Niche Differentiation of Active Methane-Oxidizing Bacteria in Estuarine Mangrove Forest Soils in Taiwan

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
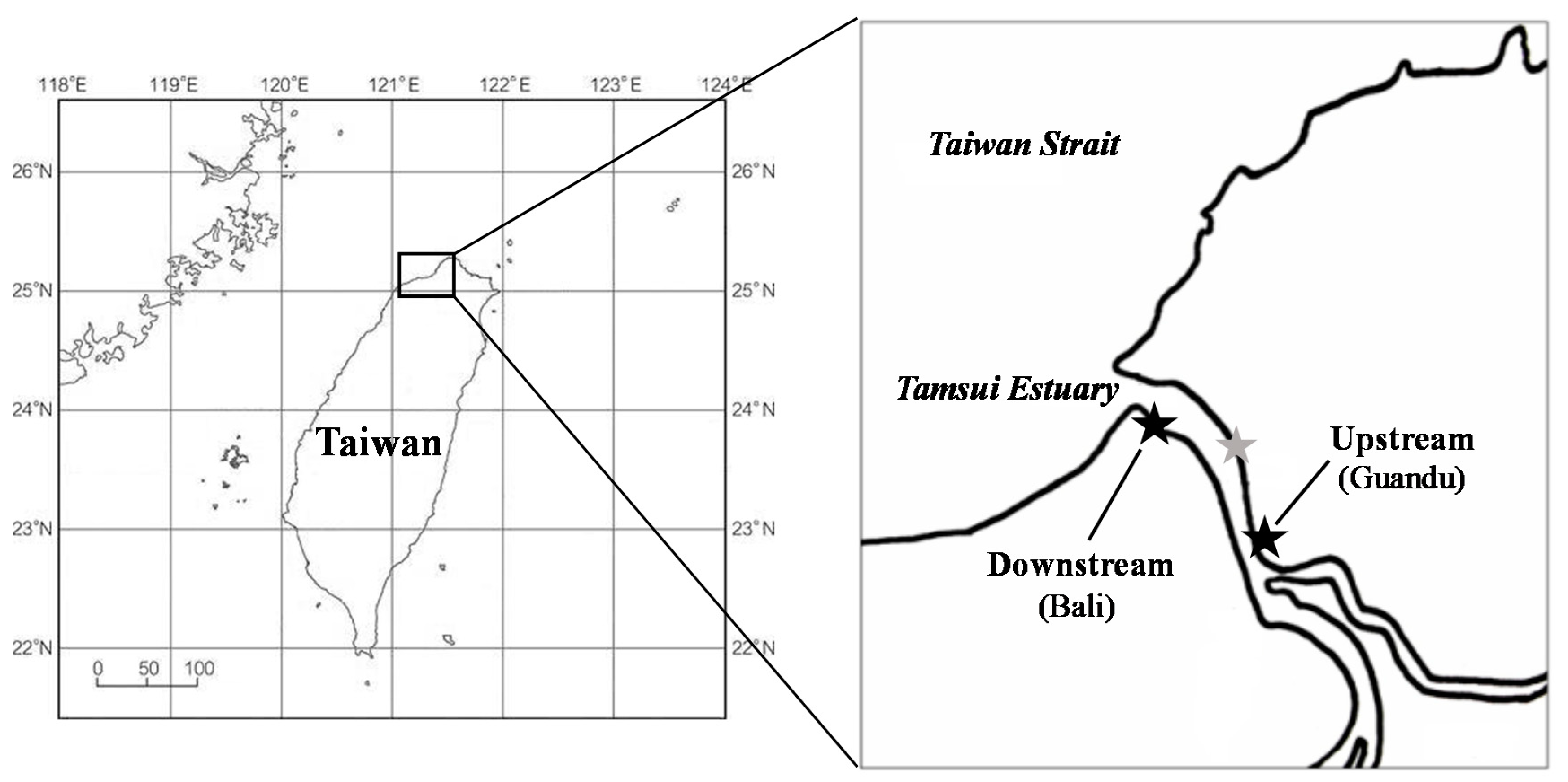
2.1. Site Description and Soil Sampling
2.2. Aerobic CH4 Oxidation Experiment and DNA-SIP Gradient Fractionation
2.3. Analyses of Active Methanotrophic Communities
2.4. Statistical Analyses
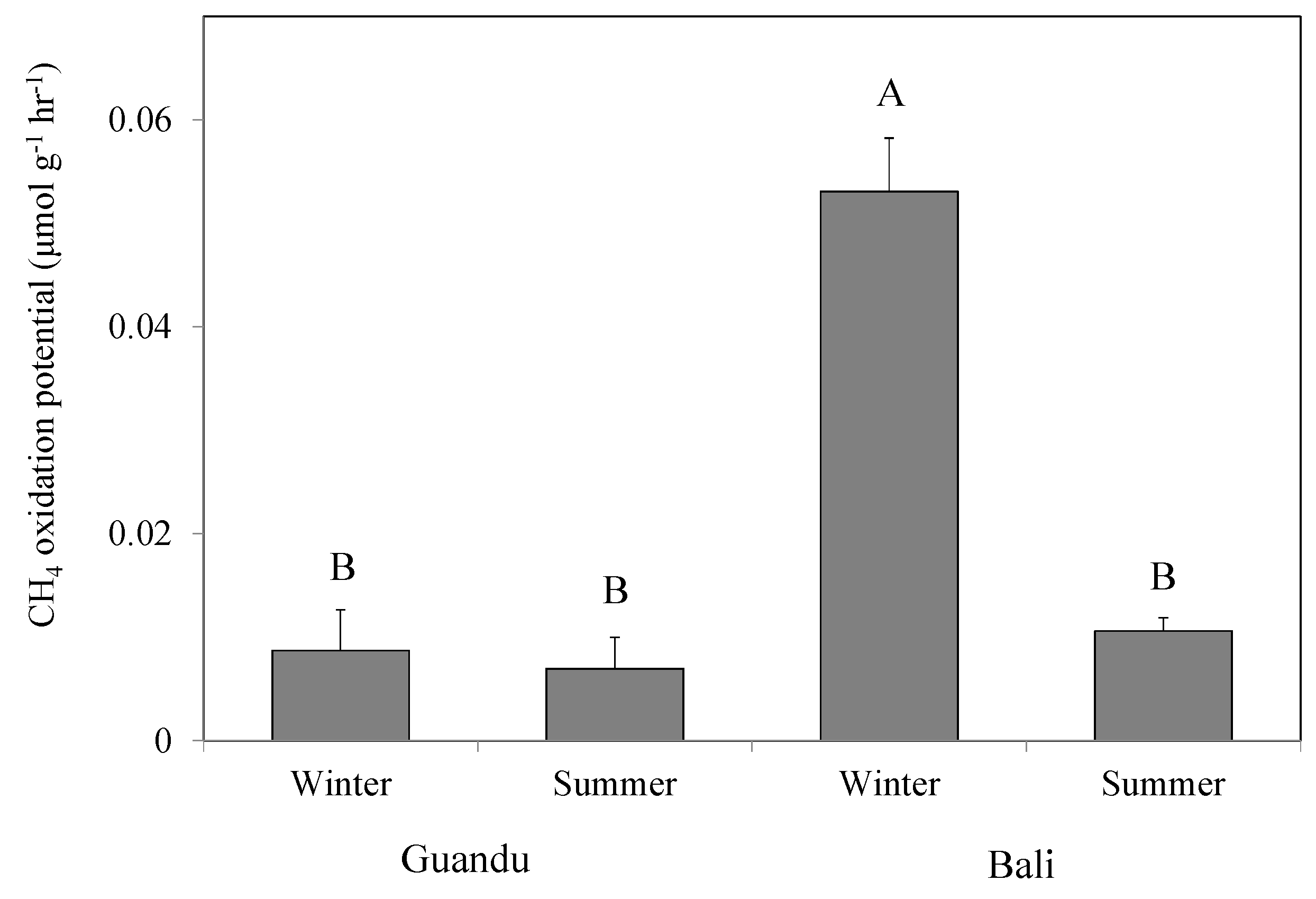
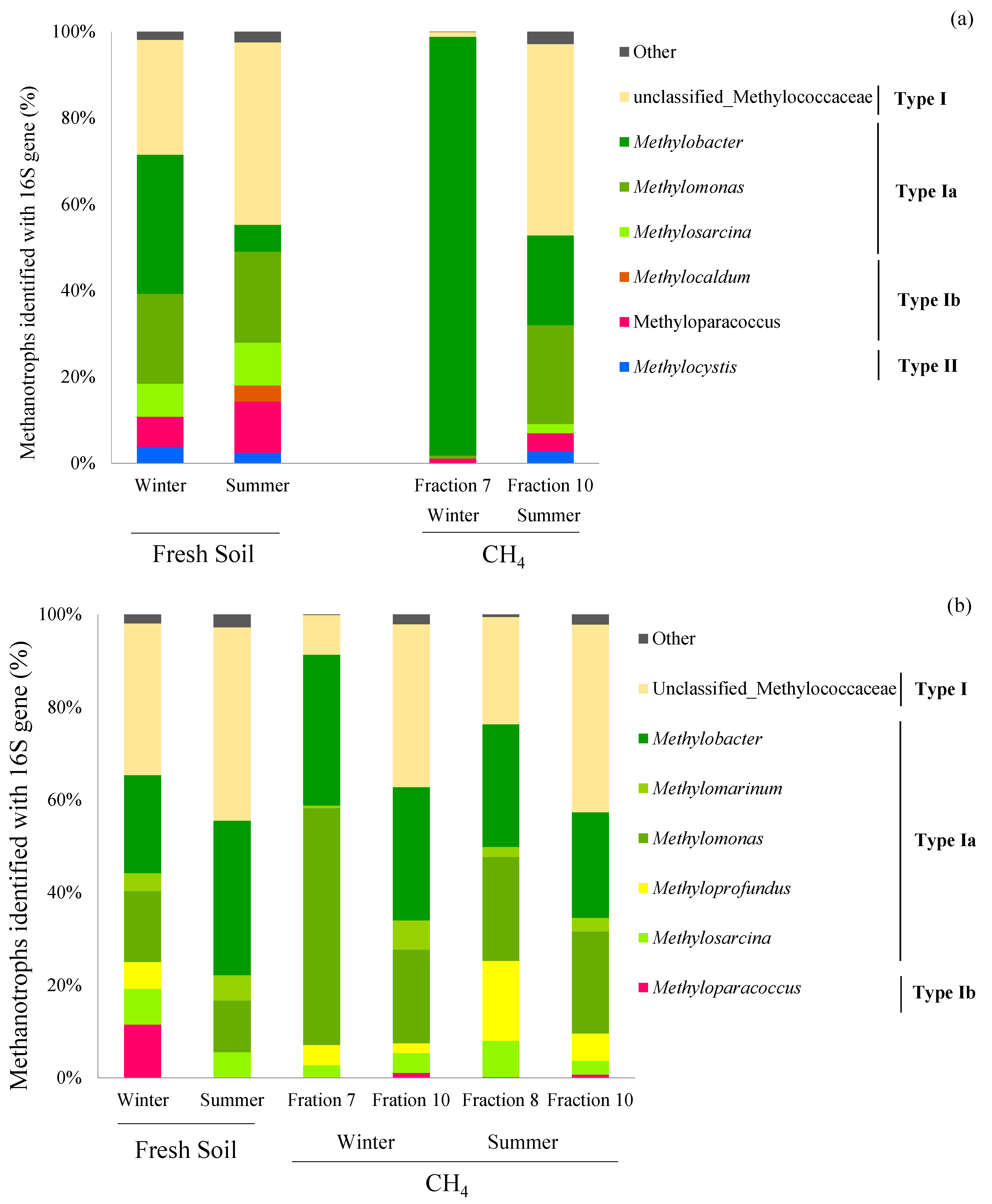
3. Results
4. Discussion
4.1. Composition and Adaptation of Methanotrophs in Field Mangrove Soils
4.2. Active Methanotrophs Identified with 13CH4 DNA-SIP
4.3. pmoA and 16S rRNA Sequences
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Weart, S.R. The Discovery of Global Warming; Harvard University Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Yang, S.-S.; Chang, H.-L. Effect of environmental conditions on methane production and emission from paddy soil. Agric. Ecosyst. Environ. 1998, 69, 69–80. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- He, R.; Wooller, M.J.; Pohlman, J.W.; Catranis, C.; Quensen, J.; Tiedje, J.M.; Leigh, M.B. Identification of functionally active aerobic methanotrophs in sediments from an arctic lake using stable isotope probing. Environ. Microbiol. 2012, 14, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Angel, R.; Veraart, A.J.; Daebeler, A.; Jia, Z.; Kim, S.Y.; Kerckhof, F.-M.; Boon, N.; Bodelier, P.L.E. Biotic interactions in microbial communities as modulators of biogeochemical processes: Methanotrophy as a model system. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, C.E.; Martínez-Lorenzo, A.; Brady, A.L.; Grasby, S.E.; Dunfield, P.F. Methanotrophic bacteria in warm geothermal spring sediments identified using stable-isotope probing. FEMS Microbiol. Ecol. 2014, 90, 92–102. [Google Scholar] [CrossRef]
- Zheng, Y.; Huang, R.; Wang, B.Z.; Bodelier, P.L.E.; Jia, Z.J. Competitive interactions between methane- and ammonia-oxidizing bacteria modulate carbon and nitrogen cycling in paddy soil. Biogeosciences 2014, 11, 3353–3368. [Google Scholar] [CrossRef]
