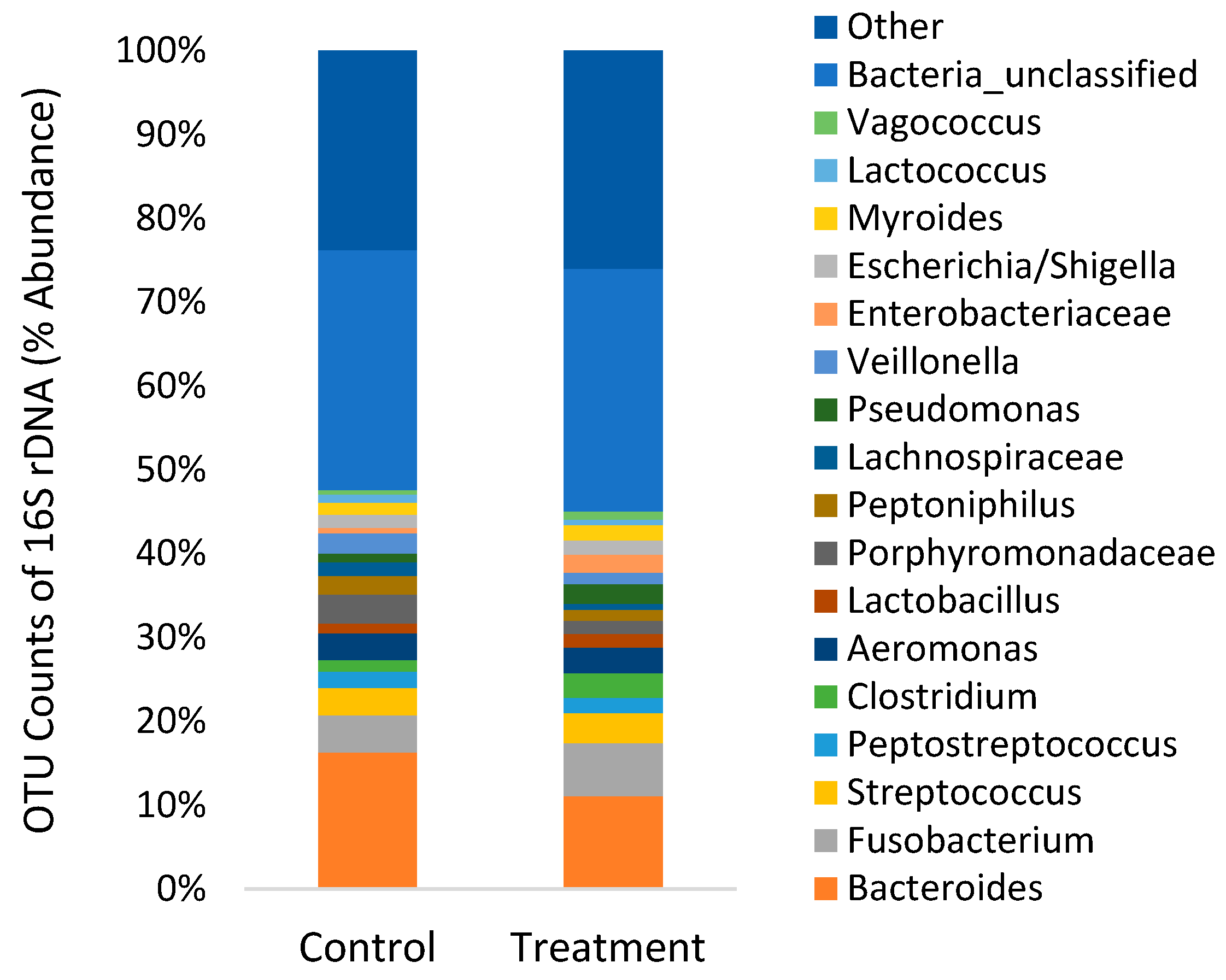
3.3. 16S rDNA Sequencing of Gut Microbiome
3.4. Feeding and Body Indices
4. Discussion
4.1. Gut Histology
4.2. Gut Microbiome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, Z.; Li, X.-Q.; Chowdhury, M.A.K.; Chen, J.-N.; Leng, X.-J. Effects of protease supplementation in low fish meal pelleted and extruded diets on growth, nutrient retention and digestibility of gibel carp, Carassius auratus gibelio. Aquaculture 2016, 460, 37–44. [Google Scholar] [CrossRef]
- Vandenberg, G.W.; Scott, S.L.; Sarker, P.K.; Dallaire, V.; de la Noüe, J. Encapsulation of microbial phytase: Effects on phosphorus bioavailability in rainbow trout (Oncorhynchus mykiss). Anim. Feed Sci. Technol. 2011, 169, 230–243. [Google Scholar] [CrossRef]
- Lee, S.; Small, B.C.; Patro, B.; Overturf, K.; Hardy, R.W. The dietary lysine requirement for optimum protein retention differs with rainbow trout (Oncorhynchus mykiss Walbaum) strain. Aquaculture 2020, 514, 1–28. [Google Scholar] [CrossRef]
- Pandey, A.; Satoh, S. Effects of organic acids on growth and phosphorus utilization in rainbow trout Oncorhynchus mykiss. Fish. Sci. 2008, 74, 867–874. [Google Scholar] [CrossRef]
- Huan, D.; Li, X.; Chowdhury, M.A.K.; Yang, H.; Liang, G.; Leng, X. Organic acid salts, protease and their combination in fish meal-free diets improved growth, nutrient retention and digestibility of tilapia (Oreochromis niloticus × O. aureus). Aquacult. Nutr. 2018, 24, 1813–1821. [Google Scholar] [CrossRef]
- Yao, W.; Li, X.; Kabir Chowdhury, M.A.; Wang, J.; Leng, X. Dietary protease, carbohydrase and micro-encapsulated organic acid salts individually or in combination improved growth, feed utilization and intestinal histology of Pacific white shrimp. Aquaculture 2019, 503, 88–95. [Google Scholar] [CrossRef]
- Diler, O.; Gormez, O.; Diler, I.; Metin, S. Effect of oregano (Origanum onites L.) essential oil on growth, lysozyme and antioxidant activity and resistance against Lactococcus garvieae in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquacult. Nutr. 2017, 23, 844–851. [Google Scholar] [CrossRef]
- Ricke, S.C. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef]
- Ng, W.-K.; Koh, C.-B. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquac. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Baruah, K.; Pal, A.K.; Sahu, N.P.; Jain, K.K.; Mukherjee, S.C.; Debnath, D. Dietary protein level, microbial phytase, citric acid and their interactions on bone mineralization of Labeo rohita (Hamilton) juveniles. Aquacult. Res. 2005, 36, 803–812. [Google Scholar] [CrossRef]
- Baruah, K.; Sahu, N.P.; Pal, A.K.; Jain, K.K.; Debnath, D.; Mukherjee, S.C. Dietary microbial phytase and citric acid synergistically enhances nutrient digestibility and growth performance of Labeo rohita (Hamilton) juveniles at sub-optimal protein level. Aquacult. Res. 2007, 38, 109–120. [Google Scholar] [CrossRef]
