Tularemia as a Mosquito-Borne Disease



Abstract
:1. Francisella tularensis and Tularemia
1.1. Historical Background
1.2. Francisella tularensis
1.3. Tularemia Geographical Distribution
1.4. Tularemia, the Disease
2. Review Objectives
2.1. Mosquito
2.1.1. Taxonomy
2.1.2. Geographical Distribution of Arthropod Vector Genera
2.1.3. Mosquito-Borne Diseases
3. Mosquitoes and Francisella
3.1. Human Cases of Tularemia Related to Mosquito Bites
3.1.1. Geographical Areas Concerned: Epidemiology, Climate, Seasonality, and Type of Landscapes
3.1.2. Gender and Age Influence
3.1.3. Clinical Manifestations
3.2. Mosquito Species Associated with Tularemia
3.2.1. Major Mosquito Species Associated with Tularemia by Geographic Area
3.2.2. Particularity of Mosquito Species Associated with Tularemia
3.3. Mosquito Life Cycle
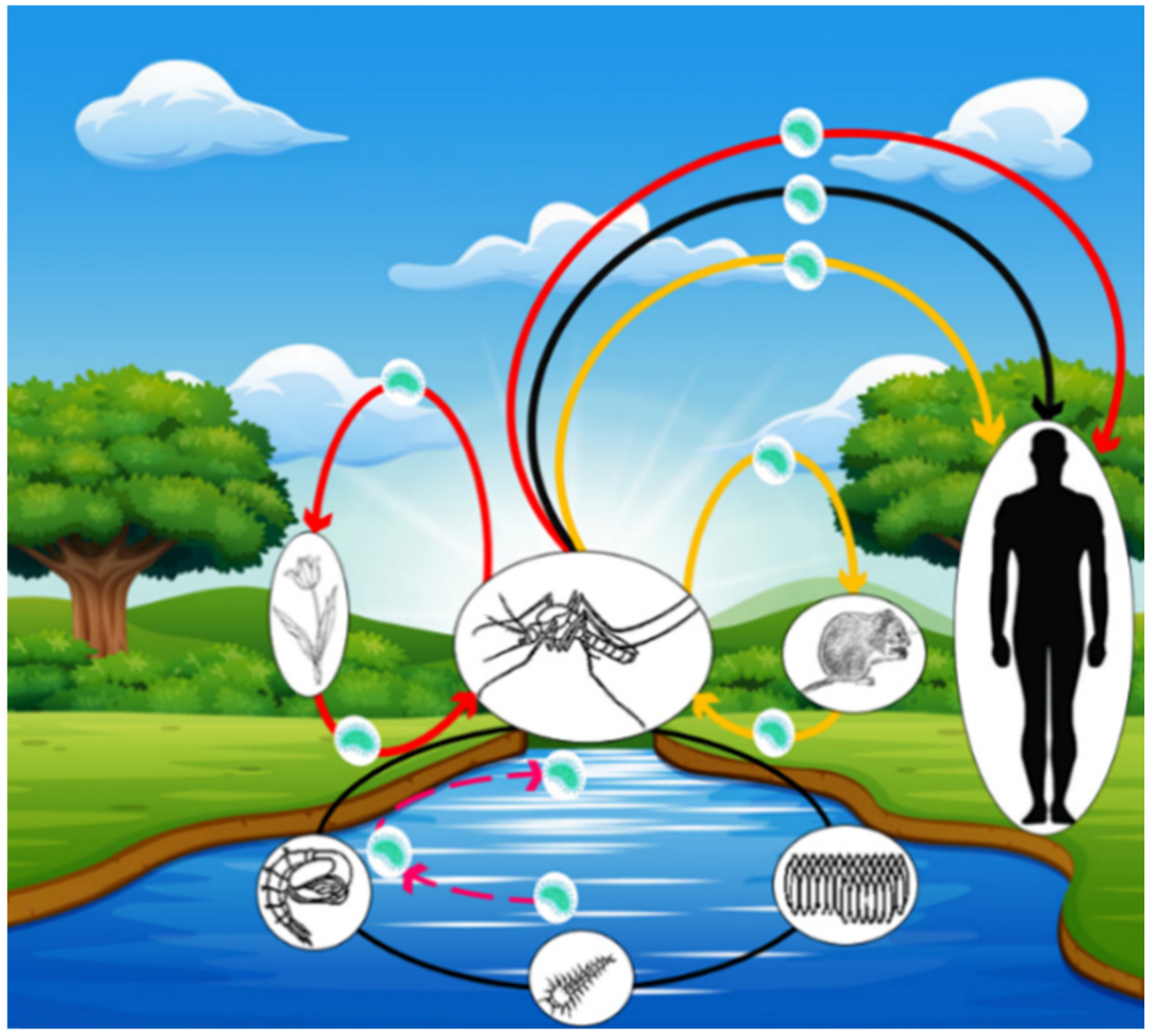
