Functional Identification and Evolutionary Analysis of Two Novel Plasmids Mediating Quinolone Resistance in Proteus vulgaris

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Condition
2.2. Plasmid Isolation and Sequencing
2.3. Construction of p3M-2A* and p3M-2B*
2.4. Plasmid Transformation
2.5. Antimicrobial Susceptibility Testing
2.6. Spot Growth Assays
2.7. Phylogenetic Analysis and Sequence Alignment
2.8. Plasmid Curing
2.9. Quantitative Real-Time PCR
2.10. Accession Number
3. Results
3.1. Effect of p3M-2A on the Expression of qnrD in p3M-2B
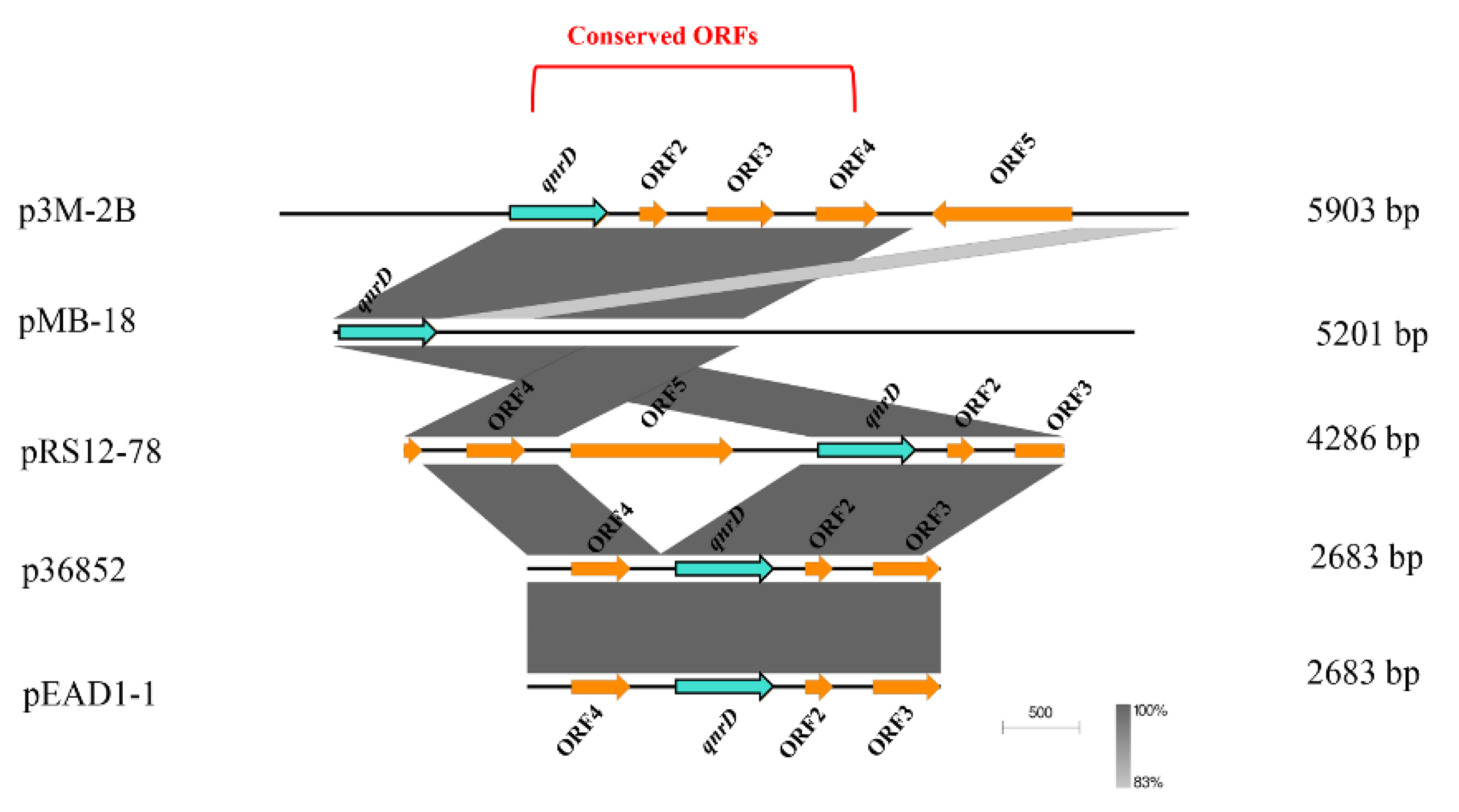
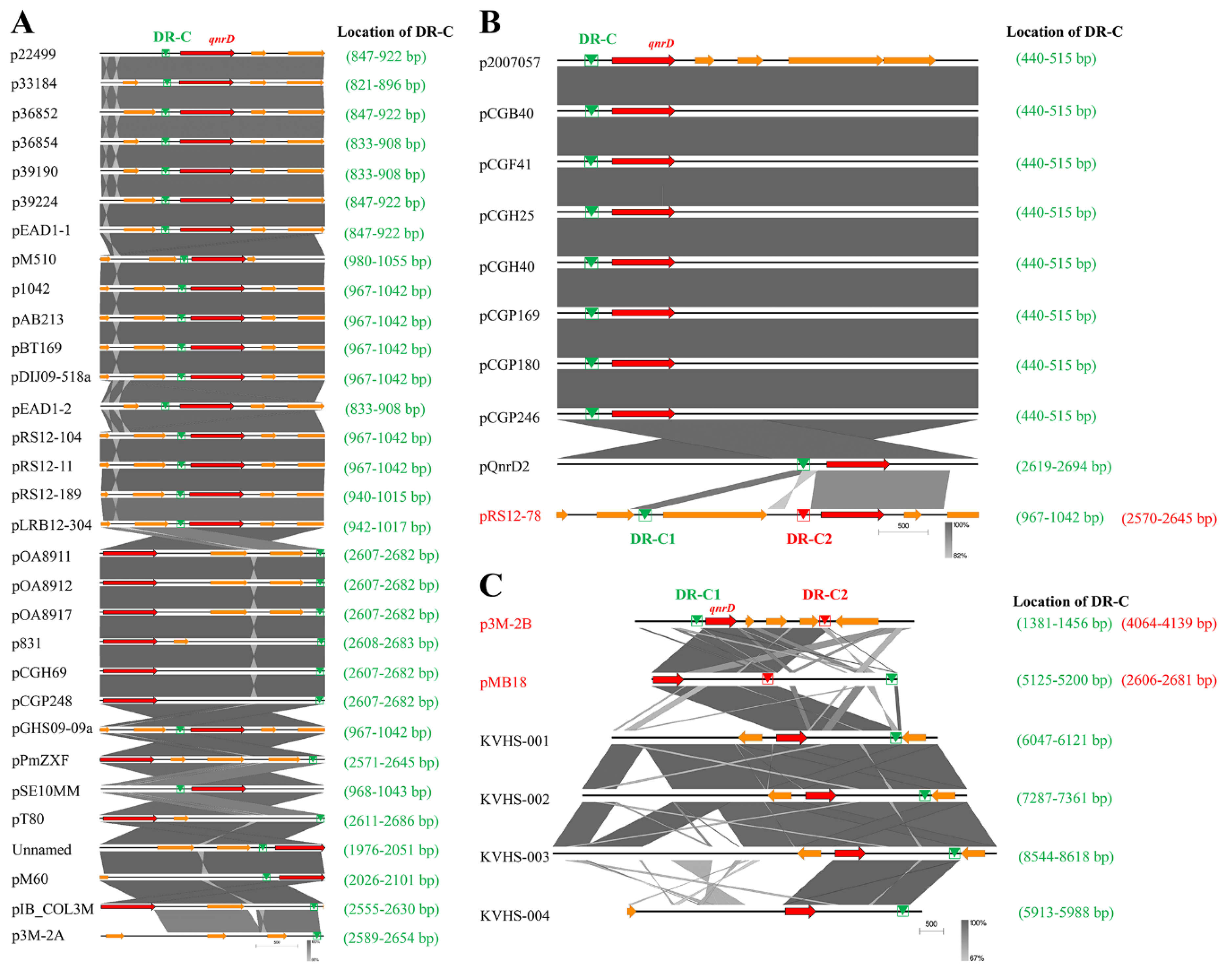
3.2. Phylogenetic Analysis of qnrD-Carrying Plasmids
3.3. Possible Formation Process of p3M-2B
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kumburu, H.H.; Sonda, T.; van Zwetselaar, M.; Leekitcharoenphon, P.; Lukjancenko, O.; Mmbaga, B.T.; Alifrangis, M.; Lund, O.; Aarestrup, F.M.; Kibiki, G.S. Using WGS to identify antibiotic resistance genes and predict antimicrobial resistance phenotypes in MDR Acinetobacter baumannii in Tanzania. J. Antimicrob. Chemother. 2019, 74, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Mcewen, S.A.; Collignon, P. Antimicrobial resistance: A one health perspective. Microbiol. Spectr. 2018, 6, 521–547. [Google Scholar]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. 2017, 57, 2857–2876. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Tate, H.P.; Zhao, S.; Li, C.; Dessai, U.; Simmons, M.; McDermott, P.F. Identification of plasmid-mediated quinolone resistance in Salmonella isolated from swine ceca and retail pork chops in the United States. Antimicrob. Agents Chemother. 2017, 61, e01318-17. [Google Scholar] [CrossRef] [Green Version]
- Adachi, F.; Yamamoto, A.; Takakura, K.I.; Kawahara, R. Occurrence of fluoroquinolones and fluoroquinolone-resistance genes in the aquatic environment. Sci. Total. Environ. 2013, 444, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.C. Emerging mechanisms of fluoroquinolone resistance. Emerg. Infect. Dis. 2001, 7, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Lautenbach, E.; Strom, B.L.; Nachamkin, I.; Bilker, W.B.; Marr, A.M.; Larosa, L.A.; Fishman, N.O. Longitudinal trends in fluoroquinolone resistance among Enterobacteriaceae isolates from inpatients and outpatients, 1989–2000: Differences in the emergence and epidemiology of resistance across organisms. Clin. Infect. Dis. 2004, 38, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.; Pons, M.J.; Gomes, C. Transferable mechanisms of quinolone resistance. Int. J. Antimicrob. Agents. 