Fungal Community in Antarctic Soil Along the Retreating Collins Glacier (Fildes Peninsula, King George Island)


Abstract
:1. Introduction
2. Methods
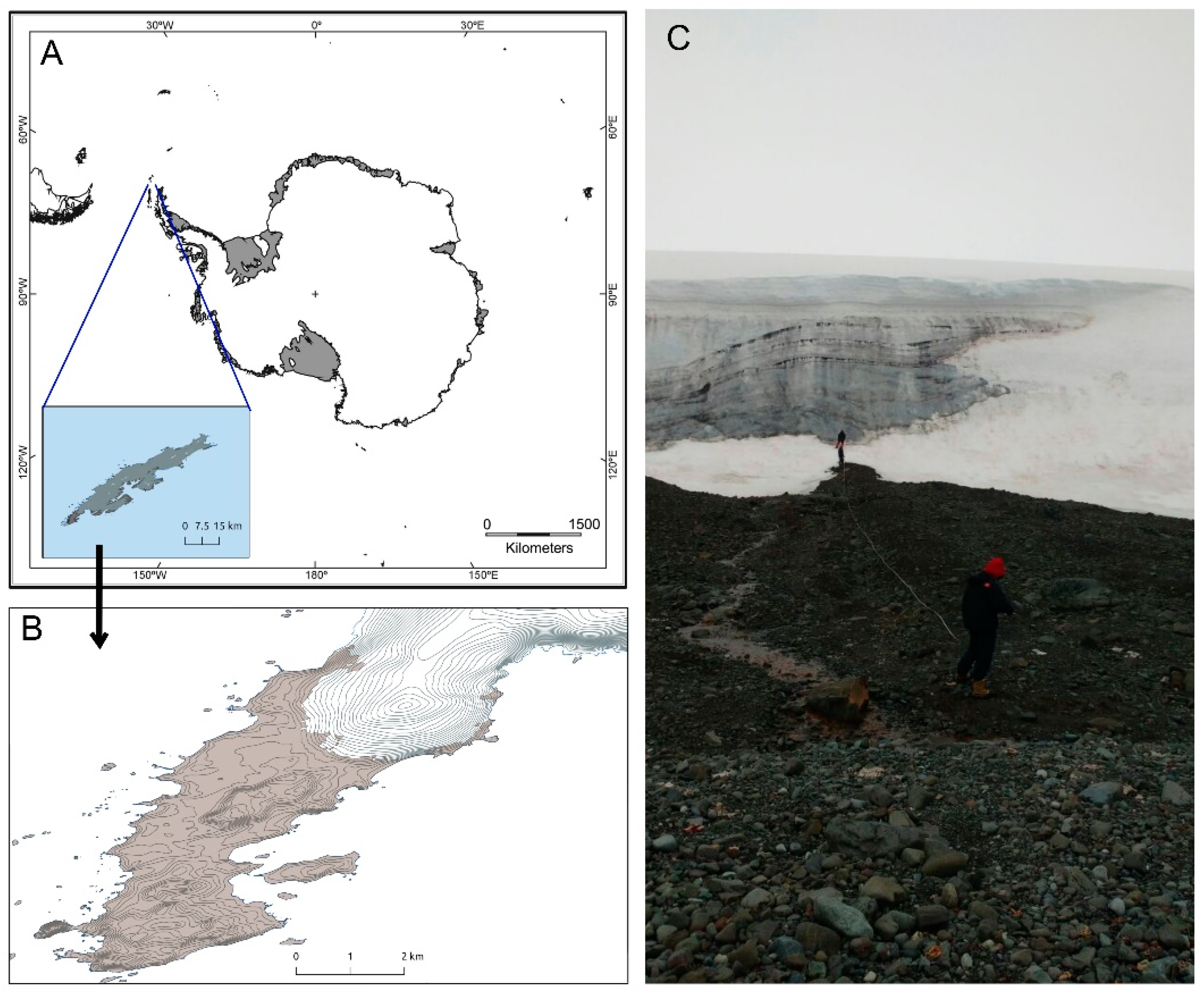
2.1. Sampling Site
2.2. Physicochemical Analysis
2.3. Fungal Isolation
2.4. Fungal Identification
2.5. Accession Numbers
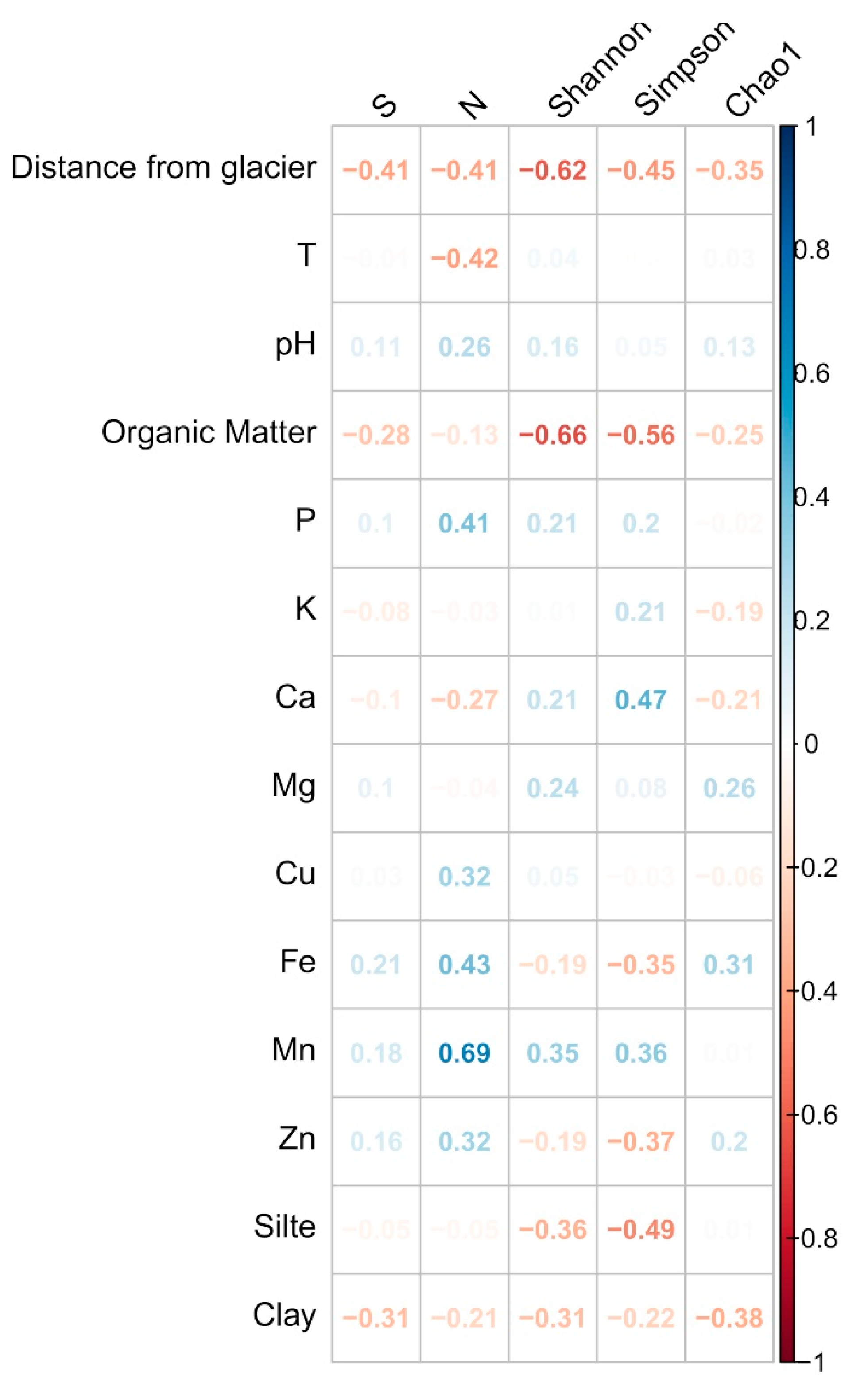
2.6. Structure and Composition of Fungal Communities
3. Results
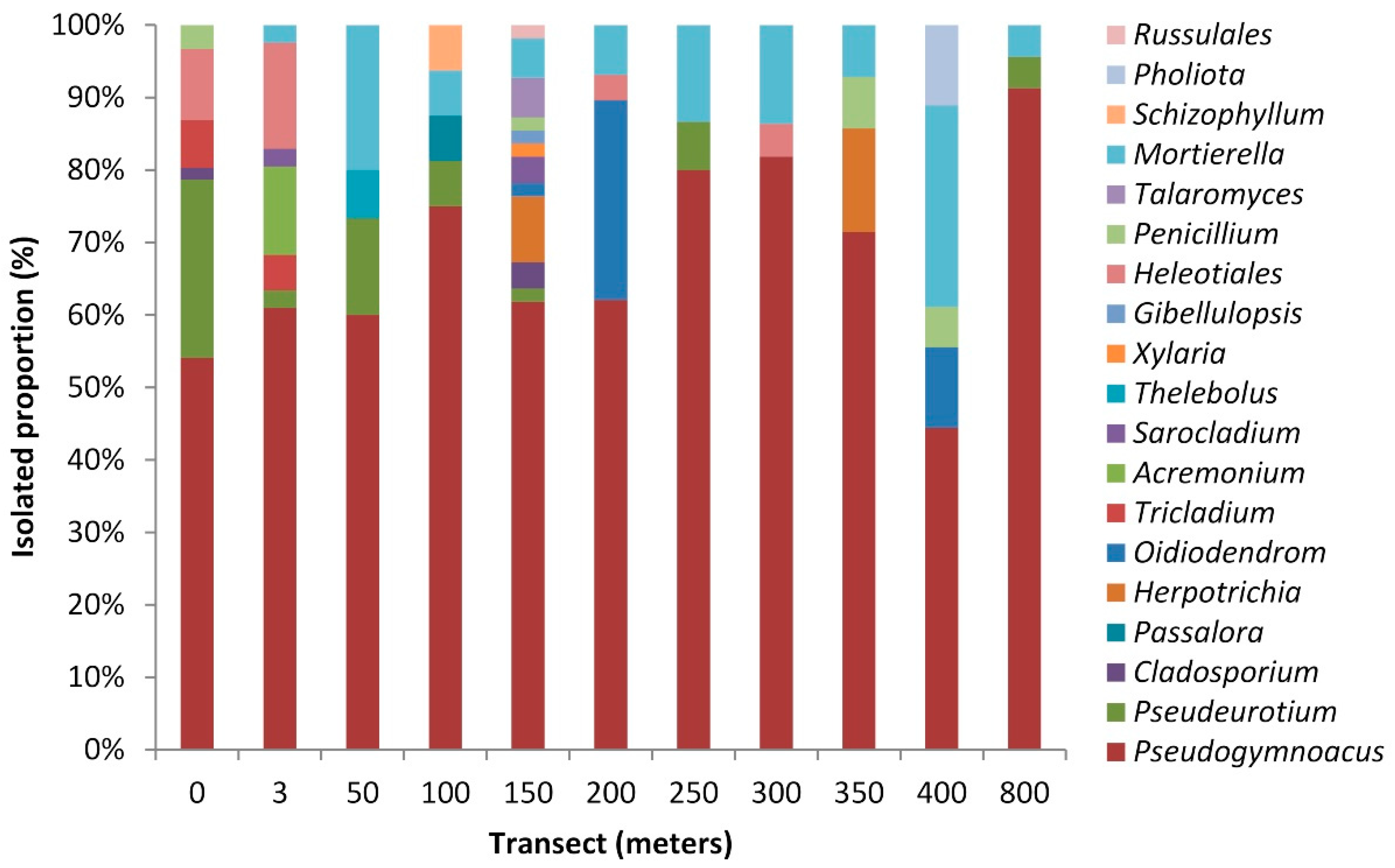
3.1. Fungi from Collins Glacier Soil Samples
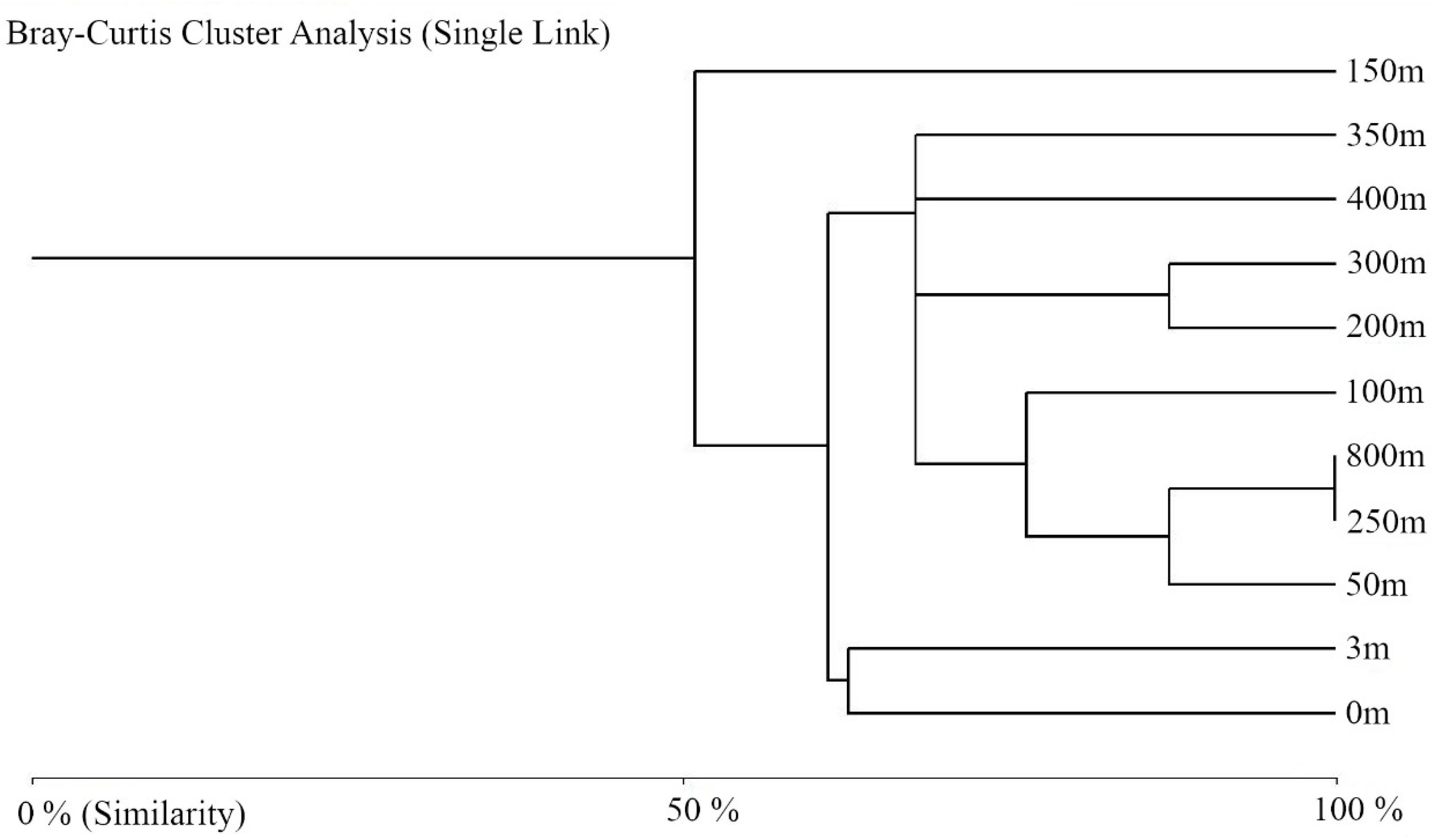
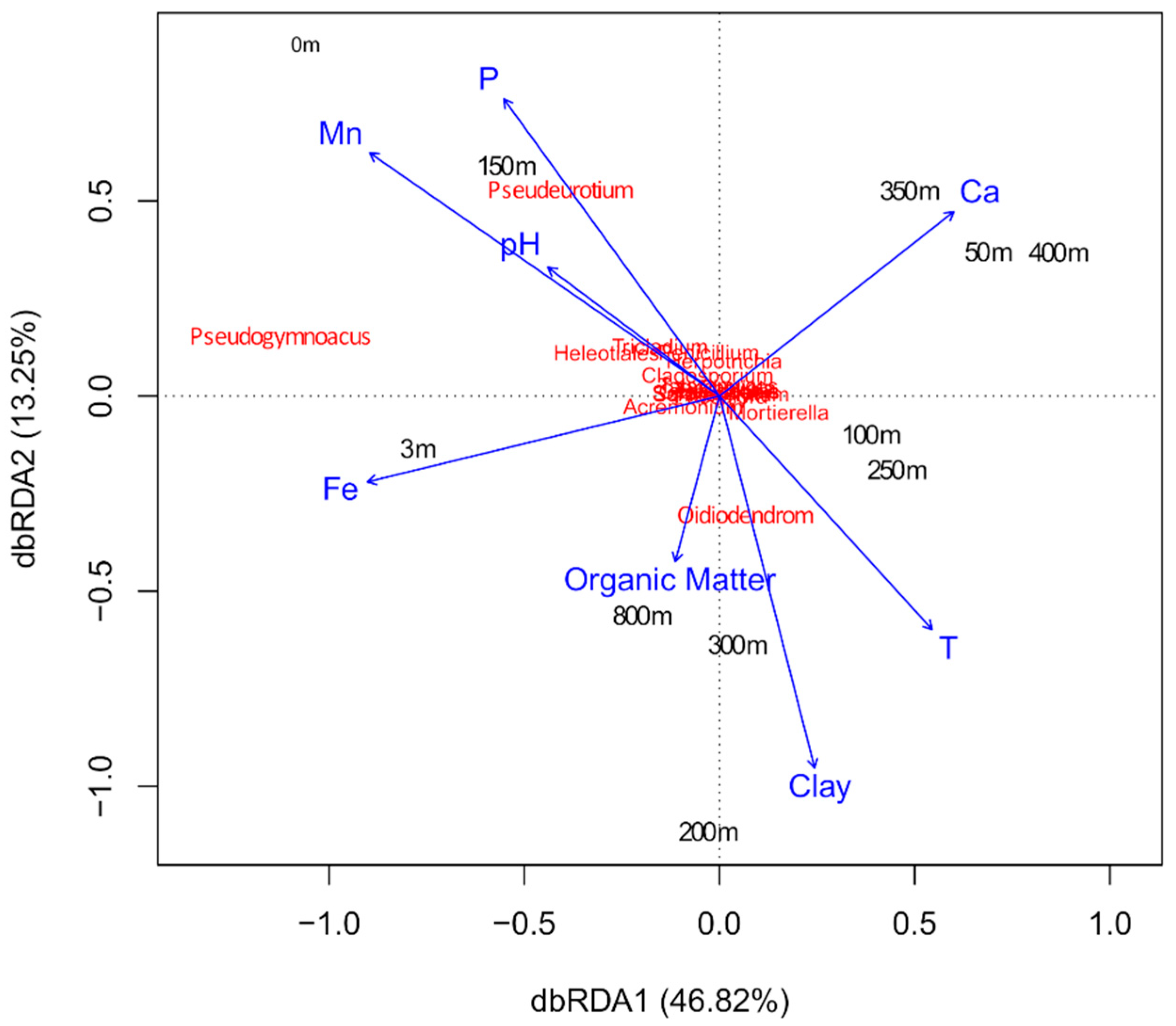
3.2. Structure and Composition of the Fungal Communities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McClintock, J.; Ducklow, H.; Fraser, W. Ecological responses to climate change on the Antarctic Peninsula. Am. Sci. 2008, 4, 302–310. [Google Scholar] [CrossRef]
- Turner, J.; Barrand, N.E.; Bracegirdle, T.J.; Convey, P.; Hodgson, D.; Jarvis, M.; Jenkins, A.; Marshall, G.; Meredith, M.P.; Roscoe, H.; et al. Antarctic climate change and the environment: An update. Polar Rec. 2014, 50, 237–259. [Google Scholar] [CrossRef] [Green Version]
- Bromwich, D.H.; Nicolas, J.P.; Monaghan, A.J.; Lazzara, M.A.; Keller, L.M.; Weidner, G.A.; Wilson, A.B. Central West Antarctica among the most rapidly warming regions on Earth. Nat. Geosci. 2013, 6, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, J.P.; Bromwich, D.H. New reconstruction of Antarctic near-surface temperatures: Multidecadal trends and reliability of global reanalyses. J. Clim. 2014, 21, 8070–8093. [Google Scholar] [CrossRef]
- Rückamp, M.; Braun, M.; Suckro, S.; Blindow, N. Observed glacial changes on the King George Island ice cap, Antarctica, in the last decade. Glob. Planet. Chang. 2011, 79, 99–109. [Google Scholar] [CrossRef]
- Meredith, M.P.; King, J.C. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys. Res. Lett. 2005, 32, 1–5. [Google Scholar] [CrossRef]
- Simões, C.L.; Rosa, K.K.D.; Czapela, F.F.; Vieira, R.; Simões, J.C. Collins Glacier retreat process and regional climatic variations, King George Island, Antarctica. Geogr. Rev. 2015, 105, 462–471. [Google Scholar] [CrossRef]
- Hall, B.L. Late-Holocene advance of the Collins Ice Cap, King George Island, South Shetland Islands. Holocene 2007, 17, 1253–1258. [Google Scholar] [CrossRef]
