The Halotolerant Rhizobacterium—Pseudomonas koreensis MU2 Enhances Inorganic Silicon and Phosphorus Use Efficiency and Augments Salt Stress Tolerance in Soybean (Glycine max L.)

 ,
,  ,
,
Abstract
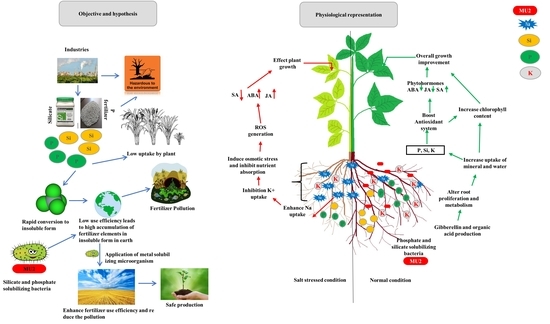
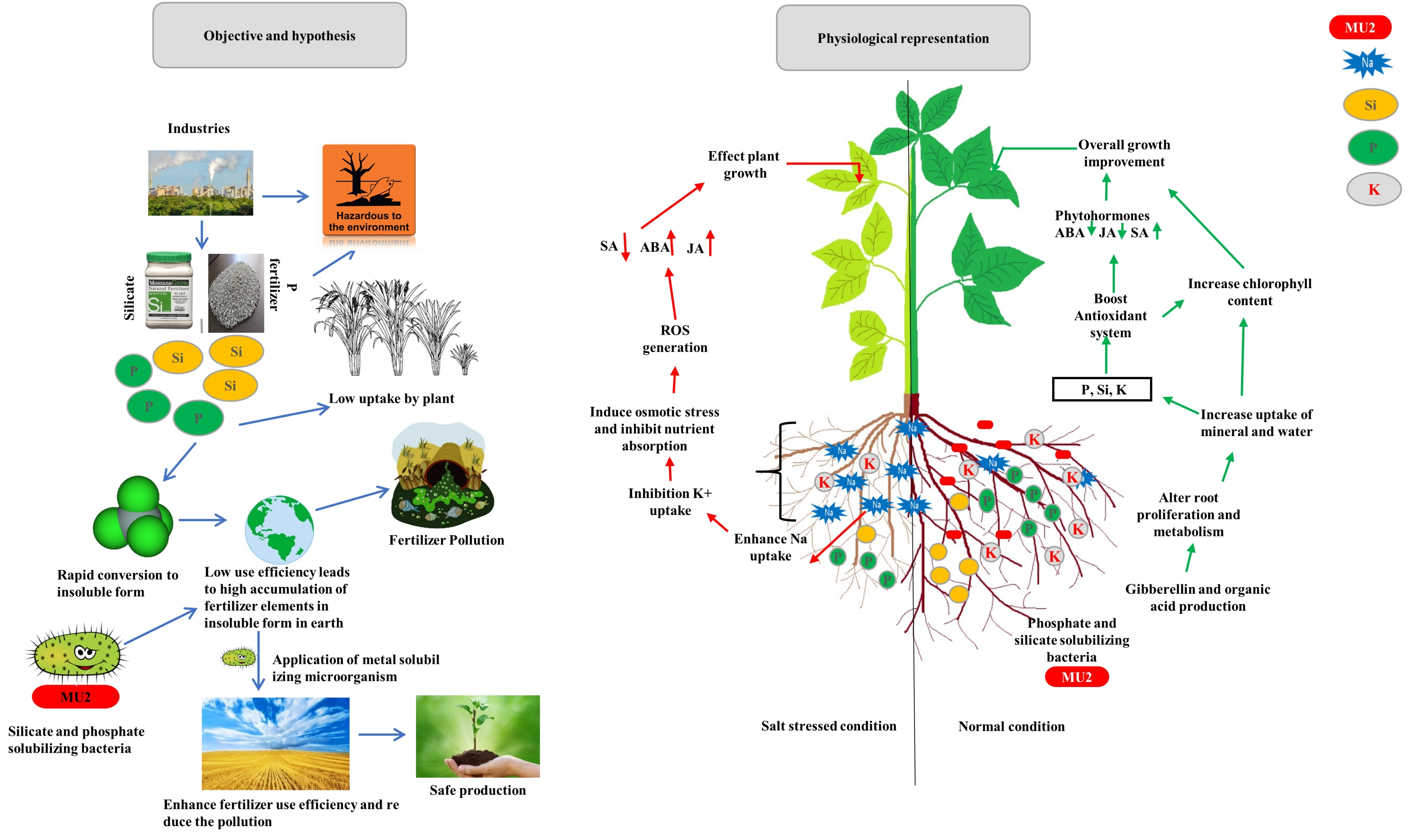
:
1. Introduction
2. Materials and Methods
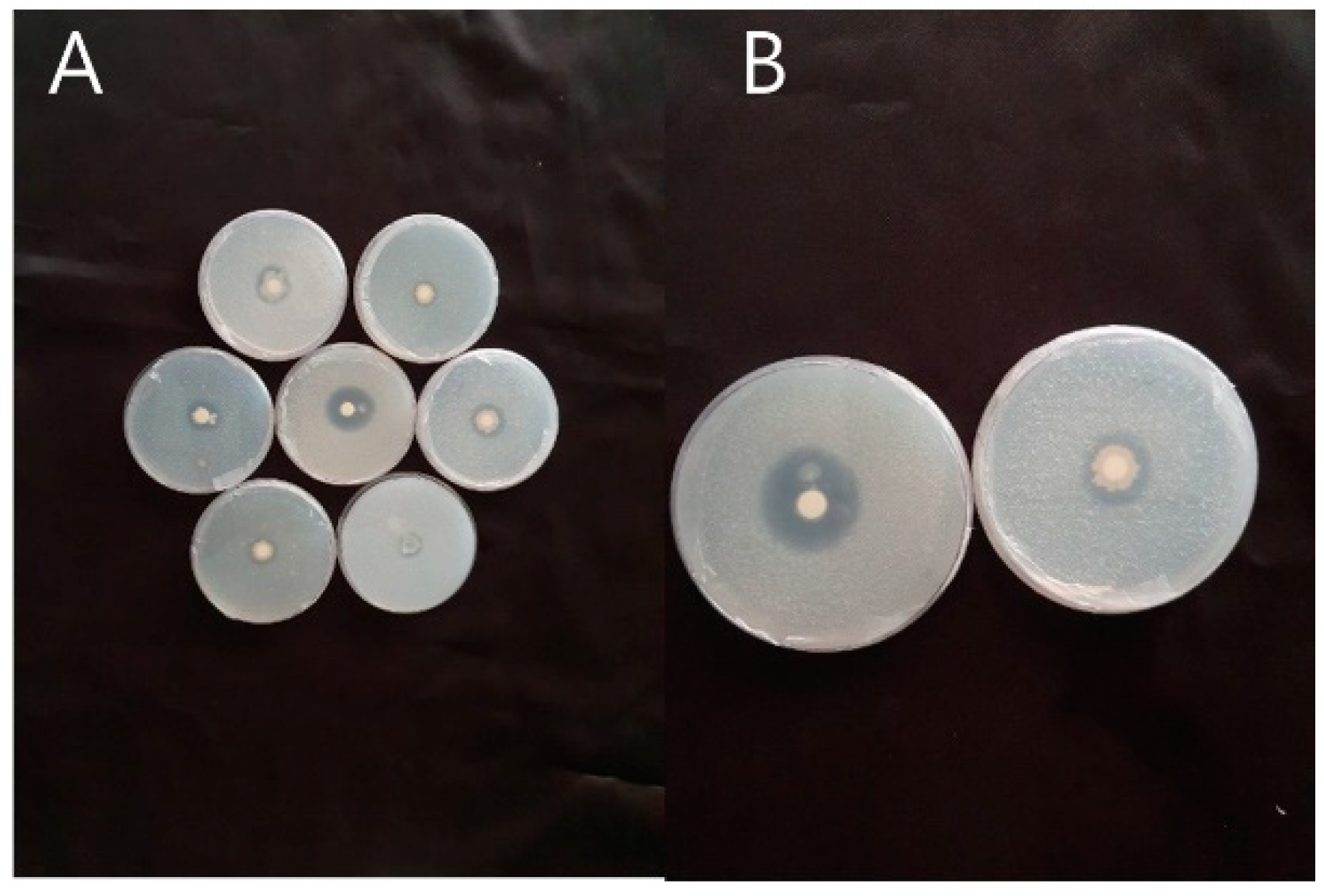
2.1. Microbe Selection
2.2. Quantification of Gibberellins and Organic Acid
2.3. Plant Materials and Experiment
2.3.1. Screening of Si and P Dose
2.3.2. Soil Preparation
2.3.3. Plant Experiment
2.4. Chlorophyll Measurement
2.5. Biochemical Analysis
2.5.1. Quantification of Si, P, Na, and K
2.5.2. RNA Isolation and qPCR Analysis
2.5.3. Quantification of Phytohormones
- ABA-The method described by Shahzad et al. [42] was used to determine ABA. In brief, 0.5 g of lyophilized ground samples was extracted with isopropanol and acetic acid (95:5 v/v). The solution obtained was filtered, and 1 mL of the internal ABA standard ((±)-3,5,5,7,7,7-d6)-ABA) was added. The extract was washed using 1 N NaOH, and the pH was adjusted to 12–13 followed by the removal of chlorophyll through CH2Cl2. The solution was separated by ethyl acetate, concentrated, and collected by washing using a phosphate buffer (pH 8.0). Polyvinylpolypyrrolidone was added, stirred for 30 min and filtered. The filtrate was evaporated and ABA was extracted through methanol/diethylether (2:3 v/v). ABA extracts were analyzed by GC-MS/SIM (6890 N Network GC System and 5973 Network Mass Selective Detector; Agilent Technologies, Santa Clara, CA, USA).
- JAs-The method described by Adhikari et al. [43] was used to determine the JA content of the plant. In brief, 0.1 g freeze-dried ground sample was extracted using acetone/citric acid (70:30 v/v) and filtered. The (9,10–2H2) Dihydro-JA (100 ng) was added as an internal standard. The solvent was evaporated, washed using a 0.1 m potassium phosphate buffer, and the pH was adjusted to 2.5. The solution was stirred using diethyl-amino cellulose, filtered, and washed with chloroform in a column fixed with sodium sulfate. The extract was dried, washed with diethyl ether and filtered with chloroform/isopropanol (2:1), and passed through diethyl ether/acetic acid (98:2) using an amino cartridge. After filtration, the extracts were dried and recovered with diethyl ether. Methylation was performed using diethyl ether followed by dichloromethane. The peaks were obtained by GC-MS/SIM (6890 N Network GC System and 5973 Network Mass Selective Detector; Agilent Technologies, Santa Clara, CA, USA).