2012, 40, 196–203. [Google Scholar] [CrossRef]
- Pathirana, H.; Shin, G.W.; Wimalasena, S.; Silva, B.D.; Hossain, S.; Heo, G.J. Prevalence and characterization of quinolone resistance genes in Proteus species isolated from pet turtles. J. Exot. Pet Med. 2018, 27, 67–73. [Google Scholar] [CrossRef]
- Weigel, L.M.; Anderson, G.J.; Tenover, F.C. DNA gyrase and topoisomerase IV mutations associated with fluoroquinolone resistance in Proteus mirabilis. Antimicrob. Agents Chemother. 2002, 46, 2582–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klemm, E.J.; Shakoor, S.; Page, A.J.; Qamar, F.N.; Judge, K.; Saeed, K.D.; Wong, V.K.; Dallman, T.J.; Nair, S.; Baker, S.; et al. Emergence of an extensively drug-resistant Salmonella enterica serovar typhi clone harboring a promiscuous plasmid encoding resistance to fluoroquinolones and third-generation cephalosporins. mBio 2018, 9, e00105-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colobatiu, L.; Tabaran, A.; Flonta, M.; Oniga, O.; Mirel, S.; Mihaiu, M. First description of plasmid-mediated quinolone resistance determinants and β-lactamase encoding genes in non-typhoidal Salmonella isolated from humans, one companion animal and food in Romania. Gut. Pathog. 2015, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.W.; Thawng, C.N.; Lee, S.H.; Cha, C.J. Unique features of Aeromonas plasmid pAC3 and expression of the plasmid-mediated quinolone resistance genes. msphere 2017, 2, e00203-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strahilevitz, J.; Jacoby, G.A.; Hooper, D.C.; Robicsek, A. Plasmid-mediated quinolone resistance: A multifaceted threat. Clin. Microbiol. Rev. 2009, 22, 664–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Martínez, J.M.; Machuca, J.; Cano, M.E.; Calvo, J.; Martínez-Martínez, L.; Pascual, A. Plasmid-mediated quinolone resistance: Two decades on. Drug Resist. Updat. 2016, 29, 13–29. [Google Scholar] [CrossRef]