- Sarker, S.; Satoh, S.; Kiron, V. Supplementation of citric acid and amino acid-chelated trace element to develop environment-friendly feed for red sea bream, Pagrus major. Aquaculture 2005, 248, 3–11. [Google Scholar] [CrossRef]
- Sarker, M.S.A.; Satoh, S.; Kiron, V. Inclusion of citric acid and/or amino acid-chelated trace elements in alternate plant protein source diets affects growth and excretion of nitrogen and phosphorus in red sea bream Pagrus major. Aquaculture 2007, 262, 436–443. [Google Scholar] [CrossRef]
- Vielma, J.; Ruohonen, K.; Lall, S. Supplemental citric acid and particle size of fish bone-meal influence the availability of minerals in rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Nutr. 2001, 5, 65–71. [Google Scholar] [CrossRef]
- Edris, A.E. Pharmaceutical and therapeutic Potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. 2007, 21, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complementary Altern. Med. 2016, 2016, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.d.F.; Baldissera, M.D.; Baldisserotto, B.; Heinzmann, B.M.; Martos-Sitcha, J.A.; Mancera, J.M. Essential oils as stress-reducing agents for fish aquaculture: A review. Front. Physiol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Chowdhury, M.A.K.; Huo, Y.; Gong, J. Phytogenic compounds as alternatives to in-feed antibiotics: Potentials and challenges in application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef] [Green Version]
- Sutili, F.J.; Gatlin, D.M., III; Heinzmann, B.M.; Baldisserotto, B. Plant essential oils as fish diet additives: Benefits on fish health and stability in feed. Rev. Aquac. 2018, 10, 716–726. [Google Scholar] [CrossRef]
- Sönmez, A.Y.; Bilen, S.; Alak, G.; Hisar, O.; Yanık, T.; Biswas, G. Growth performance and antioxidant enzyme activities in rainbow trout (Oncorhynchus mykiss) juveniles fed diets supplemented with sage, mint and thyme oils. Fish Physiol. Biochem. 2015, 41, 165–175. [Google Scholar] [CrossRef]
- Sönmez, A.Y.; Bilen, S.; Albayrak, M.; Yılmaz, S.; Biswas, G.; Hisar, O.; Yanık, T. Effects of dietary supplementation of herbal oils containing 1, 8-cineole, carvacrol or pulegone on growth performance, survival, fatty acid composition, and liver and kidney histology of rainbow trout (Oncorhynchus mykiss) fingerlings. Turk. J. Fish. Aquat. Sci. 2015, 15, 813–819. [Google Scholar] [CrossRef]
- Giannenas, I.; Triantafillou, E.; Stavrakakis, S.; Margaroni, M.; Mavridis, S.; Steiner, T.; Karagouni, E. Assessment of dietary supplementation with carvacrol or thymol containing feed additives on performance, intestinal microbiota and antioxidant status of rainbow trout (Oncorhynchus mykiss). Aquaculture 2012, 350, 26–32. [Google Scholar] [CrossRef]
- Boyd, C.E.; D’Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.J.; Teletchea, F.; Tomasso, J.R., Jr.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. J. World Aquacult. Soc. 2020, 51, 578–633. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento, M.; Ouwehand, A.; Tiihonen, K.; Lahtinen, S.; Nurminen, P.H.; Saarinen, M.; Schulze, H.; Mygind, T.; Fischer, J. Essential oils and their use in animal feeds for monogastric animals—Effects on feed quality, gut microbiota, growth performance and food safety: A review. Veterinarni Med. 2018, 58, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 100, 296–305. [Google Scholar] [CrossRef]