- Lee, J.R.; Raymond, B.; Bracegirdle, T.J.; Chades, I.; Fuller, R.A.; Shaw, J.D.; Terauds, A. Climate change drives expansion of Antarctic ice-free habitat. Nature 2017, 547, 49–54. [Google Scholar] [CrossRef]
- Duffy, G.A.; Coetzee, B.W.; Latombe, G.; Akerman, A.H.; McGeoch, M.A.; Chown, S.L. Barriers to globally invasive species are weakening across the Antarctic. Divers. Distrib. 2017, 23, 982–996. [Google Scholar] [CrossRef] [Green Version]
- Pertierra, L.R.; Aragón, P.; Shaw, J.D.; Bergstrom, D.M.; Terauds, A.; Olalla-Tárraga, M.Á. Global thermal niche models of two European grasses show high invasion risks in Antarctica. Glob. Chang. Biol. 2017, 23, 2863–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunde-Cimerman, N.; Sonjak, S.; Zalar, P.; Frisvad, J.C.; Diderichsen, B.; Plemenitaš, A. Extremophilic fungi in arctic ice: A relationship between adaptation to low temperature and water activity. Phys. Chem. Earth Parts A/B/C 2003, 28, 1273–1278. [Google Scholar] [CrossRef]
- Abyzov, S.S. Microorganisms in the Antarctic ice. Antarct. Microbiol. 1993, 1, 265–296. [Google Scholar]
- Brown, S.P.; Jumpponen, A. Contrasting primary successional trajectories of fungi and bacteria in retreating glacier soils. Mol. Ecol. 2014, 23, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Freedman, Z.; Zak, D.R. Soil bacterial communities are shaped by temporal and environmental filtering: Evidence from a long-term chronosequence. Environ. Microbiol. 2015, 17, 3208–3218. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.; Blaalid, R.; Vik, U.; Carlsen, T.; Kauserud, H.; Eidesen, P.B. Primary succession of Bistorta vivipara (L.) Delabre (Polygonaceae) root-associated fungi mirrors plant succession in two glacial chronosequences. Environ. Microbiol. 2015, 17, 2777–2790. [Google Scholar] [CrossRef]
- Blaalid, R.; Carlsen, T.; Kumar, S.; Halvorsen, R.; Ugland, K.I.; Fontana, G.; Kauserud, H. Changes in the root- associated fungal communities along a primary successional gradient analysed by 454 pyrosequencing. Mol. Ecol. 2012, 21, 1897–1908. [Google Scholar] [CrossRef]
- Mühlmann, O.; Peintner, U. Ectomycorrhiza of Kobresia myosuroides at a primary successional glacier forefront. Mycorrhiza 2008, 18, 355–362. [Google Scholar] [CrossRef]