3.3.1. Possible Modes of Francisella tularensis Contamination for Mosquitoes
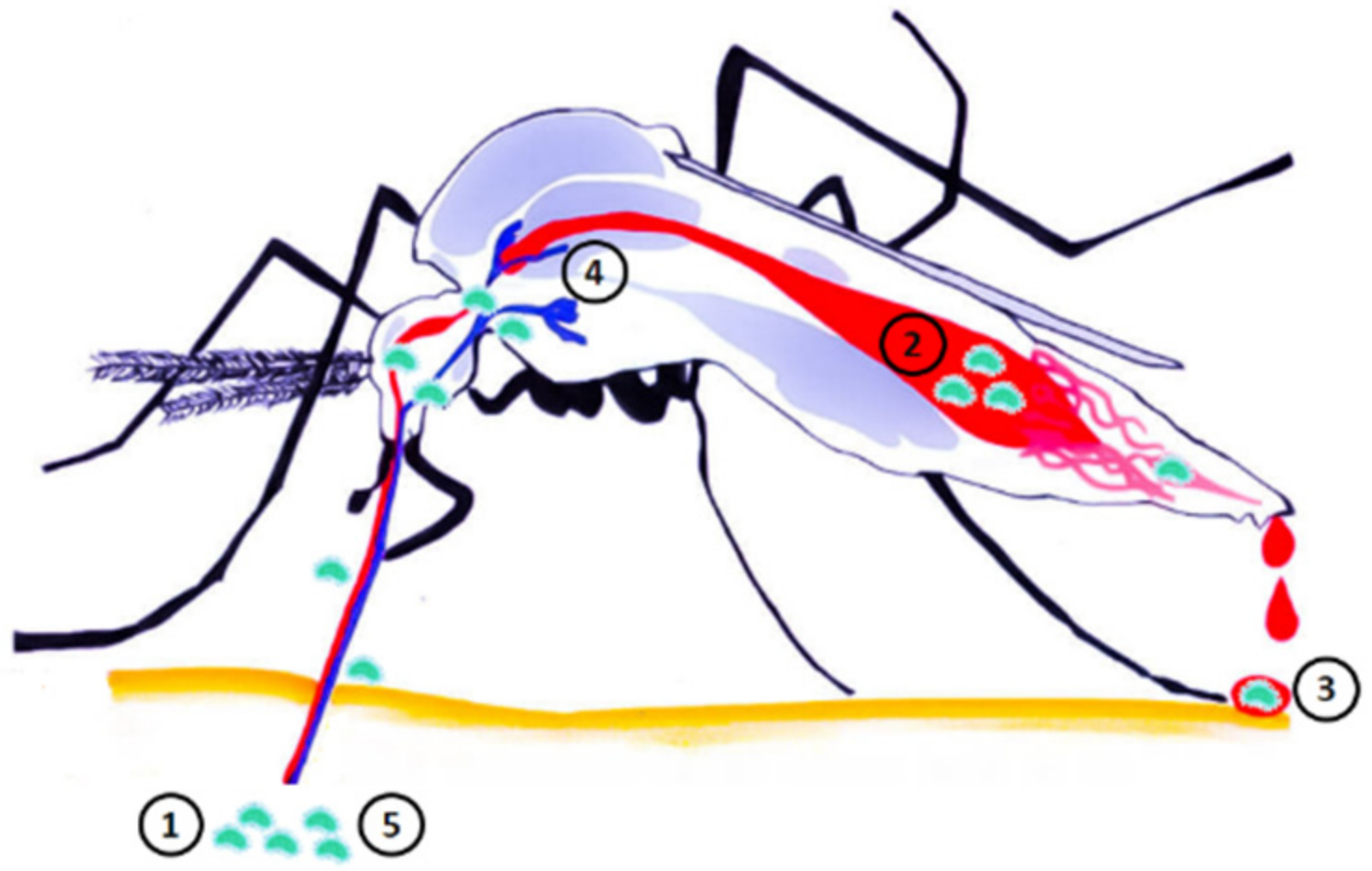
3.3.2. Possible Modes of Transmission of Francisella tularensis to Humans
4. Perspectives
4.1. Gaps to Confirm the Role of Mosquitoes as Vectors of Francisella tularensis
- -
- Does Fth multiply in the salivary glands of mosquitoes?
- -
- After ingestion, how does Fth resist the digestive enzymes of mosquitoes? Can it multiply in the mosquito’s digestive system and travel up to colonize the salivary glands?
- -
- What is the preferred microhabitat of Fth in mosquitoes?
- -
- What is the involvement of amoebae in the transmission of Fth to larval stages of mosquitoes?
- -
- What are the mechanisms used by Fth to survive through the various molts during the development of mosquito larval stages to adult?