- Thapa, D.; Louis, P.; Losa, R.; Zweifel, B.; Wallace, R.J. Essential oils have different effects on human pathogenic and commensal bacteria in mixed faecal fermentations compared with pure cultures. Microbiology 2015, 161, 441–449. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Freccia, A.; Sousa, S.M.d.N.; Meurer, F.; Butzge, A.J.; Mewes, J.K.; Bombardelli, R.A. Essential oils in the initial phase of broodstock diets of Nile tilapia. Rev. Brasileira Zootec. 2014, 43, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sutili, F.J.; Velasquez, A.; Pinheiro, C.G.; Heinzmann, B.M.; Gatlin, D.M.; Baldisserotto, B. Evaluation of Ocimum americanum essential oil as an additive in red drum (Sciaenops ocellatus) diets. Fish Shellfish. Immunol. 2016, 56, 155–161. [Google Scholar] [CrossRef]
- Navarrete, P.; Toledo, I.; Mardones, P.; Opazo, R.; Espejo, R.; Romero, J. Effect of Thymus vulgaris essential oil on intestinal bacterial microbiota of rainbow trout, Oncorhynchus mykiss (Walbaum) and bacterial isolates. Aquacult. Res. 2010, 41, e667–e678. [Google Scholar] [CrossRef]
- Sutaphanit, P.; Chitprasert, P. Optimisation of microencapsulation of holy basil essential oil in gelatin by response surface methodology. Food Chem. 2014, 150, 313–320. [Google Scholar] [CrossRef]
- Piva, A.; Pizzamiglio, V.; Morlacchini, M.; Tedeschi, M.; Piva, G. Lipid microencapsulation allows slow release of organic acids and natural identical flavors along the swine intestine. J. Anim. Sci. 2007, 85, 486–493. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.K.; Song, H.; Liu, Y.; Bunod, J.-D.; Dong, X. Effects of Microencapsulated Organic Acid and Their Salts on Growth Performance, Immunity, and Disease Resistance of Pacific White Shrimp Litopenaeus vannamei. Sustainability 2021, 13, 7791. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, 1–11. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2013, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R-Core-Team. R: A Language and Environment for Statistical Computing, 3.2.2; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Liu, W.; Yang, Y.; Zhang, J.; Gatlin, D.M.; Ringø, E.; Zhou, Z. Effects of dietary microencapsulated sodium butyrate on growth, intestinal mucosal morphology, immune response and adhesive bacteria in juvenile common carp (Cyprinus carpio) pre-fed with or without oxidised oil. Brit. J. Nutr. 2014, 112, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wang, H.; Zhang, K.; Gao, F.; Chen, S.; Li, D. Physicochemical properties and storage stability of microencapsulated DHA-rich oil with different wall materials. Appl. Biochem. Biotechnol. 2016, 179, 1129–1142. [Google Scholar] [CrossRef]