- Newsham, K.K.; Hopkins, D.W.; Carvalhais, L.C.; Fretwell, P.T.; Rushton, S.P.; O’Donnel, A.G.; Dennis, P.G. Relationship between soil fungal diversity and temperature in the maritime Antarctic. Nat. Clim. Chang. 2016, 6, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Cavicchioli, R. Microbial ecology of Antarctic aquatic systems. Nat. Rev. Microbiol. 2015, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. The ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Dane, J.H.; Hopmans, J.W. Pressure plate extractor. In Methods of Soil Analysis. Physical Methods. Madison, Soil Science Society of America Part 4; (SSSA Book Series. 5) SSSA: Madison, WI, USA, 2002; pp. 643–670. [Google Scholar]
- Van Raij, B.; Andrade, J.C.; Cantarella, H.; Quaggio, J.A. Análise Química para Avaliação da Fertilidade de Solos Tropicais. Fundação Cargill 2001, 285. [Google Scholar]
- Lacerda, L.T.; Gusmão, L.F.P.; Rodrigues, A. Diversity of endophytic fungi in Eucalyptus microcorys assessed by complementary isolation methods. Mycol. Prog. 2018, 17, 1–9. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Burns, T.; Lee, S.; Taylor, J. PCR Protocols: A guide to method and applications. In Amplification and Direct Sequencing of Fungal Ribosomal rRNA Genes for Phylogenetics; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–352. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nuclei. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Edwards, A.; Irvine-Fynn, T.; Mitchell, A.C.; Rassner, S.M.E. A germ theory for glacial systems? Wiley Interdiscip. Rev. Water 2014, 1, 331–340. [Google Scholar] [CrossRef]
- Bradley, J.A.; Singarayer, J.S.; Anesio, A.M. Microbial Community dynamics in the forefield of glaciers. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruisi, S.; Barreca, D.; Selbmann, L.; Zucconi, L.; Onofri, S. Fungi in Antarctica. Rev. Environ. Sci. Biotechnol. 2007, 6, 127–141. [Google Scholar] [CrossRef]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Farrell, R.L.; Blanchette, R.A. Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol. Biochem. 2006, 38, 3057–3064. [Google Scholar] [CrossRef]
- Dreesens, L.; Lee, C.; CARY, S. The distribution and identity of edaphic fungi in the McMurdo Dry Valleys. Biology 2014, 3, 466–483. [Google Scholar] [CrossRef] [Green Version]