- -
- Recently, phylogeography studies have shown several subpopulations of Fth around the word. In Scandinavia, Fth subpopulations B.Br.013/014, B.Br.012/013, B.Br.007/008, and B.Br.OSU18 are the most predominant [89]. Are there specific relationships between Fth subpopulations and mosquito species in Scandinavian areas?
- -
- What are the particularities of the mosquito species capable of transmitting Fth?
- -
- Are there mosquito species more adapted to transmit Fth than others?
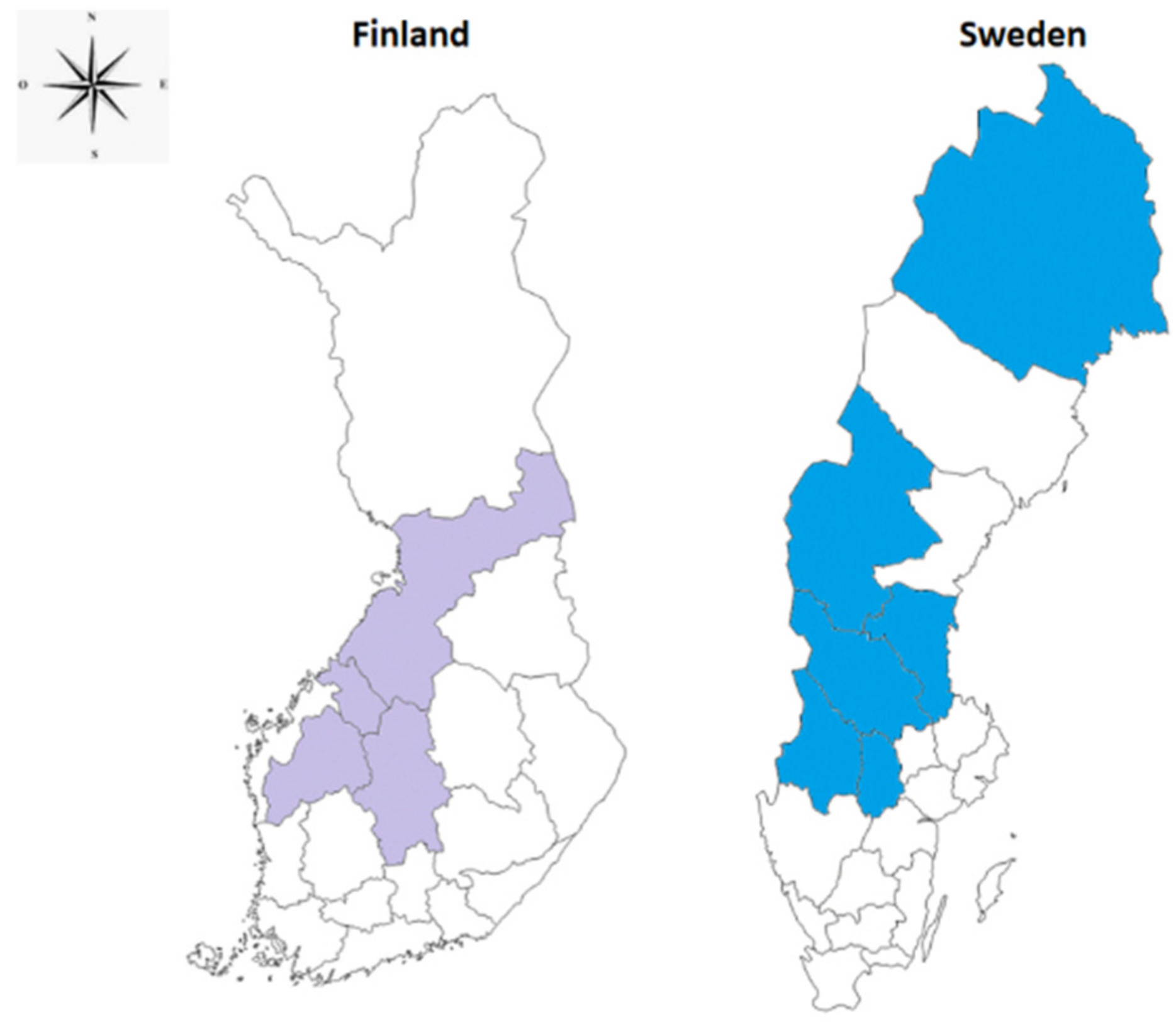
4.2. Why Transmission of Tularemia by Mosquitoes is Restricted to Finland and Sweden?
- -
- Climate and geology of Finland and Sweden could be factors favoring the association of Fth with mosquitoes?