- Cavaco, L.M.; Hasman, H.; Xia, S.; Aarestrup, F.M. qnrD, a novel gene conferring transferable quinolone resistance in Salmonella enterica serovar Kentucky and Bovismorbificans strains of human origin. Antimicrob. Agents Chemother. 2009, 53, 603–608. [Google Scholar] [CrossRef] [Green Version]
- Guillard, T.; Cambau, E.; Neuwirth, C.; Nenninger, T.; Mbadi, A.; Brasme, L.; Vernet-Garnier, V.; Bajolet, O.; De Champs, C. Description of a 2683-base-pair plasmid containing qnrD in two Providencia rettgeri isolates. Antimicrob. Agents Chemother. 2012, 56, 565–568. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, Y.; Du, J.; Zhang, X.; Li, M.; Chen, H.; Yu, X.; Sun, Y.; Zhou, T. Description and plasmid characterization of the qnrD determinant in Proteeae in Wenzhou, Southern China. J. Microbiol. Immunol. Infect. 2018, 51, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Tran, J.H.; Jacoby, G.A.; Hooper, D.C. Interaction of the plasmid-encoded quinolone resistance protein QnrA with Escherichia coli topoisomerase IV. Antimicrob. Agents Chemother. 2005, 49, 3050–3052. [Google Scholar] [CrossRef] [Green Version]
- Guillard, T.; Grillon, A.; De Champs, C.; Cartier, C.; Madoux, J.; Berçot, B.; Lebreil, A.L.; Lozniewski, A.; Riahi, J.; Vernet-Garnier, V.; et al. Mobile insertion cassette elements found in small non-transmissible plasmids in Proteeae may explain qnrD mobilization. PLoS ONE 2014, 9, e87801. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, B.C.L.; Remondini, D.; Pasquini, G.; Janes, V.A.; Matamoros, S.; Schultsz, C. Quantifying the contribution of four resistance mechanisms to ciprofloxacin MIC in Escherichia coli: A systematic review. J. Antimicrob. Chemother. 2019, 74, 298–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraychete, G.B.; Campana, E.H.; Picão, R.; Bonelli, R.R. qnrD-harboring plasmids in Providencia spp. recovered from food and environmental Brazilian sources. Sci. Total Environ. 2019, 646, 1290–1292. [Google Scholar] [CrossRef] [PubMed]
- Prado, F.; Aguilera, A. Role of reciprocal exchange, one-ended invasion crossover and single-strand annealing on inverted and direct repeat recombination in Yeast: Different requirements for the RAD1, RAD10, and RAD52 genes. Genetics 1995, 139, 109–123. [Google Scholar] [PubMed]