- Ho, A.; Kerckhof, F.-M.; Luke, C.; Reim, A.; Krause, S.; Boon, N.; Bodelier, P.L.E. Conceptualizing functional traits and ecological characteristics of methane-oxidizing bacteria as life strategies. Environ. Microbiol. Rep. 2013, 5, 335–345. [Google Scholar] [CrossRef]
- Shrestha, M.; Shrestha, P.M.; Frenzel, P.; Conrad, R. Effect of nitrogen fertilization on methane oxidation, abundance, community structure, and gene expression of methanotrophs in the rice rhizosphere. ISME J. 2010, 4, 1545–1556. [Google Scholar] [CrossRef] [Green Version]
- Bowman, J.P.; Sly, L.I.; Nichols, P.D.; Hayward, A.C. Revised taxonomy of the methanotrophs—Description of Methylobacter gen. nov., emendation of Methylococcus, validation of Methylosinus and Methylocystis species, and a proposal that the family Methylococcaceae includes only the group I Methanotrophs. Int. J. Syst. Bacteriol. 1993, 43, 735–753. [Google Scholar] [CrossRef]
- Gagliano, A.L.; D’Alessandro, W.; Tagliavia, M.; Parello, F.; Quatrini, P. Methanotrophic activity and diversity of methanotrophs in volcanic geothermal soils at Pantelleria (Italy). Biogeosciences 2014, 11, 5865–5875. [Google Scholar] [CrossRef] [Green Version]
- Wise, M.G.; McArthur, J.V.; Shimkets, L.J. Methanotroph diversity in landfill soil: Isolation of novel Type I and Type II methanotrophs whose presence was suggested by culture-independent 16S Ribosomal DNA analysis. Appl. Environ. Microbiol. 1999, 65, 4887–4897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakemian, A.S.; Rosenzweig, A.C. The biochemistry of methane oxidation. Annu. Rev. Biochem. 2007, 76, 223–241. [Google Scholar] [CrossRef] [PubMed]
- McDonald, I.R.; Bodrossy, L.; Chen, Y.; Murrell, J.C. Molecular ecology techniques for the study of aerobic methanotrophs. Appl. Environ. Microbiol. 2008, 74, 1305–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Zheng, Y.; Bodelier, P.L.E.; Conrad, R.; Jia, Z. Conventional methanotrophs are responsible for atmospheric methane oxidation in paddy soils. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Heyer, J.; Berger, U.; Hardt, M.; Dunfield, P.F. Methylohalobius crimeensis gen. nov., sp. nov., a moderately halophilic, methanotrophic bacterium isolated from hypersaline lakes of Crimea. Int. J. Sys. Evol. Microbiol. 2005, 55, 1817–1826. [Google Scholar] [CrossRef] [Green Version]