- SA-SA was extracted and quantified using the method by Seskar et al. [23]. In brief, 0.5 g of lyophilized samples were extracted with 100% methanol, vacuum-dried, and suspended in trichloroacetic acid (5%). The solution was partitioned by ethyl acetate, cyclopentane, and isopropanol after which it was left to dry using N2. The excitation and emission peaks at 305 nm and 355 nm were recorded using HPLC (Shimadzu RF-10AXL fluorescence detector).
2.5.4. Antioxidant and Extent of Lipid Peroxidation Analysis
2.5.5. Proline Content
2.6. Statistical Analysis
3. Results
3.1. Quantification of Organic and Gibberellic Acids
3.2. Measurement of Morphological Parameters
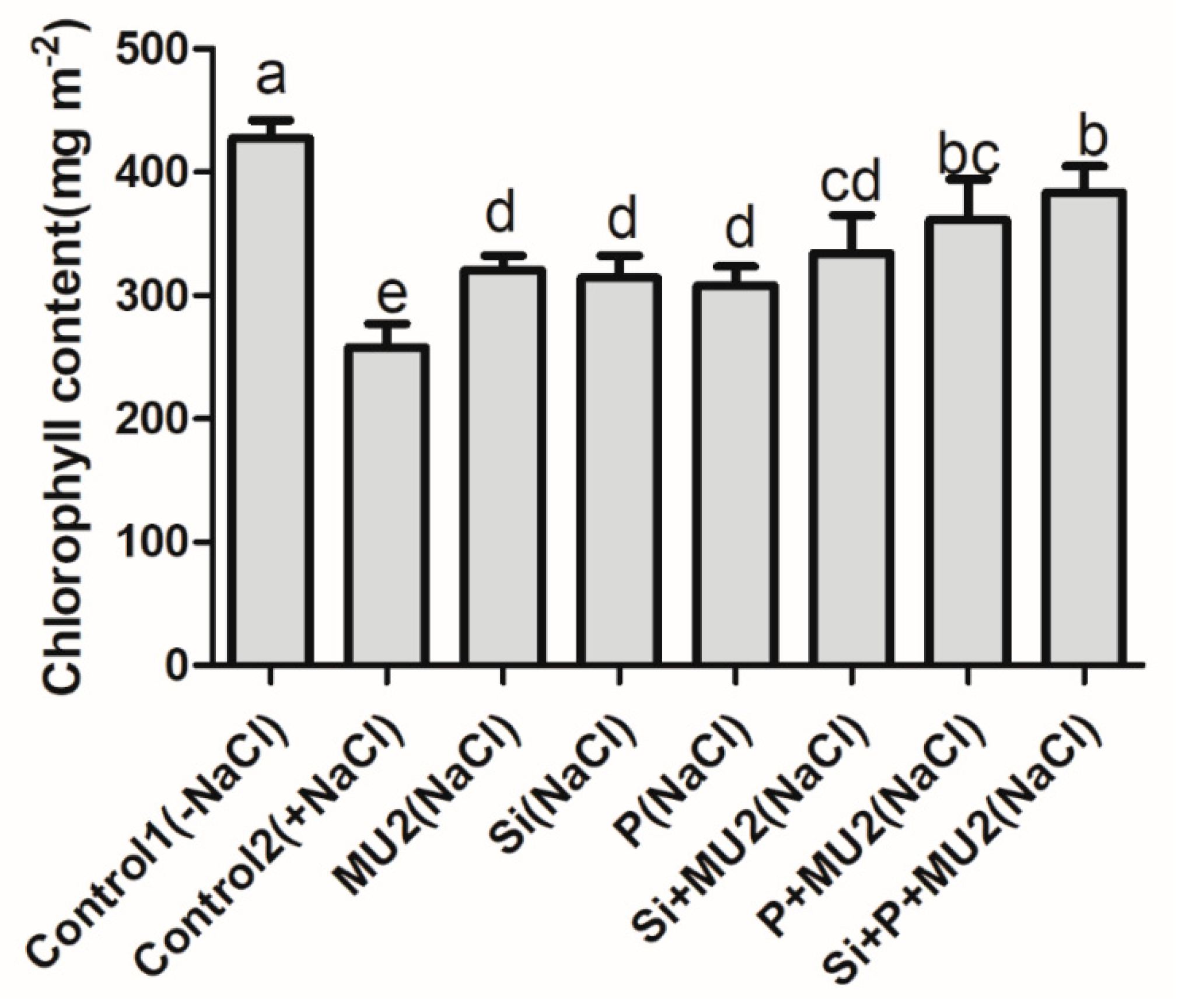
3.3. Chlorophyll Measurement
3.4. Analysis of Physiological Parameter
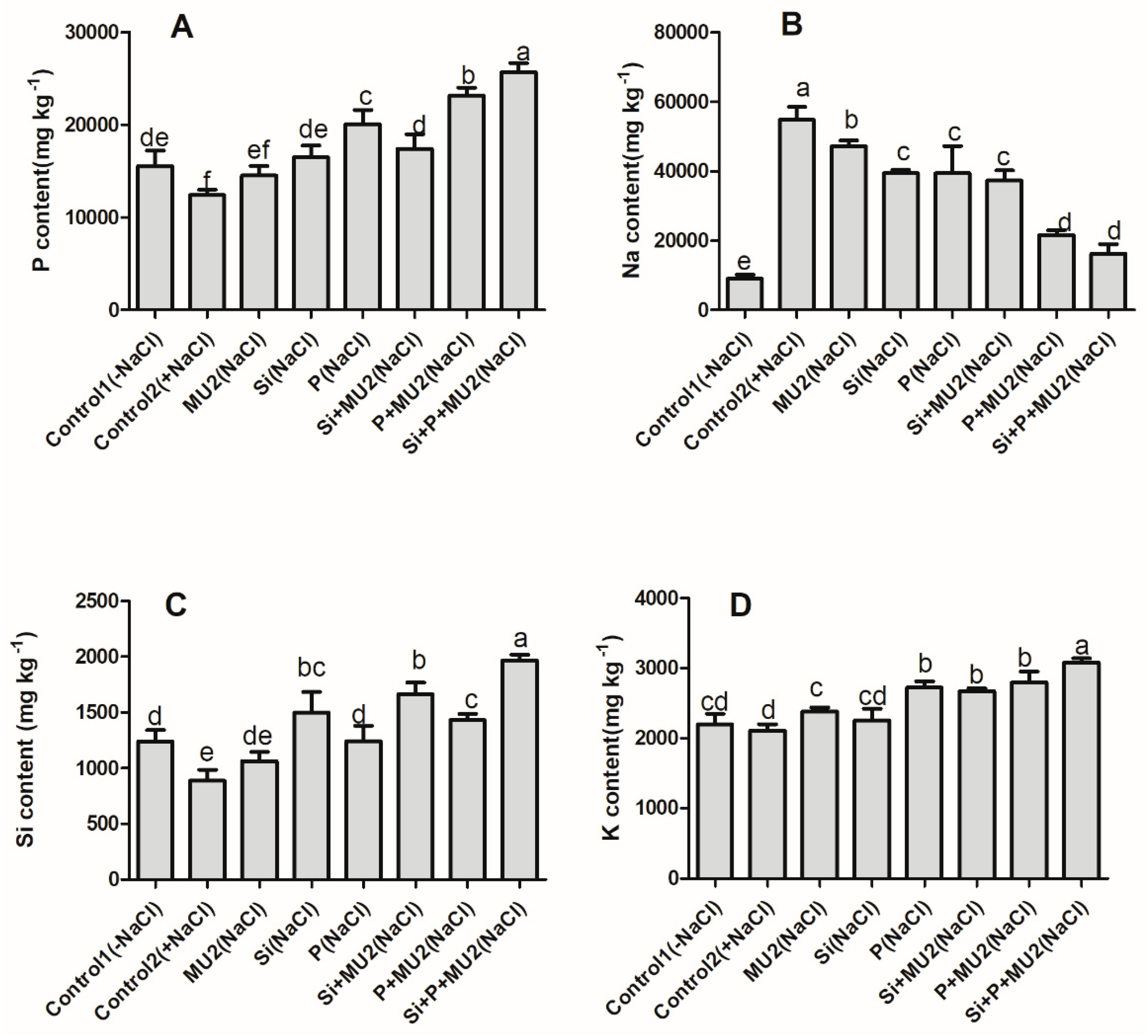
3.4.1. Quantification of Mineral Elements (Si, P, Na, and K)
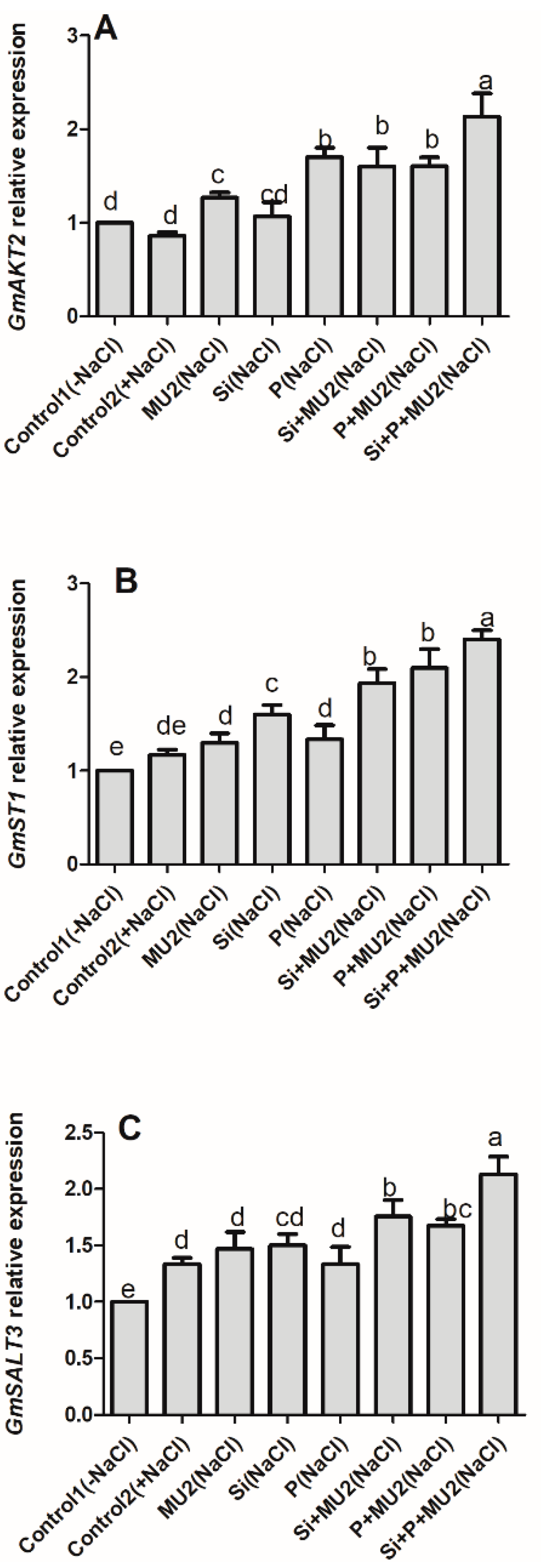
3.4.2. P. koreensis MU2 Modulates Stress-Resistant Gene in Soybean Leaves
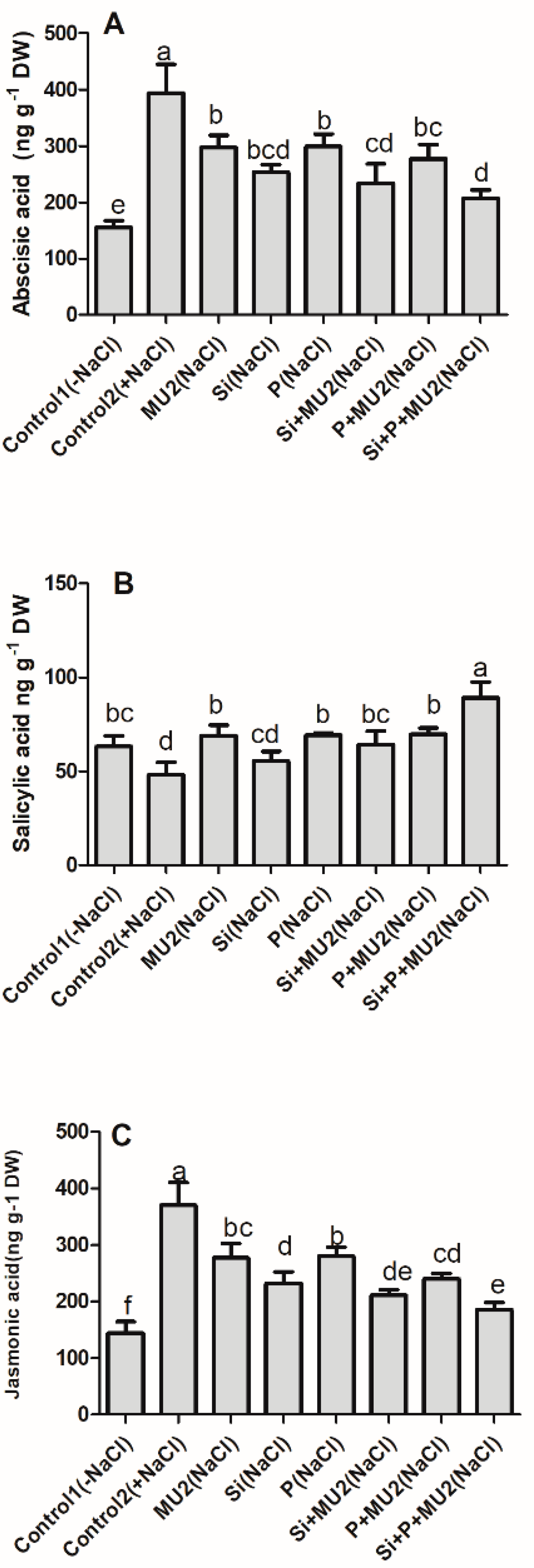
3.4.3. Effect on Phytohormones Level of Soybean Shoot
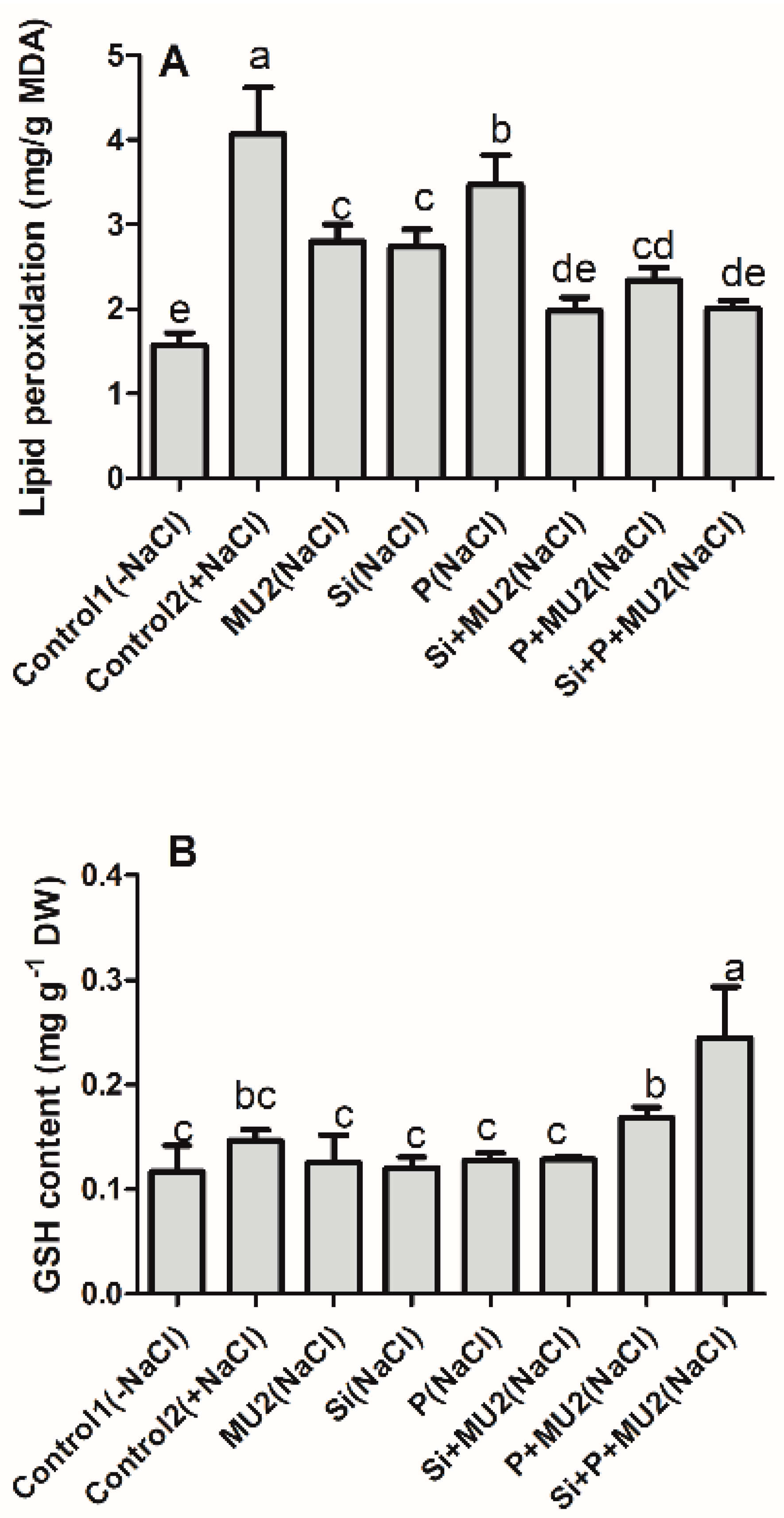
3.4.4. Effect on Plant Antioxidant System and Lipid Peroxidation under Stress Conditions
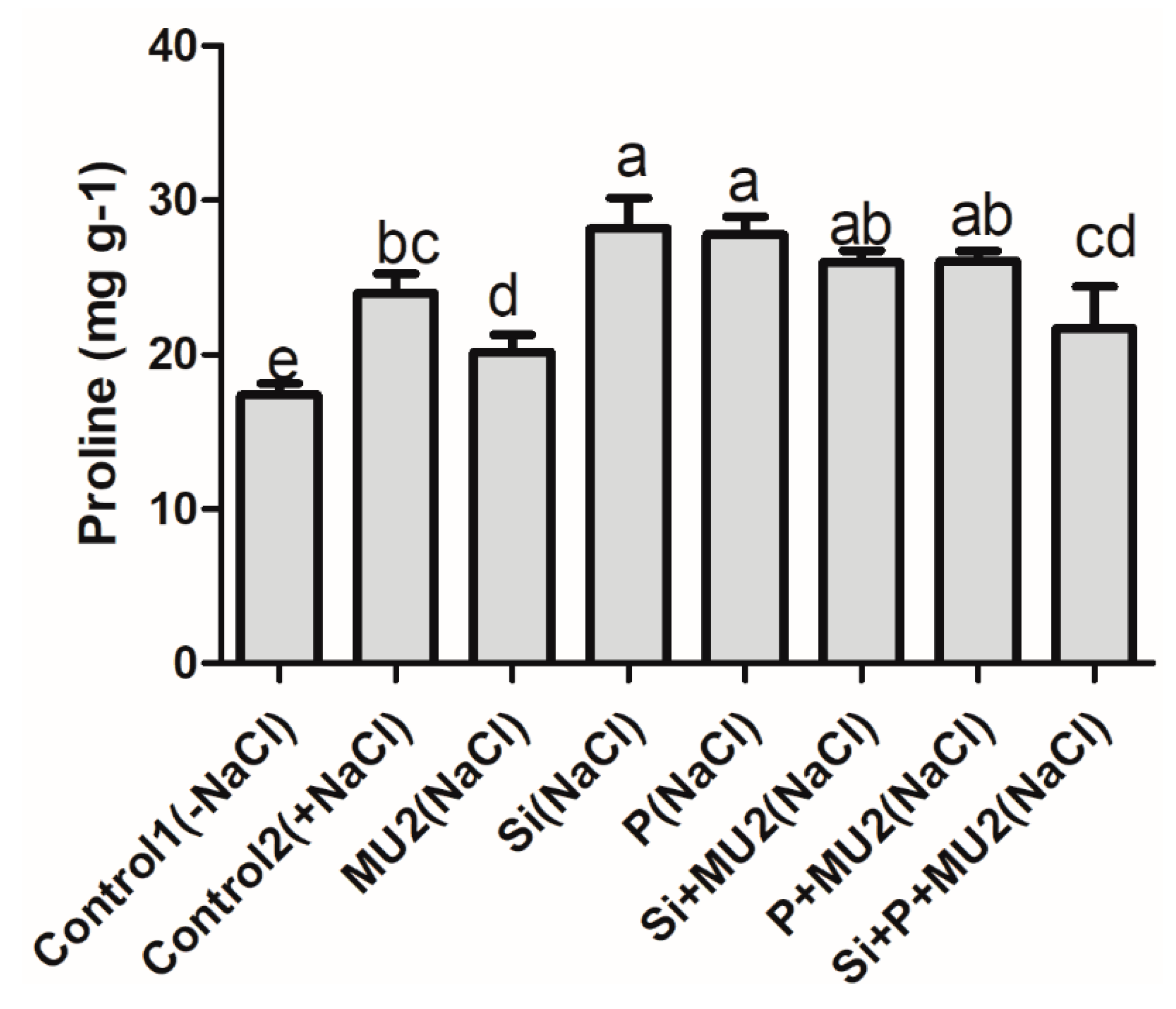
3.4.5. Proline Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Butcher, K.; Wick, A.F.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil salinity: A threat to global food security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Maji, D.; Chanotiya, C.S.; Kalra, A. ACC deaminase-containing Arthrobacter protophormiae induces NaCl stress tolerance through reduced ACC oxidase activity and ethylene production resulting in improved nodulation and mycorrhization in Pisum sativum. J. Plant Physiol. 2014, 171, 884–894. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D.L. Alleviating salt stress on soybean (Glycine max (L.) Merr.)—Bradyrhizobium japonicum symbiosis, using signal molecule genistein. Eur. J. Soil Biol. 2009, 45, 146–152. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Shao, H.; Qi, W.; Hamoud, Y.A.; Shaghaleh, H.; Khan, N.U.; Yang, R.; Tang, B. GABA-Alleviated oxidative injury induced by salinity, osmotic stress and their combination by regulating cellular and molecular signals in rice. Int. J. Mol. Sci. 2019, 20, 5709. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.M.; Lee, I.J. Plant growth-promoting endophytic bacteria augment growth and salinity tolerance in rice plants. Plant Biol. 2020. [Google Scholar] [CrossRef]