- -
- Native mosquito species in Finland and Sweden might have undergone adaptations to the Fth, creating new vector-bacterium relationships
4.3. Transovarian Transmission
4.4. Mosquito Control and Tularemia
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McCoy, G.W. A plague-like disease of rodents. Public Health Bull. 1911, 45, 53–71. [Google Scholar]
- McCoy, G.W.; Chapin, C.W. Further observations on a plague-like disease of rodents with a preliminary note on the causative agent, Bacterium Tularense. J. Infect. Dis. 1912, 10, 61–72. [Google Scholar] [CrossRef]
- Sjostedt, A. Tularemia: History, epidemiology, pathogen physiology, and clinical manifestations. Ann. N. Y. Acad. Sci. 2007, 1105, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Francis, E. The occurrence of tularemia in nature as a disease of man. Public Health Rep. 1921, 36, 1731–1738. [Google Scholar] [CrossRef]
- Green, R.G. The occurrence of Bacterium tularense in the eastern wood tick Dermacentor variabilis. Am. J. Epidemiol. 1931, 14, 600–613. [Google Scholar] [CrossRef]
- Gürcan, S. Epidemiology of tularemia. Balkan. Med. J. 2014, 31, 3–10. [Google Scholar] [CrossRef]
- Sjöstedt, A.B.; Brenner, D.J.; Krieg, N.R.; Staley, J.T.; Garrity, G.M. Family XVII. Francisellaceae, genus I. Francisella. In Bergey’s Manual of Systematic Bacteriology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 200–210. [Google Scholar]
- Petersen, J.M.; Mead, P.S.; Schriefer, M.E. Francisella tularensis: An arthropod-borne pathogen. Vet. Res. 2009, 40, 7. [Google Scholar] [CrossRef] [Green Version]
- Keim, P.; Johansson, A.; Wagner, D.M. Molecular epidemiology, evolution, and ecology of Francisella. Ann. N.Y. Acad. Sci. 2007, 1105, 30–66. [Google Scholar] [CrossRef]
- Maurin, M.; Gyuranecz, M. Tularaemia: Clinical aspects in Europe. Lancet Infect. Dis. 2016, 16, 113–124. [Google Scholar] [CrossRef]
- Olsufjev, N.G.; Meshcheryakova, I.S. Infraspecific taxonomy of tularemia agent Francisella tularensis McCoy et Chapin. J. Hyg. Epidemiol. Microbiol. Immunol. 1982, 26, 291–299. [Google Scholar]
- Petersen, J.M.; Schriefer, M.E. Tularemia: Emergence/re-emergence. Vet. Res. 2005, 36, 455–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollis, D.G.; Weaver, R.E.; Steigerwalt, A.G.; Wenger, J.D.; Moss, C.W.; Brenner, D.J. Francisella philomiragia comb. nov. (formerly Yersinia philomiragia) and Francisella tularensis biogroup novicida (formerly Francisella novicida) associated with human disease. J. Clin. Microbiol. 1989, 27, 1601–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triebenbach, A.N.; Vogl, S.J.; Lotspeich-Cole, L.; Sikes, D.S.; Happ, G.M.; Hueffer, K. Detection of Francisella tularensis in Alaskan mosquitoes (Diptera: Culicidae) and assessment of a laboratory model for transmission. J. Med. Entomol. 2010, 47, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, D.T.; Inglesby, T.V.; Henderson, A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Friedlander, A.M.; Hauer, J.; Layton, M.; et al. Tularemia as a biological weapon: Medical and public health management. JAMA 2001, 285, 2763–2773. [Google Scholar] [CrossRef] [PubMed]
- Darling, R.G.; Catlett, C.L.; Huebner, K.D.; Jarrett, D.G. Threats in bioterrorism. I: CDC category a agents. Emerg. Med. Clin. N. Am. 2002, 20, 273–309. [Google Scholar] [CrossRef]
- Hennebique, A.; Boisset, S.; Maurin, M. Tularemia as a waterborne disease: A review. Emerg. Microbes. Infect. 2019, 8, 1027–1042. [Google Scholar] [CrossRef]
- Whipp, M.J.; Davis, J.M.; Lum, G.; De Boer, J.; Zhou, Y.; Bearden, S.W.; Petersen, J.M.; Chu, M.C.; Hogg, G. Characterization of a novicida-like subspecies of Francisella tularensis isolated in Australia. J. Med. Microbiol. 2003, 52, 839–842. [Google Scholar] [CrossRef]
- Grunow, R.; Kalaveshi, A.; Kuhn, A.; Mulliqi-Osmani, G.; Ramadani, N. Surveillance of tularaemia in Kosovo, 2001 to 2010. Euro. Surveill. 2012, 17, 20217. [Google Scholar] [CrossRef]
- Ulu-Kilic, A.; Doganay, M. An overview: Tularemia and travel medicine. Travel. Med. Infect. Dis. 2014, 12, 609–616. [Google Scholar] [CrossRef]