- Pelusio, N.F.; Rossi, B.; Parma, L.; Volpe, E.; Ciulli, S.; Piva, A.; D’Amico, F.; Scicchitano, D.; Candela, M.; Gatta, P.P.; et al. Effects of increasing dietary level of organic acids and nature-identical compounds on growth, intestinal cytokine gene expression and gut microbiota of rainbow trout (Oncorhynchus mykiss) reared at normal and high temperature. Fish Shellfish. Immunol. 2020, 107, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Khojasteh, B.; Mahdi, S. The morphology of the post-gastric alimentary canal in teleost fishes: A brief review. Int. J. Aquat. Sci. 2012, 3, 71–88. [Google Scholar]
- Brum, A.; Pereira, S.A.; Owatari, M.S.; Chagas, E.C.; Chaves, F.C.M.; Mouriño, J.L.P.; Martins, M.L. Effect of dietary essential oils of clove basil and ginger on Nile tilapia (Oreochromis niloticus) following challenge with Streptococcus agalactiae. Aquaculture 2017, 468, 235–243. [Google Scholar] [CrossRef]
- Zheng, Z.L.; Tan, J.Y.W.; Liu, H.Y.; Zhou, X.H.; Xiang, X.; Wang, K.Y. Evaluation of oregano essential oil (Origanum heracleoticum L.) on growth, antioxidant effect and resistance against Aeromonas hydrophila in channel catfish (Ictalurus punctatus). Aquaculture 2009, 292, 214–218. [Google Scholar] [CrossRef]
- Al-Sagheer, A.A.; Mahmoud, H.K.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Supplementation of diets for Oreochromis niloticus with essential oil extracts from lemongrass (Cymbopogon citratus) and geranium (Pelargonium graveolens) and effects on growth, intestinal microbiota, antioxidant and immune activities. Aquacult. Nutr. 2018, 24, 1006–1014. [Google Scholar] [CrossRef]
- He, W.; Rahimnejad, S.; Wang, L.; Song, K.; Lu, K.; Zhang, C. Effects of organic acids and essential oils blend on growth, gut microbiota, immune response and disease resistance of Pacific white shrimp (Litopenaeus vannamei) against Vibrio parahaemolyticus. Fish Shellfish. Immunol. 2017, 70, 164–173. [Google Scholar] [CrossRef]
- Huyben, D.; Nyman, A.; Vidaković, A.; Passoth, V.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Effects of dietary inclusion of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus on gut microbiota of rainbow trout. Aquaculture 2017, 473, 528–537. [Google Scholar] [CrossRef]
- Huyben, D.; Sun, L.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Dietary live yeast and increased water temperature influence the gut microbiota of rainbow trout. J. Appl. Microbiol. 2018, 124, 1377–1392. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Vidaković, A.; Werner Hallgren, S.; Langeland, M. High-throughput sequencing of gut microbiota in rainbow trout (Oncorhynchus mykiss) fed larval and pre-pupae stages of black soldier fly (Hermetia illucens). Aquaculture 2019, 500, 485–491. [Google Scholar] [CrossRef]