- Gengmao, Z.; Shihui, L.; Xing, S.; Yizhou, W.; Zipan, C. The role of silicon in physiology of the medicinal plant (Lonicera japonica L.) under salt stress. Sci. Rep. 2015, 5, 12696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Guo, F.; Cornelis, J.-T.; Song, Z.; Wang, X.; Delvaux, B.J.F.i.P.S. Combined silicon-phosphorus fertilization affects the biomass and phytolith stock of rice plants. Front. Plant. Sci. 2020, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargaz, A.; Nassar, R.; Rady, M.; Gaballah, M.; Thompson, S.; Brestic, M.; Schmidhalter, U.; Abdelhamid, M.T. Improved salinity tolerance by phosphorus fertilizer in two Phaseolus vulgaris recombinant inbred lines contrasting in their P-efficiency. J. Agron. Crop Sci. 2016, 202, 497–507. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Bakhat, H.F.; Bibi, N.; Zia, Z.; Abbas, S.; Hammad, H.M.; Fahad, S.; Ashraf, M.R.; Shah, G.M.; Rabbani, F.; Saeed, S. Silicon mitigates biotic stresses in crop plants: A review. Crop Prot. 2018, 104, 21–34. [Google Scholar] [CrossRef]
- Vaculík, M.; Lux, A.; Luxová, M.; Tanimoto, E.; Lichtscheidl, I. Silicon mitigates cadmium inhibitory effects in young maize plants. Environ. Exp. Bot. 2009, 67, 52–58. [Google Scholar] [CrossRef]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Muneer, S.; Park, Y.G.; Kim, S.; Jeong, B.R. Foliar or subirrigation silicon supply mitigates high temperature stress in strawberry by maintaining photosynthetic and stress-responsive proteins. J. Plant Growth Regul. 2017, 36, 836–845. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum ‘Bugwang’. J. Biomed. Biotechnol. 2016, 2016, 4. [Google Scholar]
- Tounkara, A.; Clermont-Dauphin, C.; Affholder, F.; Ndiaye, S.; Masse, D.; Cournac, L. Inorganic fertilizer use efficiency of millet crop increased with organic fertilizer application in rainfed agriculture on smallholdings in central Senegal. Agric. Ecosyst. Environ. 2020, 294, 106878. [Google Scholar] [CrossRef]
- Tittonell, P.; Giller, K.E. When yield gaps are poverty traps: The paradigm of ecological intensification in African smallholder agriculture. Field Crop. Res. 2013, 143, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Fofana, B.; Wopereis, M.; Bationo, A.; Breman, H.; Mando, A. Millet nutrient use efficiency as affected by natural soil fertility, mineral fertilizer use and rainfall in the West African Sahel. Nutr. Cycl. Agroecosyst. 2008, 81, 25–36. [Google Scholar] [CrossRef]
- Tubaña, B.S.; Heckman, J.R. Silicon in soils and plants. In Silicon and Plant Diseases; Springer: Berlin/Heidelberg, Germany, 2015; pp. 7–51. [Google Scholar]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Functions and transport of silicon in plants. Cell Mol. Life Sci. 2008, 65, 3049–3057. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations: 2017. World Fertilizer Trends and Outlook to 2020. Available online: http://www.fao.org/3/a-i6895e.pdf (accessed on 19 August 2020).