- Gürcan, Ş. Epidemiology of Francisella tularensis and tularemia disease. In Proceedings of the XXXV. Turkish Microbiology Congress, Kuşadası-Aydın, Turkey, 3 November 2012. [Google Scholar]
- Zargar, A.; Maurin, M.; Mostafavi, E. Tularemia, a re-emerging infectious disease in Iran and neighboring countries. Epidemiol. Health 2015, 37, e2015011. [Google Scholar] [CrossRef] [Green Version]
- Champion, M.D.; Zeng, Q.; Nix, E.B.; Nano, F.E.; Keim, P.; Kodira, C.D.; Borowsky, M.; Young, S.; Koehrsen, M.; Engels, R.; et al. Comparative genomic characterization of Francisella tularensis strains belonging to low and high virulence subspecies. PLoS. Pathog. 2009, 5, e1000459. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, M.J.; Simpson, A.J. Tularemia: Current diagnosis and treatment options. Expert. Rev. Anti. Infect. Ther. 2008, 6, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Francis, E. Deer-fly fever, or Pahvant valley plague. A disease of man of hitherto unknown etiology. Public Health Rep. 1919, 34, 2061–2062. [Google Scholar] [CrossRef]
- Klock, L.E.; Olsen, P.F.; Fukushima, T. Tularemia epidemic associated with the deerfly. JAMA 1973, 226, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Service, M.V. The Encyclopedia of Arthropod-transmitted Infections of Man and Domesticated Animals; CABI Publishing: Wallingford, UK, 2001. [Google Scholar]
- Harbach, R.E. The Culicidae (Diptera): A review of taxonomy, classification, and phylogeny. Zootaxa 2007, 1668, 591–638. [Google Scholar] [CrossRef] [Green Version]
- Harbach, R.E. Mosquito Taxonomic Inventory. 2018. Available online: http://mosquitotaxonomic-inventory.info/ (accessed on 29 July 2020).
- Reiter, P. Climate change and mosquito-borne disease. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar] [PubMed] [Green Version]
- Gratz, N. Vector and Rodent-Borne Diseases in Europe and North America: Distribution, Public Health Burden and Control; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Vinogradova, E.B. Culex Pipiens Pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control; Sofia-Moscow: Pensoft: Moscow, Russia, 2000. [Google Scholar]
- Shaikevich, E.V.; Vinogradova, E.B.; Bouattour, A.; Gouveia de Almeida, A.P. Genetic diversity of Culex pipiens mosquitoes in distinct populations from Europe: Contribution of Cx. quinquefasciatus in Mediterranean populations. Parasit. Vectors 2016, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; Van Bortel, W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 2015, 105, 637–663. [Google Scholar] [CrossRef]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W.A. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Robert, V.; Gunay, F.; Le Goff, G.; Philippe, B.; Șuleșco, T.; Khalin, A.; Medlock, J.; Kampen, H.; Petrić, D.; Schaffner, F. Distribution chart for Euro-Mediterranean mosquitoes (Western Palaearctic region). J. Eur. Mosq. Control Assoc. 2019, 37, 1–28. [Google Scholar]
- World Health Organization. World Malaria Report 2014; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Calzolari, M. Mosquito-borne diseases in Europe: An emerging public health threat. Rep. Parasitol. 2016, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thelaus, J.; Andersson, A.; Broman, T.; Backman, S.; Granberg, M.; Karlsson, L.; Kuoppa, K.; Larsson, E.; Lundmark, E.; Lundstrom, J.O.; et al. Francisella tularensis subspecies holarctica occurs in Swedish mosquitoes, persists through the developmental stages of laboratory-infected mosquitoes and is transmissible during blood feeding. Microb. Ecol. 2014, 67, 96–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Lu, G.; Li, J.; Kelly, P.; Li, M.; Wang, J.; Huang, K.; Qiu, H.; You, J.; Zhang, R.; et al. Molecular Detection of Rickettsia felis and Rickettsia bellii in Mosquitoes. Vector Borne Zoonotic Dis. 2019, 19, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Turell, M.J.; Knudson, G.B. Mechanical transmission of Bacillus anthracis by stable flies (Stomoxys calcitrans) and mosquitoes (Aedes aegypti and Aedes taeniorhynchus). Infect. Immun. 1987, 55, 1859–1861. [Google Scholar] [CrossRef] [Green Version]