- Pudasaini, S.; Wilson, J.; Ji, M.; van Dorst, J.; Snape, I.; Palmer, A.S.; Burns, B.P.; Ferrari, B.C. Microbial diversity of browning peninsula, Eastern Antarctica revealed using molecular and cultivation methods. Front. Microbiol. 2017, 8, 591. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.W.F.; Barato, M.B.; Nobre, F.S.; Polezel, D.A.; de Oliveira, T.B.; dos Santos, J.A.; Rodrigues, A.; Sette, L.D. Production of cold-adapted enzymes by filamentous fungi from King George Island, Antarctica. Polar Biol. 2018, 41, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- Canini, F.; Geml, J.; D’Acqui, L.P.; Selbmann, L.; Onofri, S.; Ventura, S.; Zucconi, L. Exchangeable cations and pH drive diversity and functionality of fungal communities in biological soil crusts from coastal sites of Victoria Land, Antarctica. Fungal Ecol. 2020, 45, 1–11. [Google Scholar] [CrossRef]
- Lorch, J.M.; Lindner, D.L.; Gargas, A.; Muller, L.K.; Minnis, A.M.; Blehert, D.S. A culture based survey of fungi in soil from bat hibernacula in the eastern United States and its implications for detection of Geomyces destructans, the causal agent of bat white-nose syndrome. Mycologia 2013, 105, 237–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnis, A.M.; Lindner, D.L. Phylogenetic evaluation of Geomyces and allies reveals no close relatives of Pseudogymnoascus destructans, comb. nov., in bat hibernacula of eastern North America. Fungal Biol. 2013, 117, 638–649. [Google Scholar] [CrossRef]
- Kochkina, G.A.; Ivanushkina, N.E.; Akimov, V.N.; Gilichinskii, D.A.; Ozerskaya, S.M. Halo-and psychrotolerant Geomyces fungi from Arctic cryopegs and marine deposits. Microbiology 2007, 76, 31–38. [Google Scholar] [CrossRef]
- Wentzel, L.C.P.; Inforsato, F.J.; Montoya, Q.V.; Rossin, B.G.; Nascimento, N.R.; Rodrigues, A.; Sette, L.D. Fungi from admiralty bay (King George Island, Antarctica) soils and marine sediments. Microb. Ecol. 2019, 77, 12–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosi, S.; Casado, B.; Gerdol, R.; Caretta, G. Fungi isolated from antarctic mosses. Polar Biol. 2002, 25, 262–268. [Google Scholar] [CrossRef]