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.-M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Pande, A.; Pandey, P.; Mehra, S.; Singh, M.; Kaushik, S. Biotechnology. Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. J. Genet. Eng. Biotechnol. 2017, 15, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Shah, Z.; Fahad, S.; Arif, M.; Alam, M.; Khan, I.A.; Mian, I.A.; Basir, A.; Ullah, H.; Arshad, M.; et al. Phosphate-solubilizing bacteria nullify the antagonistic effect of soil calcification on bioavailability of phosphorus in alkaline soils. Sci. Rep. 2017, 7, 16131. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S.; Rizvi, A.; Saif, S.; Ahmad, B.; Shahid, M. Role of phosphate-solubilizing bacteria in legume improvement. In Microbes for Legume Improvement; Springer: Berlin/Heidelberg, Germany, 2017; pp. 175–197. [Google Scholar]
- Bano, A.; Fatima, M. Salt tolerance in Zea mays (L). following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Joe, M.M.; Devaraj, S.; Benson, A.; Sa, T. Isolation of phosphate solubilizing endophytic bacteria from Phyllanthus amarus Schum & Thonn: Evaluation of plant growth promotion and antioxidant activity under salt stress. J. Appl. Res. Med. Aromat. Plants 2016, 3, 71–77. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Asaf, S.; Khan, M.A.; Kang, S.-M.; Yun, B.-W.; Lee, I.-J. Biochemistry. Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol. Biochem. 2016, 106, 236–243. [Google Scholar] [CrossRef]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Lee, I.-J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef]
- Qadir, M.; Hussain, A.; Hamayun, M.; Shah, M.; Iqbal, A.; Husna; Murad, W. Phytohormones producing rhizobacterium alleviates chromium toxicity in Helianthus annuus L. by reducing chromate uptake and strengthening antioxidant system. Chemosphere 2020, 258, 127386. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.-M.; Lee, I.-J. Halotolerant rhizobacterial strains mitigate the adverse effects of NaCl stress in soybean seedlings. BioMed Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H. Can interaction between silicon and plant growth promoting rhizobacteria benefit in alleviating abiotic and biotic stresses in crop plants? Agric. Ecosyst. Environ. 2018, 253, 98–112. [Google Scholar] [CrossRef]
- Pathan, M.S.; Lee, J.-D.; Shannon, J.G.; Nguyen, H.T. Recent advances in breeding for drought and salt stress tolerance in soybean. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Springer: Berlin/Heidelberg, Germany, 2007; pp. 739–773. [Google Scholar]