- Kettle, D.S. Medical and Veterinary Entomology, 2nd ed.; CAB international: London, UK, 1995; 725p. [Google Scholar]
- Martinet, J.P.; Ferté, H.; Failloux, A.B.; Schaffner, F.; Depaquit, J. Mosquitoes of North-Western Europe as Potential Vectors of Arboviruses: A Review. Viruses 2019, 11, 1059. [Google Scholar] [CrossRef] [Green Version]
- Dauphin, G.; Zientara, S.; Zeller, H.; Murgue, B. West Nile: Worldwide current situation in animals and humans. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 343–355. [Google Scholar] [CrossRef]
- Arsevska, E.; Hellal, J.; Mejri, S.; Hammami, S.; Marianneau, P.; Calavas, D.; Hénaux, V. Identifying Areas Suitable for the Occurrence of Rift Valley Fever in North Africa: Implications for Surveillance. Transbound. Emerg. Dis. 2016, 63, 658–674. [Google Scholar] [CrossRef]
- Lo Presti, A.; Cella, E.; Angeletti, S.; Ciccozzi, M. Molecular epidemiology, evolution and phylogeny of Chikungunya virus: An updating review. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 41, 270–278. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, M.; Wang, H.; Liang, G. Japanese encephalitis and Japanese encephalitis virus in mainland China. Rev. Med. Virol. 2012, 22, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Chiles, R.E.; Martinez, V.M.; Fang, Y.; Green, E.N. Experimental infection of California birds with western equine encephalomyelitis and St. Louis encephalitis viruses. J. Med. Entomol. 2003, 40, 968–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Lymphatic Filariasis: A Handbook of Practical Entomology for National Lymphatic Filariasis Elimination Programmes; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Capelli, G.; Genchi, C.; Baneth, G.; Bourdeau, P.; Brianti, E.; Cardoso, L.; Danesi, P.; Fuehrer, H.P.; Giannelli, A.; Ionică, A.M.; et al. Recent advances on Dirofilaria repens in dogs and humans in Europe. Parasites Vectors 2018, 11, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossati, A.; Bargiacchi, O.; Kroumova, V.; Zaramella, M.; Caputo, A.; Garavelli, P.L. Climate, environment and transmission of malaria. Infez. Med. 2016, 24, 93–104. [Google Scholar]
- Rossow, H.; Ollgren, J.; Klemets, P.; Pietarinen, I.; Saikku, J.; Pekkanen, E.; Nikkari, S.; Syrjala, H.; Kuusi, M.; Nuorti, J.P. Risk factors for pneumonic and ulceroglandular tularaemia in Finland: A population-based case-control study. Epidemiol. Infect. 2014, 142, 2207–2216. [Google Scholar] [CrossRef]
- Desvars, A.; Furberg, M.; Hjertqvist, M.; Vidman, L.; Sjostedt, A.; Ryden, P.; Johansson, A. Epidemiology and ecology of tularemia in Sweden, 1984–2012. Emerg. Infect. Dis. 2015, 21, 32–39. [Google Scholar] [CrossRef]
- Rossow, H.; Ollgren, J.; Hytonen, J.; Rissanen, H.; Huitu, O.; Henttonen, H.; Kuusi, M.; Vapalahti, O. Incidence and seroprevalence of tularaemia in Finland, 1995 to 2013: Regional epidemics with cyclic pattern. Euro. Surveill. 2015, 20, 21209. [Google Scholar] [CrossRef] [Green Version]
- Desvars-Larrive, A.; Liu, X.; Hjertqvist, M.; Sjostedt, A.; Johansson, A.; Ryden, P. High-risk regions and outbreak modelling of tularemia in humans. Epidemiol. Infect. 2017, 145, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Philip, C.B. Experimental transmission of tularemia by mosquitoes. Public Health Rep. 1932, 47, 12. [Google Scholar] [CrossRef]
- Olin, G. The occurrence and mode of transmission of tularemia in Sweden. Acta Pathol. Microbiol. Scand. 1942, 19, 220–247. [Google Scholar] [CrossRef]
- Eliasson, H.; Lindback, J.; Nuorti, J.P.; Arneborn, M.; Giesecke, J.; Tegnell, A. The 2000 tularemia outbreak: A case-control study of risk factors in disease-endemic and emergent areas, Sweden. Emerg. Infect. Dis. 2002, 8, 956–960. [Google Scholar] [CrossRef] [PubMed]
- Hanke, C.A.; Otten, J.E.; Berner, R.; Serr, A.; Splettstoesser, W.; Von, S.C. Ulceroglandular tularemia in a toddler in Germany after a mosquito bite. Eur. J. Pediatr. 2009, 168, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Dryselius, R.; Hjertqvist, M.; Makitalo, S.; Dryselius, R.; Hjertqvist, M.; Mäkitalo, S.; Lindblom, A.; Lilja, T.; Eklöf, D.; Lindström, A. Large outbreak of tularaemia, central Sweden, July to September 2019. Euro. Surveill. 2019, 24, 1900603. [Google Scholar] [CrossRef] [PubMed]