- Godinho, V.M.; Furbino, L.E.; Santiago, I.F.; Pellizzari, F.M.; Yokoya, N.S.; Pupo, D.; Alves, T.M.A.; Junior, P.A.S.; Romanha, A.J.; Zani, C.L.; et al. Diversity and bioprospecting of fungal communities associated with endemic and cold-adapted macroalgae in Antarctica. ISME J. 2013, 7, 1434–1451. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Xu, X.; Qi, S. Diverse deep-sea fungi from the South China Sea and their antimicrobial activity. Curr. Microbiol. 2013, 67, 525–530. [Google Scholar] [CrossRef]
- Furbino, L.E.; Godinho, V.M.; Santiago, I.F.; Pellizari, F.M.; Alves, T.M.A.; Zani, C.L.; Junior, P.A.S.; Romanha, A.J.; Carvalho, A.G.O.; Gil, L.H.V.G.; et al. Diversity patterns, ecology and biological activities of fungal communities associated with the endemic macroalgae across the Antarctic Peninsula. Microb. Ecol. 2014, 67, 775–787. [Google Scholar] [CrossRef]
- Gilichinsky, D.A.; Wilson, G.S.; Friedmann, E.I.; McKay, C.P.; Sletten, R.S.; Rivkina, E.M.; Vishnivetskaya, T.A.; Erokhina, L.G.; Ivanushkina, N.E.; Kochkina, G.A.; et al. Microbial populations in Antarctic permafrost: Biodiversity, state, age, and implication for astrobiology. Astrobiology 2007, 7, 275–311. [Google Scholar] [CrossRef]
- Edgington, S.; Thompson, E.; Moore, D.; Hughes, K.A.; Bridge, P. Investigating the insecticidal potential of Geomyces (Myxotrichaceae: Helotiales) and Mortierella (Mortierellacea: Mortierellales) isolated from Antarctica. SpringerPlus 2014, 3, 289. [Google Scholar] [CrossRef] [Green Version]
- Melo, I.S.; Santos, S.N.; Rosa, L.H.; Parma, M.M.; Silva, L.J.; Queiroz, S.C.; Pellizari, V.H. Isolation and biological activities of an endophytic Mortierella alpina strain from the Antarctic moss Schistidium antarctici. Extremophiles 2014, 18, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.N.; Carvalho, C.R.; Johann, S.; Mendes, G.; Alves, T.M.A.; Zani, C.L.; Junior, P.A.S.; Murta, S.M.F.; Romanha, A.J.; Cantrell, C.L.; et al. Antibacterial, antifungal and antiprotozoal activities of fungal communities present in different substrates from Antarctica. Polar Biol. 2015, 38, 1143–1152. [Google Scholar] [CrossRef]
- Newsham, K.K.; Garnett, M.H.; Robinson, C.H.; Cox, F. Discrete taxa of saprotrophic fungi respire different ages of carbon from Antarctic soils. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinho, V.M.; de Paula, M.T.R.; Silva, D.A.S.; Paresque, K.; Martins, A.P.; Colepicolo, P.; Rosa, C.A.; Rosa, L.H. Diversity and distribution of hidden cultivable fungi associated with marine animals of Antarctica. Fungal Biol. 2019, 123, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, M.; Kim, S.; Yadav, D.R.; Um, Y.H.; Kim, H.S.; Lee, H.B.; Lee, Y.S. A New Record of Pseudeurotium bakeri from Crop Field Soil in Korea. Korean J. Mycol. 2016, 44, 145–149. [Google Scholar]