- Aghaei, K.; Ehsanpour, A.A.; Shah, A.; Komatsu, S. Proteome analysis of soybean hypocotyl and root under salt stress. Amino Acids 2009, 36, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, J.-H.; Lee, I.-J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Lee, K.-E.; Radhakrishnan, R.; Kang, S.-M.; You, Y.-H.; Joo, G.-J.; Lee, I.-J.; Ko, J.-H.; Kim, J.-H. Enterococcus faecium LKE12 cell-free extract accelerates host plant growth via gibberellin and indole-3-acetic acid secretion. J. Microbiol. Biotechnol. 2015, 25, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-E.; Adhikari, A.; Kang, S.-M.; You, Y.-H.; Joo, G.-J.; Kim, J.-H.; Kim, S.-J.; Lee, I.-J. Isolation and characterization of the high silicate and phosphate solubilizing novel strain Enterobacter ludwigii GAK2 that promotes growth in rice plants. Agronomy 2019, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Bilal, S.; Khan, A.L.; Shahzad, R.; Asaf, S.; Kang, S.-M.; Lee, I.-J. Endophytic Paecilomyces formosus LHL10 Augments Glycine max L. Adaptation to Ni-Contamination through Affecting Endogenous Phytohormones and Oxidative Stress. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Chan, C.-X.; Teo, S.-S.; Ho, C.-L.; Othman, R.Y.; Phang, S.-M. Optimisation of RNA extraction from Gracilaria changii (Gracilariales, Rhodophyta). J. Appl. Phycol. 2004, 16, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Hamayun, M.; Kang, S.-M.; Lee, I.-J. Foliar application of methyl jasmonate induced physio-hormonal changes in Pisum sativum under diverse temperature regimes. Plant Physiol. Biochem. 2015, 96, 406–416. [Google Scholar] [CrossRef]
- Adhikari, A.; Lee, K.-E.; Khan, M.A.; Kang, S.-M.; Adhikari, B.; Imran, M.; Jan, R.; Kim, K.-M.; Lee, I.-J. Effect of silicate and phosphate solubilizing rhizobacterium Enterobacter ludwigii GAK2 on Oryza sativa L. under Cadmium Stress. J. Microbiol. Biotechnol. 2020, 30, 118–126. [Google Scholar] [CrossRef]
- Adhikari, B.; Dhungana, S.K.; Ali, M.W.; Adhikari, A.; Kim, I.-D.; Shin, D.-H. Resveratrol, total phenolic and flavonoid contents, and antioxidant potential of seeds and sprouts of Korean peanuts. Food Sci. Biotechnol. 2018, 27, 1275–1284. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Rhizobacteria AK1 remediates the toxic effects of salinity stress via regulation of endogenous phytohormones and gene expression in soybean. Biochem. J. 2019, 476, 2393–2409. [Google Scholar] [CrossRef] [PubMed]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, R.I.; Egamberdieva, D. Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.M.; Adhikari, A.; Lee, K.E.; Khan, M.A.; Khan, A.L.; Shahzad, R.; Dhungana, S.K.; Lee, I.J. Inoculation with Indole-3-acetic acid-producing rhizospheric Rhodobacter sphaeroides KE149 augments growth of adzuki bean plants under water stress. J. Microbiol. Biotechnol. 2020, 30, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Lütke, S.F.; Oliveira, M.L.; Silva, L.F.; Cadaval, T.R., Jr.; Dotto, G.L. Nanominerals assemblages and hazardous elements assessment in phosphogypsum from an abandoned phosphate fertilizer industry. Chemosphere 2020, 256, 127138. [Google Scholar] [CrossRef]