- Oliveira, P.H.; Lemos, F.; Monteiro, G.A.; Prazeres, D.M.F. Recombination frequency in plasmid DNA containing direct repeats--predictive correlation with repeat and intervening sequence length. Plasmid 2005, 60, 159–165. [Google Scholar] [CrossRef]
- Dunn, M.J.; Anderson, M.Z. To repeat or not to repeat: Repetitive sequences regulate genome stability in Candida albicans. Genes 2019, 10, 866. [Google Scholar] [CrossRef] [Green Version]
- Matfield, M.; Badawi, R.; Brammar, W.J. Rec-dependent and Rec-independent recombination of plasmid-borne duplications in Escherichia Coli K12. Mol. Gen. Genet. 1985, 199, 518–523. [Google Scholar] [CrossRef]
- Bi, X.; Liu, L.F. recA-independent and recA-dependent intramolecular plasmid recombination. Differential homology requirement and distance effect. J. Mol. Biol. 1994, 235, 414–423. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Oliveira, P.H.; Prazeres, D.M.F.; Monteiro, G.A. High frequency plasmid recombination mediated by 28 bp direct repeats. Mol. Biotechnol. 2008, 40, 252–260. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.B.; Puhler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Teng, F.; Murray, B.E.; Weinstock, G.M. Conjugal transfer of plasmid DNA from Escherichia coli to Enterococci: A method to make insertion mutations. Plasmid 1998, 39, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. CLSI Approved Standard M100-S15; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Zhang, H.; Zhan, Y.; Yan, Y.; Liu, Y.; Hu, G.; Wang, S.; Yang, H.; Qiu, X.; Liu, Y.; Li, J.; et al. The Pseudomonas stutzeri-specific regulatory noncoding RNA NfiS targets katB mRNA encoding a catalase essential for optimal oxidative resistance and nitrogenase activity. J. Bacteriol. 2019, 201, e00334-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Verma, J.; Bag, S.; Saha, B.; Kumar, P.; Ghosh, T.S.; Dayal, M.; Senapati, T.; Mehra, S.; Dey, P.; Desigamani, A.; et al. Genomic plasticity associated with antimicrobial resistance in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2019, 116, 6226–6231. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Cai, J.; Zhang, R.; Zhou, H.; Sun, Q.; Chen, G. Emergence of Proteus mirabilis harboring blaKPC-2 and qnrD in a Chinese hospital. Antimicrob. Agents Chemother. 2012, 56, 2278–2282. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method, a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Goios, A.; Meirinhos, J.; Rocha, R.; Lopes, R.; Amorim, A.; Pereira, L. RepeatAround, A software tool for finding and visualizing repeats in circular genomes and its application to a human mtDNA database. Mitochondrion 2006, 6, 218–224. [Google Scholar] [CrossRef]