- Sherry, A.; Osborne, K.A.; Sidgwick, F.R.; Gray, N.D.; Talbot, H.M. A temperate river estuary is a sink for methanotrophs adapted to extremes of pH, temperature and salinity. Environ. Microbiol. Rep. 2016, 8, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Shiau, Y.J.; Cai, Y.F.; Lin, Y.T.; Jia, Z.; Chiu, C.Y. Community structure of active aerobic methanotrophs in red mangrove (Kandelia obovata) soils under different frequency of tides. Microb. Ecol. 2017, 75. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Cai, Y.F.; Jia, Z.J.; Chen, C.L.; Chiu, C.Y. Phylogenetically distinct methanotrophs modulate methane oxidation in rice paddies across Taiwan. Soil Biol. Biochem. 2018, 124, 59–69. [Google Scholar] [CrossRef]
- Zheng, Y.; Jia, Z. The application of biomarker genes for DNA/RNA-Stable Isotope Probing of active methanotrophs responsible for aerobic methane oxidation in six paddy soils. Acta Pedol. Sin. 2016, 53, 490–501. [Google Scholar] [CrossRef]
- Bowman, J.P. Methylobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Ho, A.; Mo, Y.L.; Lee, H.J.; Sauheitl, L.; Jia, Z.J.; Horn, M.A. Effect of salt stress on aerobic methane oxidation and associated methanotrophs; a microcosm study of a natural community from a non-saline environment. Soil Biol. Biochem. 2018, 125, 210–214. [Google Scholar] [CrossRef]
- Radajewski, S.; Ineson, P.; Parekh, N.R.; Murrell, J.C. Stable-isotope probing as a tool in microbial ecology. Nature 2000, 403, 646. [Google Scholar] [CrossRef] [PubMed]
- Shiau, Y.J.; Dham, V.; Tian, G.; Chiu, C.Y. Factors influencing removal of sewage nitrogen through denitrification in mangrove soils. Wetlands 2016, 36, 621–630. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chou, C.H. The distribution and influence of heavy-metals in mangrove forests of the tamshui estuary in Taiwan. Soil Sci. Plant Nutr. 1991, 37, 659–669. [Google Scholar] [CrossRef]