- Muller, L.K.; Lorch, J.M.; Lindner, D.L.; O’Connor, M.; Gargas, A.; Blehert, D.S. Bat white-nose syndrome: A real-time TaqMan polymerase chain reaction test targeting the intergenic spacer region of Geomyces destructans. Mycologia 2013, 105, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Malloch, D.; Sigler, L.; Hambleton, S.; Vanderwolf, K.J.; Gibas, C.F.C.; McAlpine, D.F. Fungi associated with hibernating bats in New Brunswick caves: The genus Leuconeurospora. Botany 2016, 94, 1171–1181. [Google Scholar] [CrossRef] [Green Version]
- Arenz, B.E.; Blanchette, R.A. Investigations of fungal diversity in wooden structures and soils at historic sites on the Antarctic Peninsula. Can. J. Microbiol. 2009, 55, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Ogaki, M.B.; Teixeira, D.R.; Vieira, R.; Lírio, J.M.; Felizardo, J.P.; Abuchacra, R.C.; Cardoso, R.P.; Zanif, C.L.; Alves, T.M.A.; Junior, P.A.S.; et al. Diversity and bioprospecting of cultivable fungal assemblages in sediments of lakes in the Antarctic Peninsula. Fungal Biol. 2020, 124, 601–611. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Gottlich, E.; Platas, G.; Genilloud, O.; Leotta, G.; van Brummelen, J. Evolution, taxonomy and ecology of the genus Thelebolus in Antarctica. Stud. Mycol. 2005, 51, 33–76. [Google Scholar]
- Marfenina, O.E.; Nikitin, D.A.; Ivanova, A.E. The structure of fungal biomass and diversity of cultivated micromycetes in Antarctic soils (progress and Russkaya Stations). Eurasian Soil Sci. 2016, 49, 934–941. [Google Scholar] [CrossRef]
- Santiago, I.F.; Alves, T.M.A.; Rabello, A.; Junior, P.A.S.; Romanha, A.J.; Zani, C.L.; Rosa, C.A.; Rosa, L.H. Leishmanicidal and antitumoral activities of endophytic fungi associated with the Antarctic angiosperms Deschampsia antarctica Desv. and Colobanthus quitensis (Kunth) Bartl. Extremophiles 2012, 16, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Kochkina, G.A.; Ivanushkina, N.E.; Lupachev, A.V.; Starodumova, I.P.; Vasilenko, O.V.; Ozerskaya, S.M. Diversity of mycelial fungi in natural and human-affected Antarctic soils. Polar Biol. 2019, 42, 47–64. [Google Scholar] [CrossRef]