- Solgi, E.; Sheikhzadeh, H.; Solgi, M. Role of irrigation water, inorganic and organic fertilizers in soil and crop contamination by potentially hazardous elements in intensive farming systems: Case study from Moghan agro-industry, Iran. J. Geochem. Explor. 2018, 185, 74–80. [Google Scholar] [CrossRef]
- Hussain, S.; Shuxian, L.; Mumtaz, M.; Shafiq, I.; Iqbal, N.; Brestic, M.; Shoaib, M.; Sisi, Q.; Li, W.; Mei, X.; et al. Foliar application of silicon improves stem strength under low light stress by regulating lignin biosynthesis genes in soybean (Glycine max (L.) Merr.). J. Hazard. Mater. 2020, 401, 123256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, Y.; Zhao, X.; Jin, X.; Hou, L.; Shi, Y.; Ahammed, G.J. Silicon compensates phosphorus deficit-induced growth inhibition by improving photosynthetic capacity, antioxidant potential, and nutrient homeostasis in tomato. Agronomy 2019, 9, 733. [Google Scholar] [CrossRef] [Green Version]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Patel, P.R.; Kajal, S.S.; Patel, V.R.; Patel, V.J.; Khristi, S.M. Impact of saline water stress on nutrient uptake and growth of cowpea. Braz. J. Plant Physiol. 2010, 22, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; Ahmed, A.-H. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Kang, S.-M.; Khan, A.L.; Hamayun, M.; Hussain, J.; Joo, G.-J.; You, Y.-H.; Kim, J.-G.; Lee, I.-J. Gibberellin-producing Promicromonospora sp. SE188 improves Solanum lycopersicum plant growth and influences endogenous plant hormones. J. Microbiol. 2012, 50, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Khan, A.R.; Jung, B.K.; Khan, A.L.; Lee, I.-J.; Shin, J.-H. Gibberellins synthesized by the entomopathogenic bacterium, Photorhabdus temperata M1021 as one of the factors of rice plant growth promotion. J. Plant Interact. 2014, 9, 775–782. [Google Scholar] [CrossRef]
- Kang, S.-M.; Khan, A.L.; Waqas, M.; You, Y.-H.; Hamayun, M.; Joo, G.-J.; Shahzad, R.; Choi, K.-S.; Lee, I.-J. Gibberellin-producing Serratia nematodiphila PEJ1011 ameliorates low temperature stress in Capsicum annuum L. Eur. J. Soil Biol. 2015, 68, 85–93. [Google Scholar] [CrossRef]
- Sgroy, V.; Cassán, F.; Masciarelli, O.; Del Papa, M.F.; Lagares, A.; Luna, M.V. Isolation and characterization of endophytic plant growth-promoting (PGPB) or stress homeostasis-regulating (PSHB) bacteria associated to the halophyte Prosopis strombulifera. Appl. Microbiol. Biotechnol. 2009, 85, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Stella, A.; Storni, M.; Zulpa, G.; Zaccaro, M. Effects of cyanobacterial extracellular products and gibberellic acid on salinity tolerance in Oryza sativa L. Saline Syst. 2006, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baset, M.; Shamsuddin, Z.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured’musa’plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85. [Google Scholar]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef]