- Olmstead, L.B.; Alexander, L.T.; Middleton, H.E. A Pipette Method of Mechanical Analysis of Soils Based on Improved Dispersion Procedure; United States Department of Agriculture, Economic Research Service: Washington, DC, USA, 1930. [Google Scholar]
- Shiau, Y.J.; Chiu, C.Y. Changes in soil biochemical properties in a cedar plantation invaded by moso bamboo. Forests 2017, 8, 222. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Wang, H.C.; Chen, T.H.; Jien, S.H.; Tian, G.L.; Chiu, C.Y. Improvement in the biochemical and chemical properties of badland soils by thorny bamboo. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, J.W. Determination of Nitrate-Nitrite Nitrogen by Automated Colorimetry 600/R-93-100; U.S. Environmental Protection Agency: Cincinnati, OH, USA, 1993.
- Huang, C.-Y.; Jien, S.-H.; Chen, T.-H.; Tian, G.; Chiu, C.-Y. Soluble organic C and N and their relationships with soil organic C and N and microbial characteristics in moso bamboo (Phyllostachys edulis) plantations along an elevation gradient in Central Taiwan. J. Soils Sediments 2014, 14, 1061–1070. [Google Scholar] [CrossRef]
- Sollins, P.; Glassman, C.; Paul, E.A.; Swanston, C.; Lajtha, K.; Heil, J.W.; Elliott, E.T. Soil Carbon and Nitrogen: Pools and Fractions. In Standard Soil Methods for Long-Term Ecological Research; Robertson, G.P., Coleman, D.C., Bledsoe, C.S., Sollins, P., Eds.; Oxford University Press: New York, NY, USA, 1999; pp. 89–105. [Google Scholar]
- Waring, S.A.; Bremner, J.M. Effect of soil mesh-size on estimation of mineralizable nitrogen in soils. Nature 1964, 202, 1141. [Google Scholar] [CrossRef]
- Kolb, S.; Knief, C.; Stubner, S.; Conrad, R. Quantitative Detection of Methanotrophs in Soil by Novel pmoA-Targeted Real-Time PCR Assays. Appl. Environ. Microbiol. 2003, 69, 2423. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Jia, Z.J. Urease gene-containing Archaea dominate autotrophic ammonia oxidation in two acid soils. Environ. Microbiol. 2013, 15, 1795–1809. [Google Scholar] [CrossRef]