- Michl, S.C.; Ratten, J.-M.; Beyer, M.; Hasler, M.; LaRoche, J.; Schulz, C. The malleable gut microbiome of juvenile rainbow trout (Oncorhynchus mykiss): Diet-dependent shifts of bacterial community structures. PLoS ONE 2017, 12, e0177735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Exploring the microbial diversity of the distal intestinal lumen and mucosa of farmed rainbow trout Oncorhynchus mykiss (Walbaum) using next generation sequencing (NGS). Aquacult. Res. 2017, 48, 77–91. [Google Scholar] [CrossRef]
- Zhou, Z.; Yao, B.; Romero, J.; Waines, P.; Ringø, E.; Emery, M.; Liles, M.R.; Merrifield, D.L. Methodological approaches used to assess fish gastrointestinal communities. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; Merrifield, D.L., Ringø, E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 101–127. [Google Scholar]
- Gómez-García, M.; Argüello, H.; Puente, H.; Mencía-Ares, Ó.; González, S.; Miranda, R.; Rubio, P.; Carvajal, A. In-depth in vitro evaluation of the activity and mechanisms of action of organic acids and essential oils against swine enteropathogenic bacteria. Front. Vet. Sci. 2020, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.; Liang, H.; Yuan, Q.; Li, C. In vitro antimicrobial effects and mechanism of action of selected plant essential oil combinations against four food-related microorganisms. Food Res. Int. 2011, 44, 3057–3064. [Google Scholar] [CrossRef]
- Dibner, J.J.; Buttin, P. Use of organic acids as a model to study the impact of gut microflora on nutrition and metabolism. J. Appl. Poult. Res. 2002, 11, 453–463. [Google Scholar] [CrossRef]
- Mzula, A.; Wambura, P.N.; Mdegela, R.H.; Shirima, G.M. Current state of modern biotechnological-based Aeromonas hydrophila vaccines for aquaculture: A systematic review. BioMed Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Patil, H.J.; Benet-Perelberg, A.; Naor, A.; Smirnov, M.; Ofek, T.; Nasser, A.; Minz, D.; Cytryn, E. Evidence of increased antibiotic resistance in phylogenetically-diverse Aeromonas isolates from semi-intensive fish ponds treated with antibiotics. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Joly-Guillou, M.L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Koh, C.-B.; Romano, N.; Zahrah, A.S.; Ng, W.-K. Effects of a dietary organic acids blend and oxytetracycline on the growth, nutrient utilization and total cultivable gut microbiota of the red hybrid tilapia, Oreochromis sp., and resistance to Streptococcus agalactiae. Aquacult. Res. 2016, 47, 357–369. [Google Scholar] [CrossRef]
- Soltani, M.; Ghodratnama, M.; Ebrahimzadeh-Mosavi, H.A.; Nikbakht-Brujeni, G.; Mohamadian, S.; Ghasemian, M. Shirazi thyme (Zataria multiflora Boiss) and Rosemary (Rosmarinus officinalis) essential oils repress expression of sagA, a streptolysin S-related gene in Streptococcus iniae. Aquaculture 2014, 430, 248–252. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | g/kg |
|---|---|
| Fish meal-anchovy, 67% CP | 50.0 |
| Poultry by-product meal, 60% CP | 150.0 |
| Distillers dried grain soluble, 23% CP | 100.0 |
| Corn gluten meal, 60% CP | 50.0 |
| Dehulled soybean meal, 48% CP | 255.0 |
| Corn | 125.0 |
| Wheat, grain | 125.0 |
| Wheat middling | 80.0 |