- Oyston, P.C.; Sjostedt, A.; Titball, R.W. Tularaemia: Bioterrorism defense renews interest in Francisella tularensis. Nat. Rev. Microbiol. 2004, 2, 967–978. [Google Scholar] [CrossRef]
- Ryden, P.; Bjork, R.; Schafer, M.L.; Lundstrom, J.O.; Petersen, B.; Lindblom, A.; Forsman, M.; Sjostedt, A.; Johansson, A. Outbreaks of tularemia in a boreal forest region depends on mosquito prevalence. J. Infect. Dis. 2012, 205, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Hjertqvist, M.; Ahlm, C.; Klingstrom, J. Sex patterns in diagnoses of tularaemia, Sweden 1997–2008. J. Infect. 2010, 60, 186–187. [Google Scholar] [CrossRef]
- Christenson, B. An outbreak of tularemia in the northern part of central Sweden. Scand. J. Infect. Dis. 1984, 16, 285–290. [Google Scholar] [CrossRef]
- Uhari, M.; Syrjala, H.; Salminen, A. Tularemia in children caused by Francisella tularensis biovar palaearctica. Pediatr. Infect. Dis. J. 1990, 9, 80–83. [Google Scholar] [CrossRef]
- Eliasson, H.; Bäck, E. Tularaemia in an emergent area in Sweden: An analysis of 234 cases in five years. Scand. J. Infect. Dis. 2007, 39, 880–889. [Google Scholar] [CrossRef]
- Lundstrom, J.O.; Andersson, A.C.; Backman, S.; Schafer, M.L.; Forsman, M.; Thelaus, J. Transstadial transmission of Francisella tularensis holarctica in mosquitoes, Sweden. Emerg. Infect. Dis. 2011, 17, 794–799. [Google Scholar] [CrossRef]
- Jackson, J.; McGregor, A.; Cooley, L.; Ng, J.; Brown, M.; Ong, C.W.; Darcy, C.; Sintchenko, V. Francisella tularensis subspecies holarctica, Tasmania, Australia, 2011. Emerg. Infect. Dis. 2012, 18, 1484–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, M.L.; Lundstrom, J.O. The present distribution and predicted geographic expansion of the floodwater mosquito Aedes sticticus in Sweden. J. Vector. Ecol. 2009, 34, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, F.; Angel, G.; Geoffroy, B.; Rhaiem, A.; Hervy, J.P.; Brunhes, J. The Mosquitoes of Europe/Les moustiques d’Europe. Programme d’identification et d’enseignement; Montpellier, I.R.D., Mediterranee, E.I.D., Eds.; IRD Editions & EID Méditerranée: Marseille, France, 2001. [Google Scholar]
- Medlock, J.; Vaux, A. Aedes (Aedes) geminus Peus (Diptera: Culicidae)-An addition to the British mosquito fauna. Dipterists Digest 2009, 16, 147–150. [Google Scholar]
- Khalin, A.; Aibulatov, S. Fauna of Blood-Sucking Insects of the Gnus Complex in the Northwestern Region of Russia. III. Mosquitoes (Culicidae). Entomol. Rev. 2020, 100, 58–82. [Google Scholar] [CrossRef]
- Schaffner, F. Un nouveau moustique pour la faune française: Aedes (Ae.) esoensis rossicus Dolbeskin, Gorickaja & Mitrofanova, 1930 (Diptera, Culicidae). Bull. Soc. Entomol. Fr. 1995, 100, 325–330. [Google Scholar]
- Marshall, J.F. The British Mosquitoes. British Museum (Natural History); Johnson Reprint Corporation: London, UK, 1938. [Google Scholar]
- Service, M.W. Observations on feeding and oviposition in some British mosquitoes. Entomol. Exp. Appl. 1968, 11, 286–290. [Google Scholar] [CrossRef]
- Lilja, T.; Troell, K.; Kirik, H.; Lindström, A.A. distinct group of north European Aedes vexans as determined by mitochondrial and nuclear markers. Med. Vet. Entomol. 2018, 32, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Ludwig, H.W. Untersuchungen zur Faunistik und Ökologie der Stechmücken (Culicinae) und ihrer Pathogene im Oberrheingebiet. Mitt. Dtsch. Ges. Allgery Angew. Entomol. 1981, 2, 186–194. [Google Scholar]
- Merdic, E.; Lovakovic, T. Population dynamic of Aedes vexans and Aedes sticticus in flooded areas of the River Drava in Osijek, Croatia. J. Am. Mosq. Contr. Assoc. 2001, 17, 275–280. [Google Scholar]
- Minar, J.; Gelbic, I.; Olejnicek, J. The effect of floods on the development of mosquito populations in the middle and lower river Morava Regions. Acta. Universit. Carolinae. Biol. 2001, 45, 139–146. [Google Scholar]