- Bi, X.; Liu, L.F. A replicational model for DNA recombination between direct repeats. J. Mol. Biol. 1996, 256, 849–858. [Google Scholar] [CrossRef]
- Al-Qahtani, M.; Safan, A.; Jassim, G.; Abadla, S. Efficacy of anti-microbial catheters in preventing catheter associated urinary tract infections in hospitalized patients: A review on recent updates. J. Infect. Public Health 2019, 12, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Mokracka, J.; Gruszczyńska, B.; Kaznowski, A. Integrons, β-lactamase and qnr genes in multidrug resistant clinical isolates of Proteus mirabilis and P. vulgaris. Apmis 2012, 120, 950–958. [Google Scholar] [CrossRef]
- Ro’żalski, A.; Sidorczyk, Z.; Kotełko, K. Potential virulence factors of Proteus bacilli. Microbiol. Mol. Biol. Rev. 1997, 61, 65–89. [Google Scholar] [CrossRef]
- Fu, Y.; Zepeda-Gurrola, R.C.; Aguilar-Gutiérrez, G.R.; Lara-Ramírez, E.E.; De Luna-Santillana, E.J.; Rodríguez-Luna, I.C.; Sánchez-Varela, A.; Carreño-López, R.; Moreno-Medina, V.R.; Rodríguez-Pérez, M.A.; et al. The detection of inherent homologous recombination between repeat sequences in H. pylori 26695 by the PCR-Based Method. Curr. Microbiol. 2014, 68, 211–219. [Google Scholar]
- He, L.; Partridge, S.R.; Yang, X.; Hou, J.; Deng, Y.; Yao, Q.; Zeng, Z.; Chen, Z.; Liu, J. Complete nucleotide sequence of pHN7A8, an F33:A-:B-type epidemic plasmid carrying blaCTX-M-65, fosA3 and rmtB from China. J. Antimicrob. Chemother. 2013, 68, 46–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Z.; Hu, L.; Jiang, X.; Zeng, L.; Feng, J.; Wu, W.; Chen, W.; Yang, H.; Yang, W.; Gao, B.; et al. Plasmid and chromosomal integration of four novel blaIMP-carrying transposons from Pseudomonas aeruginosa, Klebsiella pneumoniae and an Enterobacter sp. J. Antimicrob. Chemother. 2018, 73, 3005–3015. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids a | Relevant Characteristics | Source or Reference |
|---|---|---|
| Strains | ||