| Fish oil * | 20.0 |
| Soybean oil | 20.0 |
| Premix | 10.0 |
| Lysine HCl | 4.3 |
| Methionine | 1.7 |
| Choline chloride | 3.0 |
| Ca(H2PO4)2 | 5.0 |
| NaCl | 1.0 |
| Proximate composition | |
| Dry matter, % | 91.3 |
| Crude protein, % | 37.4 |
| Lipid *, % | 9.0 |
| Ash, % | 6.8 |
| Gross energy, MJ/kg | 16.2 |
| Proximal | Distal | p-Value 1 | |||||
|---|---|---|---|---|---|---|---|
| Control | Treatment | Control | Treatment | SE | Proximal | Distal | |
| Villi length (µm) | 645 | 686 | 989 | 1039 | 193 | 0.035 | 0.278 |
| Villi width (µm) | 155 | 158 | 150 | 149 | 10 | 0.497 | 0.968 |
| Edema | 0.67 | 0.70 | 0.22 | 0.00 | 0.59 | 1.000 | 0.343 |
| Inflammation serosa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.000 | 1.000 |
| Inflammation submucosa | 0.20 | 0.10 | 0.00 | 0.00 | 0.27 | 0.583 | 1.000 |
| Inflammation lamina propria | 0.20 | 0.10 | 0.22 | 0.00 | 0.41 | 0.583 | 0.343 |
| Vacuolization | 1.00 | 1.00 | 2.78 | 3.00 | 1.01 | 1.000 | 0.343 |
| Goblet cells | 2.30 | 2.80 | 1.22 | 1.00 | 0.93 | 0.139 | 0.343 |
| Mitoses | 1.50 | 1.10 | 0.22 | 0.20 | 0.81 | 0.239 | 0.954 |
| Necrosis/apoptosis | 0.30 | 0.10 | 0.11 | 0.00 | 0.34 | 0.301 | 0.343 |
| Control | Treatment | SE | p-Values | |
|---|---|---|---|---|
| Good’s coverage | 0.989 | 0.989 | 0.001 | 0.841 |
| No. of taxa | 483 | 448 | 104 | 0.691 |
| Shannon diversity | 3.95 | 3.91 | 0.05 | 0.781 |
| Chao-1 richness | 1833 | 2166 | 91 | 0.210 |
| Diet | Phyla | Family/Genus | LDA | p-Value |
|---|---|---|---|---|
| Control | Bacteroidetes | Bacteroides | 4.142 | 0.020 |
| Bacteroidetes | Porphyromonadaceae | 4.144 | 0.009 | |
| Firmicutes | Sporosarcina | 3.246 | 0.048 | |
| Firmicutes | Veillonella | 4.046 | 0.018 | |
| Proteobacteria | Aeromonas | 3.867 | 0.041 | |
| Proteobacteria | Acinetobacter | 3.056 | 0.050 | |
| Treatment | Firmicutes | Streptococcus | 3.700 | 0.041 |
| Fusobacteria | Fusobacterium | 4.175 | 0.040 | |
| Proteobacteria | Escherichia/Shigella | 3.959 | 0.045 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huyben, D.; Chiasson, M.; Lumsden, J.S.; Pham, P.H.; Chowdhury, M.A.K. Dietary Microencapsulated Blend of Organic Acids and Plant Essential Oils Affects Intestinal Morphology and Microbiome of Rainbow Trout (Oncorhynchus mykiss). Microorganisms 2021, 9, 2063. https://doi.org/10.3390/microorganisms9102063
Huyben D, Chiasson M, Lumsden JS, Pham PH, Chowdhury MAK. Dietary Microencapsulated Blend of Organic Acids and Plant Essential Oils Affects Intestinal Morphology and Microbiome of Rainbow Trout (Oncorhynchus mykiss). Microorganisms. 2021; 9(10):2063. https://doi.org/10.3390/microorganisms9102063
Chicago/Turabian StyleHuyben, David, Marcia Chiasson, John S. Lumsden, Phuc H. Pham, and Mohiuddin A. Kabir Chowdhury. 2021. "Dietary Microencapsulated Blend of Organic Acids and Plant Essential Oils Affects Intestinal Morphology and Microbiome of Rainbow Trout (Oncorhynchus mykiss)" Microorganisms 9, no. 10: 2063. https://doi.org/10.3390/microorganisms9102063