- Fish, J.A.; Chai, B.; Wang, Q.; Sun, Y.; Brown, C.T.; Tiedje, J.M.; Cole, J.R. FunGene: The functional gene pipeline and repository. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, A.P.; Aurikko, J.P.; Kelly, D.P. A challenge for 21st century molecular biology and biochemistry: What are the causes of obligate autotrophy and methanotrophy? FEMS Microbiol. Rev. 2004, 28, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.F.; Link, H.; Liesack, W. Response of Methylocystis sp Strain SC2 to Salt Stress: Physiology, global transcriptome, and amino acid profiles. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Song, W.; Zhao, Y.; Zhou, J.; Wang, Z.; Luo, Y.; Li, Y.; Lin, G. Differential responses of litter decomposition to regional excessive nitrogen input and global warming between two mangrove species. Estuar. Coast. Shelf Sci. 2018, 214, 141–148. [Google Scholar] [CrossRef]
- Shiau, Y.-J.; Chiu, C.-Y. Biogeochemical Processes of C and N in the Soil of Mangrove Forest Ecosystems. Forests 2020, 11, 492. [Google Scholar] [CrossRef]
- Chen, W.-B.; Liu, W.-C.; Hsu, M.-H. Modeling assessment of a saltwater intrusion and a transport time scale response to sea-level rise in a tidal estuary. Environ. Fluid Mech. 2015, 15, 491–514. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Burchell, M.R.; Krauss, K.W.; Birgand, F.; Broome, S.W. Greenhouse gas emissions from a created brackish marsh in eastern North Carolina. Wetlands 2016, 36, 1009–1024. [Google Scholar] [CrossRef]
- Trotsenko, Y.A.; Khmelenina, V.N. Aerobic methanotrophic bacteria of cold ecosystems. FEMS Microbiol. Ecol. 2005, 53, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.; Yu, Z.; Li, K.; Zhang, H. Diversity, abundance and vertical distribution of methane-oxidizing bacteria (methanotrophs) in the sediments of the Xianghai wetland, Songnen Plain, northeast China. J. Soils Sediments 2013, 13, 242–252. [Google Scholar] [CrossRef]