- Ding, Z.; Li, L.; Che, Q.; Li, D.; Gu, Q.; Zhu, T. Richness and bioactivity of culturable soil fungi from the Fildes Peninsula, Antarctica. Extremophiles 2016, 20, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.N.; Vaz, A.B.M.; Rosa, C.A.; Rosa, L.H. Diversity and distribution of fungal communities in lakes of Antarctica. FEMS Microbiol. Ecol. 2012, 82, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alborés, S.; Sanguiñedo, P.; Held, B.H.; Cerdeiras, M.P.; Blanchette, R.A. Biodiversity and antimicrobial activity of Antarctic fungi from the Fildes Peninsula, King George Island. Sydowia 2018, 70, 185. [Google Scholar]
- Singer, R. Fungi distribution. In Terrestrial Life of Antarctica; Bushnell, V.C., Ed.; American Geographical Society: New York, NY, USA, 1967; Volume 5. [Google Scholar]
- Krishnan, A.; Convey, P.; Gonzalez-Rocha, G.; Alias, S.A. Production of extracellular hydrolase enzymes by fungi from King George Island. Polar Biol. 2016, 39, 65–76. [Google Scholar] [CrossRef]
- Kirtsideli, I.Y.; Abakumov, E.V.; Teshebaev, S.B.; Zelenskaya, M.S.; Vlasov, D.Y.; Krylenkov, V.A.; Ryabusheva, Y.V.; Sokolov, V.T.; Barantsevich, E.P. Microbial communities in regions of Arctic settlements. Gig. I Sanit. 2016, 95, 923–929. [Google Scholar] [CrossRef]
- Panin, A.L.; Sboychakov, V.B.; Belov, A.B.; Kraeva, L.A.; Vlasov, D.Y.; Goncharov, A.E. Natural and technogenic focality of infectious diseases in the territory of Antarctic settlements. Biol. Bull. Rev. 2016, 6, 320–332. [Google Scholar] [CrossRef]
- Dresch, P.; Falbesoner, J.; Ennemoser, C.; Hittorf, M.; Kuhnert, R.; Peintner, U. Emerging from the ice-fungal communities are diverse and dynamic in earliest soil developmental stages of a receding glacier. Environ. Microbiol. 2019, 21, 1864–1880. [Google Scholar] [CrossRef] [Green Version]
- Cox, F.; Newsham, K.K.; Robinson, C.H. Endemic and cosmopolitan fungal taxa exhibit differential abundances in total and active communities of Antarctic soils. Environ. Microbiol. 