- Othman, R.; Panhwar, Q.A. Phosphate-solubilizing bacteria improves nutrient uptake in aerobic rice. In Phosphate Solubilizing Microorganisms; Springer: Berlin/Heidelberg, Germany, 2014; pp. 207–224. [Google Scholar]
- Iqbal, U.; Jamil, N.; Ali, I.; Hasnain, S. Effect of zinc-phosphate-solubilizing bacterial isolates on growth of Vigna radiata. Ann. Microbiol. 2010, 60, 243–248. [Google Scholar] [CrossRef]
- Kang, S.-M.; Waqas, M.; Shahzad, R.; You, Y.-H.; Asaf, S.; Khan, M.A.; Lee, K.-E.; Joo, G.-J.; Kim, S.-J.; Lee, I.-J.; et al. Isolation and characterization of a novel silicate-solubilizing bacterial strain Burkholderia eburnea CS4-2 that promotes growth of japonica rice (Oryza sativa L. cv. Dongjin). Soil Sci. Plant Nutr. 2017, 63, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Çimrin, K.M.; Türkmen, Ö.; Turan, M.; Tuncer, B. Phosphorus and humic acid application alleviate salinity stress of pepper seedling. Afr. J. Biotechnol. 2010, 9, 5845–5851. [Google Scholar]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-H.; Khan, A.L.; Lee, I.-J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Sopory, S.K. Chemical signaling under abiotic stress environment in plants. Plant Signal. Behav. 2008, 3, 525–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Thermotolerance effect of plant growth-promoting Bacillus cereus SA1 on soybean during heat stress. BMC Microbiol. 2020, 20, 175. [Google Scholar]
- Kang, S.-M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.-J.; Park, J.-M.; Kim, B.-R.; Shin, D.-H.; Lee, I.-J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Bilal, S.; Khan, A.L.; Shahzad, R.; Kim, Y.-H.; Imran, M.; Khan, M.J.; Al-Harrasi, A.; Kim, T.H.; Lee, I.-J. Mechanisms of Cr(VI) resistance by endophytic Sphingomonas sp. LK11 and its Cr(VI) phytotoxic mitigating effects in soybean (Glycine max L.). Ecotoxicol. Environ. Saf. 2018, 164, 648–658. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Cross-talk between nitric oxide, hydrogen peroxide and calcium in salt-stressed Chenopodium quinoa Willd. At seed germination stage. Plant Physiol. Biochem. 2020, 154, 657–664. [Google Scholar] [CrossRef]
- Yan, K.; Shao, H.; Shao, C.; Chen, P.; Zhao, S.; Brestic, M.; Chen, X. Physiological adaptive mechanisms of plants grown in saline soil and implications for sustainable saline agriculture in coastal zone. Acta Physiol. Plant. 2013, 35, 2867–2878. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Aldubise, A.; Egamberdieva, D. Arbuscular mycorrhizal fungi enhances salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. J. Plant Interact. 2015, 10, 230–242. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2019, 159, 55–65. [Google Scholar] [CrossRef]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GmST1 | 5′TCTAGAATGGCGTTTGTTGCAGCCATG3′ | 5′GAGCTCTCATAAGGTTCGGGGATCCTTTC3′ |