| P3M | Wild-type strain containing p3M-2A and p3M-2B plasmids | This study |
| P3M-Δ2A | p3M-2A-deficient strain | This study |
| P3M-Δ2B | p3M-2B-deficient strain | This study |
| P3M-Δ2AΔ2B | p3M-2A- and p3M-2B-deficient strain | This study |
| P3M-Δ2A/2A* | p3M-2A-deficient strain complemented with p3M-2A* | This study |
| P3M-Δ2B/2B* | p3M-2B-deficient strain complemented with p3M-2B* | This study |
| Escherichia coli DH5α | Competent cell for cloning b | CWBIO Company (CW0808S) |
| DH5α-2A | DH5α complemented with p3M-2A | This study |
| DH5α-2B | DH5α complemented with p3M-2B | This study |
| DH5α-2A2B | DH5α complemented with p3M-2A and p3M-2B | This study |
| DH5α-2B/2A* | DH5α-2B complemented with p3M-2A* | This study |
| DH5α-2A/2B* | DH5α-2A complemented with p3M-2B* | This study |
| E. coli S17-1 | Mobilizing donor strain with streptomycin resistance | [30] |
| Plasmids | ||
| p3M-2A | 2656-bp plasmid isolated from P3M | This study |
| p3M-2B | 5903-bp plasmid carrying qnrD, isolated from P3M | This study |
| p3M-2A* | p3M-2A without ORF1 | This study |
| p3M-2B* | p3M-2B without ORF5 | This study |
| Strains | MIC to Ciprofloxacin (mg/L) a |
|---|---|
| P3M | 1 |
| P3M-Δ2A | 0.5 |
| P3M-Δ2B | 0.125 |
| P3M-Δ2AΔ2B | 0.125 |
| E. coli DH5α-2A2B | 0.125 |
| E. coli DH5α-2B | 0.06 |
| E. coli DH5α-2A | 0.03 |
| E. coli DH5α | 0.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Chang, M.; Zhang, X.; Cai, P.; Dai, Y.; Song, T.; Wu, Z.; Xu, H.; Qiao, M. Functional Identification and Evolutionary Analysis of Two Novel Plasmids Mediating Quinolone Resistance in Proteus vulgaris. Microorganisms 2020, 8, 1074. https://doi.org/10.3390/microorganisms8071074
Zhang H, Chang M, Zhang X, Cai P, Dai Y, Song T, Wu Z, Xu H, Qiao M. Functional Identification and Evolutionary Analysis of Two Novel Plasmids Mediating Quinolone Resistance in Proteus vulgaris. Microorganisms. 2020; 8(7):1074. https://doi.org/10.3390/microorganisms8071074
Chicago/Turabian StyleZhang, Hongyang, Mingding Chang, Xiaochen Zhang, Peiyan Cai, Yixin Dai, Tongzhen Song, Zhenzhou Wu, Haijin Xu, and Mingqiang Qiao. 2020. "Functional Identification and Evolutionary Analysis of Two Novel Plasmids Mediating Quinolone Resistance in Proteus vulgaris" Microorganisms 8, no. 7: 1074. https://doi.org/10.3390/microorganisms8071074