2019, 21, 1586–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, A.; Gené, J.; Sutton, D.A.; Madrid, H.; De Hoog, G.S.; Cano, J.; Guarro, J. Phylogeny of Sarocladium (Hypocreales). Pers. Mol. Phylogeny Evol. Fungi 2015, 34, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trowbridge, J.; Jumpponen, A. Fungal colonization of shrub willow roots at the forefront of a receding glacier. Mycorrhiza 2004, 14, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, S.D.; Palmer, A.S.; Winsley, T.; Lamb, E.; Bissett, A.; Brown, M.V.; van Dorst, J.; Ji, M.; Ferrari, B.C.; Grogan, P.; et al. Soil fertility is associated with fungal and bacterial richness, whereas pH is associated with community composition in polar soil microbial communities. Soil Biol. Biochem. 2014, 78, 10–20. [Google Scholar] [CrossRef]
- Green, T.G.A.; Broady, P.A. Biological soil crusts of Antarctica. In Biological Soil Crusts: Structure, Function, and Management; Springer: Berlin/Heidelberg, Germany, 2001; pp. 133–139. [Google Scholar]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (Meters) | S (N) * | Shannon | Inv-Simpson | Chao1 |
|---|---|---|---|---|
| 0 | 6 (61) | 1.26 | 2.71 | 6 |
| 3 | 7 (41) | 1.26 | 2.43 | 8.46 |
| 50 | 4 (15) | 1.08 | 2.37 | 4 |
| 100 | 5 (16) | 0.91 | 1.73 | 10.63 |
| 150 | 12 (55) | 1.51 | 2.49 | 16.91 |
| 200 | 4 (29) | 0.95 | 2.14 | 4 |
| 250 | 3 (15) | 0.63 | 1.51 | 3 |
| 300 | 3 (22) | 0.58 | 1.45 | 3 |
| 350 | 4 (14) | 0.90 | 1.85 | 4.46 |
| 400 | 5 (18) | 1.37 | 3.31 | 5 |
| 800 | 3 (23) | 0.36 | 1.19 | 3.96 |
| Variable | p-Value |
|---|---|
| Sample (meters) | 0.020 |
| T | 0.089 |
| Silt | 0.416 |
| Clay | 0.049 |
| O.M. | 0.277 |
| pH | 0.129 |
| P | 0.030 |
| K | 0.604 |
| Ca | 0.188 |
| Mg | 0.455 |
| Fe | 0.218 |
| Cu | 0.257 |
| Mn | 0.059 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, J.A.d.; Meyer, E.; Sette, L.D. Fungal Community in Antarctic Soil Along the Retreating Collins Glacier (Fildes Peninsula, King George Island). Microorganisms 2020, 8, 1145. https://doi.org/10.3390/microorganisms8081145
Santos JAd, Meyer E, Sette LD. Fungal Community in Antarctic Soil Along the Retreating Collins Glacier (Fildes Peninsula, King George Island). Microorganisms. 2020; 8(8):1145. https://doi.org/10.3390/microorganisms8081145
Chicago/Turabian StyleSantos, Juliana Aparecida dos, Edenilson Meyer, and Lara Durães Sette. 2020. "Fungal Community in Antarctic Soil Along the Retreating Collins Glacier (Fildes Peninsula, King George Island)" Microorganisms 8, no. 8: 1145. https://doi.org/10.3390/microorganisms8081145