| GmAKT2 | 5′CACCATGCTAGCTGATCGTTACC3′ | 5′TCAGCTTATCCAACAAAAAAAAAGT3′ |
| GmSALT3 | 5′ATGACGTTCAACGCGAGC3′ | 5′AAGTTCTTCGATAGCATCTTTA3′ |
| Actin | 5′ATGGTGGGTATGGGTCAAAA3′ | 5′GAGGACAGGATGCTCCTCA3′ |
| Organic Acid Content (ppm) | Malic Acid | Citric Acid | Tartaric Acid | Acetic Acid |
|---|---|---|---|---|
| Control | 169.38 ± 12.36 b | 200 ± 10.9 b | 186.37 ± 15.88 a | 95.33 ± 12.66 b |
| NaCl | 220.63 ± 10.82 a | 237.8 ± 7.12 a | 219.40 ± 13.21 a | 189.66 ± 21.59 a |
| GA Content (ng/100 mL) | GA1 | GA3 |
| Control | 6.76 ± 0.77a | 11.9 ± 1.07a |
| NaCl | 3.77 ± 0.40b | 7.60 ± 0.92b |
| Treatments | Root Length | Shoot Length | Root Wt. | Shoot Wt. |
|---|---|---|---|---|
| Control 1(-NaCl) | 31 ± 1a | 39.6 ± 0.62a | 8.74 ± 0.36a | 16.78 ± 0.61a |
| Control 2(+NaCl) | 18 ± 2c | 27.97 ± 1.59d | 3.92 ± 0.79f | 8.41 ± 0.7g |
| MU2(NaCl) | 23.61 ± 1.56b | 32.11 ± 1.91c | 5.54 ± 0.31de | 10.68 ± 0.62ef |
| Si(NaCl) | 23.33 ± 2.08b | 32.39 ± 1.22c | 5.93 ± 0.23cd | 11.82 ± 0.82de |
| P(NaCl) | 23 ± 2b | 31.16 ± 1.10c | 5.02 ± 0.08e | 10.39 ± 0.47f |
| Si+MU2(NaCl) | 29 ± 1a | 36.03 ± 1.25b | 6.63 ± 0.25b | 12.0 ± 0.86d |
| P+MU2(NaCl) | 30.33 ± 2.51a | 37.35 ± 1.18ab | 6.5 ± 0.19bc | 13.31 ± 0.31c |
| Si+P+MU2(NaCl) | 30.66 ± 2.08a | 38.17 ± 1.35ab | 6.99 ± 0.2b | 14.63 ± 0.98b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, A.; Khan, M.A.; Lee, K.-E.; Kang, S.-M.; Dhungana, S.K.; Bhusal, N.; Lee, I.-J. The Halotolerant Rhizobacterium—Pseudomonas koreensis MU2 Enhances Inorganic Silicon and Phosphorus Use Efficiency and Augments Salt Stress Tolerance in Soybean (Glycine max L.). Microorganisms 2020, 8, 1256. https://doi.org/10.3390/microorganisms8091256
Adhikari A, Khan MA, Lee K-E, Kang S-M, Dhungana SK, Bhusal N, Lee I-J. The Halotolerant Rhizobacterium—Pseudomonas koreensis MU2 Enhances Inorganic Silicon and Phosphorus Use Efficiency and Augments Salt Stress Tolerance in Soybean (Glycine max L.). Microorganisms. 2020; 8(9):1256. https://doi.org/10.3390/microorganisms8091256
Chicago/Turabian StyleAdhikari, Arjun, Muhammad Aaqil Khan, Ko-Eun Lee, Sang-Mo Kang, Sanjeev Kumar Dhungana, Narayan Bhusal, and In-Jung Lee. 2020. "The Halotolerant Rhizobacterium—Pseudomonas koreensis MU2 Enhances Inorganic Silicon and Phosphorus Use Efficiency and Augments Salt Stress Tolerance in Soybean (Glycine max L.)" Microorganisms 8, no. 9: 1256. https://doi.org/10.3390/microorganisms8091256