- Schäfer, M.L.; Lundström, J.O.; Petersson, E. Comparison of mosquito (Diptera: Culicidae) populations by wetlands type and year in the lower River Dalälven region, Central Sweden. J. Vector Ecol. 2008, 33, 150–157. [Google Scholar] [CrossRef]
- Fine, P.E.M. Vectors and vertical transmission: An epidemiological perspective. Ann. N. Y. Acad. Sci. 1975, 266, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K. Epidemiology. In Arthropod-Borne Diseases; Marcondes, C.B., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 7–34. [Google Scholar]
- Mahajan, U.V.; Gravgaard, J.; Turnbull, M.; Jacobs, D.B.; McNealy, T.L. Larval exposure to Francisella tularensis LVS affects the fitness of the mosquito Culex quinquefasciatus. FEMS Microbiol. Ecol. 2011, 78, 520–530. [Google Scholar] [CrossRef]
- Backman, S.; Naslund, J.; Forsman, M.; Thelaus, J. Transmission of tularemia from a water source by transstadial maintenance in a mosquito vector. Sci. Rep. 2015, 5, 7793. [Google Scholar] [CrossRef] [PubMed]
- Kenney, A.; Cusick, A.; Payne, J.; Gaughenbaugh, A.; Renshaw, A.; Wright, J.; Seeber, R.; Barnes, R.; Florjanczyk, A.; Horzempa, J. The potential for flower nectar to allow mosquito to mosquito transmission of Francisella tularensis. PLoS ONE 2017, 12, e0175157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilo, P. Arthropod Infection Models for Francisella tularensis. Curr. Clin. Micro. Rpt. 2018, 5, 10–17. [Google Scholar] [CrossRef]
- Petersen, J.M.; Molins, C.R. Subpopulations of Francisella tularensis ssp. tularensis and holarctica: Identification and associated epidemiology. Future Microbiol. 2010, 5, 649–661. [Google Scholar]
- Mulla, M.S. Mosquito control then, now, and in the future. J. Am. Mosq. Control. Assoc. 1994, 10, 574–584. [Google Scholar]
- Word Health Organization. Handbook for Integrated Vector Management; WHO Press: Geneva, Switzerland, 2013. [Google Scholar]
- Niang, E.H.A.; Bassene, H.; Fenollar, F.; Mediannikov, O. Biological Control of Mosquito-Borne Diseases: The Potential of Wolbachia-Based Interventions in an IVM Framework. J. Trop. Med. 2018, 2018, 1470459. [Google Scholar] [CrossRef] [Green Version]
- Alout, H.; Labb’e, P.; Chandre, F.; Cohuet, A. Malaria Vector Control Still Matters despite Insecticide Resistance. Trends Parasitol. 2017, 33, 610–618. [Google Scholar] [CrossRef]
- Karunamoorthi, K.; Sabesan, S. Insecticide resistance in insect vectors of disease with special reference to mosquitoes: A potential threat to global public health. Health Scope 2013, 2, 4–18. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Human Pathogens | Vectors | References | |
|---|---|---|---|
| Viruses | Dengue Fever virus | Aedes | [44] |
| Zika virus | Aedes | [45] | |
| West Nile virus | Culex | [46] | |
| Rift Valley Fever virus | Aedes and Culex | [47] | |
| Chikungunya virus | Aedes | [48] | |
| Yellow Fever virus | Aedes | [44] | |
| Japanese Encephalitis virus | Culex | [49] | |
| Saint Louis Encephalitis virus | Culex | [50] | |
| Parasites | Wuchereria boncrofti | Aedes, Culex, and Anopheles | [51] |
| Brugia Malaya | Aedes, Culex, and Anopheles | [51] | |
| Dirofilaria immitis | Aedes, Culex, and Anopheles | [52] | |
| Dirofilaria repens | Aedes, Culex, and Anopheles | [52] | |
| Plasmodium knowlesi | Anopheles | [53] | |
| Plasmodium ovale | Anopheles | [53] | |
| Plasmodium vivax | Anopheles | [53] | |
| Plasmodium falciparum | Anopheles | [53] | |
| Plasmodium malariae | Anopheles | [53] | |
| Bacteria | Bacillus anthracis | Aedes and Culex | [43] |
| Francisella tularensis | Aedes and Culex | [41] | |
| Rickettsia felis | Cx. quinquefasciatus | [42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdellahoum, Z.; Maurin, M.; Bitam, I. Tularemia as a Mosquito-Borne Disease. Microorganisms 2021, 9, 26. https://doi.org/10.3390/microorganisms9010026
Abdellahoum Z, Maurin M, Bitam I. Tularemia as a Mosquito-Borne Disease. Microorganisms. 2021; 9(1):26. https://doi.org/10.3390/microorganisms9010026
Chicago/Turabian StyleAbdellahoum, Zakaria, Max Maurin, and Idir Bitam. 2021. "Tularemia as a Mosquito-Borne Disease" Microorganisms 9, no. 1: 26. https://doi.org/10.3390/microorganisms9010026