- Bowman, J.P.; Sly, L.I.; Stackebrandt, E. The phylogenetic position of the family Methylococcaceae. Int. J. Syst. Bacteriol. 1995, 45, 182–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, M.G.; Luke, C.; Deng, Y.; Frenzel, P. Classification of pmoA amplicon pyrosequences using BLAST and the lowest common ancestor method in MEGAN. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Mizrahi-Man, O.; Davenport, E.R.; Gilad, Y. Taxonomic classification of bacterial 16S rRNA genes using short sequencing reads: Evaluation of effective study designs. PLoS ONE 2013, 8, e53608. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Season | Surface Water Salinity (psu) | pH | Soil Redox (mV) | Soil Texture | TOC | TN | SbOC | NH4+ | NO3− | SbON | TDN | PMN |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (%) | (mg C kg−1 Soil) | (mg N kg−1 Soil) | |||||||||||
| Upstream (Guandu) | Winter | 20 | 3.6 ± 0.0 | −208 ± 73 | silty clay | 4.0 ± 0.8 | 0.3 ± 0.0 | 21.4 ± 1.8 | 10.0 ± 1.2 | 0.3 ± 0.1 | 21.0 ± 9.0 | 31.3 ± 7.9 | 14.3 ± 1.3 |
| Summer | 17 | 5.7 ± 0.0 | 3.4 ± 0.0 | 0.3 ± 0.0 | 12.7 ± 2.5 | 7.7 ± 0.2 | 0.3 ± 0.0 | 12.8 ± 4.8 | 20.7 ± 4.9 | 11.9 ± 1.0 | |||
| Downstream (Bali) | Winter | 20 | 5.6 ± 0.0 | −196 ± 16 | clay loam | 2.0 ± 0.1 | 0.3 ± 0.0 | 9.6 ± 2.0 | 6.5 ± 1.7 | 0.4 ± 0.1 | 18.3 ± 5.0 | 25.1 ± 6.2 | 32.6 ± 3.1 |
| Summer | 21 | 6.2 ± 0.0 | 2.9 ± 0.0 | 0.3 ± 0.0 | 6.4 ± 3.8 | 54.2 ± 0.7 | 0.3 ± 0.0 | 27.7 ± 15.3 | 82.2 ± 15.4 | 20.7 ± 3.7 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiau, Y.-J.; Lin, C.-W.; Cai, Y.; Jia, Z.; Lin, Y.-T.; Chiu, C.-Y. Niche Differentiation of Active Methane-Oxidizing Bacteria in Estuarine Mangrove Forest Soils in Taiwan. Microorganisms 2020, 8, 1248. https://doi.org/10.3390/microorganisms8081248
Shiau Y-J, Lin C-W, Cai Y, Jia Z, Lin Y-T, Chiu C-Y. Niche Differentiation of Active Methane-Oxidizing Bacteria in Estuarine Mangrove Forest Soils in Taiwan. Microorganisms. 2020; 8(8):1248. https://doi.org/10.3390/microorganisms8081248
Chicago/Turabian StyleShiau, Yo-Jin, Chiao-Wen Lin, Yuanfeng Cai, Zhongjun Jia, Yu-Te Lin, and Chih-Yu Chiu. 2020. "Niche Differentiation of Active Methane-Oxidizing Bacteria in Estuarine Mangrove Forest Soils in Taiwan" Microorganisms 8, no. 8: 1248. https://doi.org/10.3390/microorganisms8081248