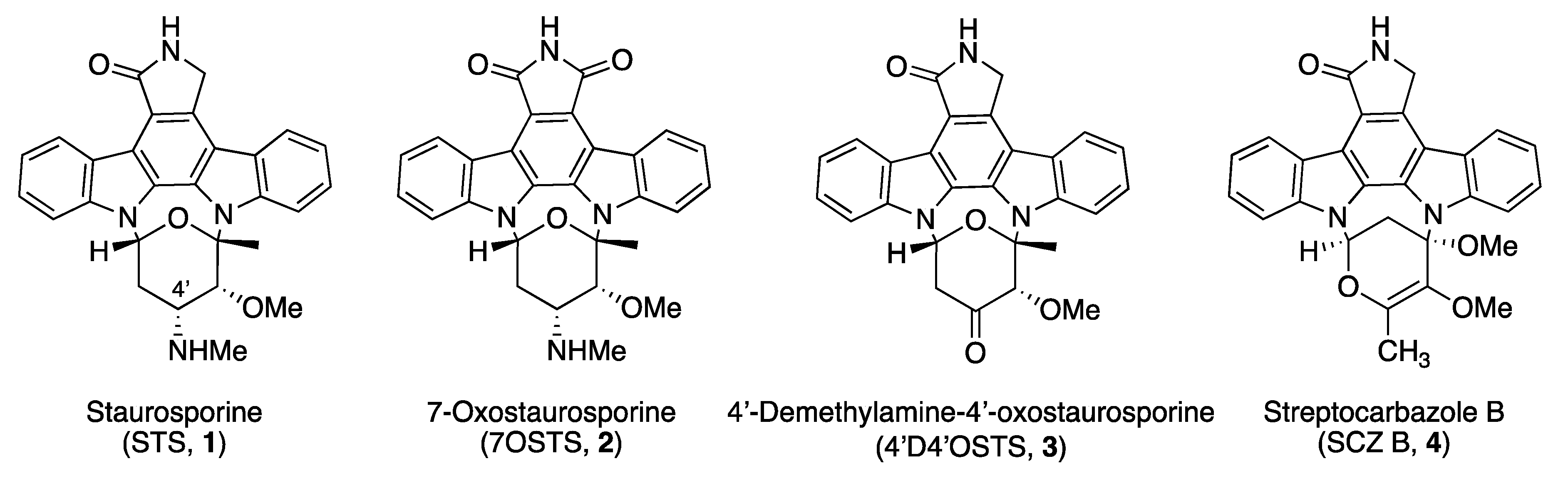
Evaluation of Indolocarbazoles from Streptomyces sanyensis as a Novel Source of Therapeutic Agents against the Brain-Eating Amoeba Naegleria fowleri

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Natural Compounds
2.2. Chemicals
2.3. Amoebic Strains and Cell Line Maintenance
2.4. Amoebicidal Activity and Cytotoxicity Assays
2.4.1. Activity Assays
2.4.2. Cytotoxicity Assays
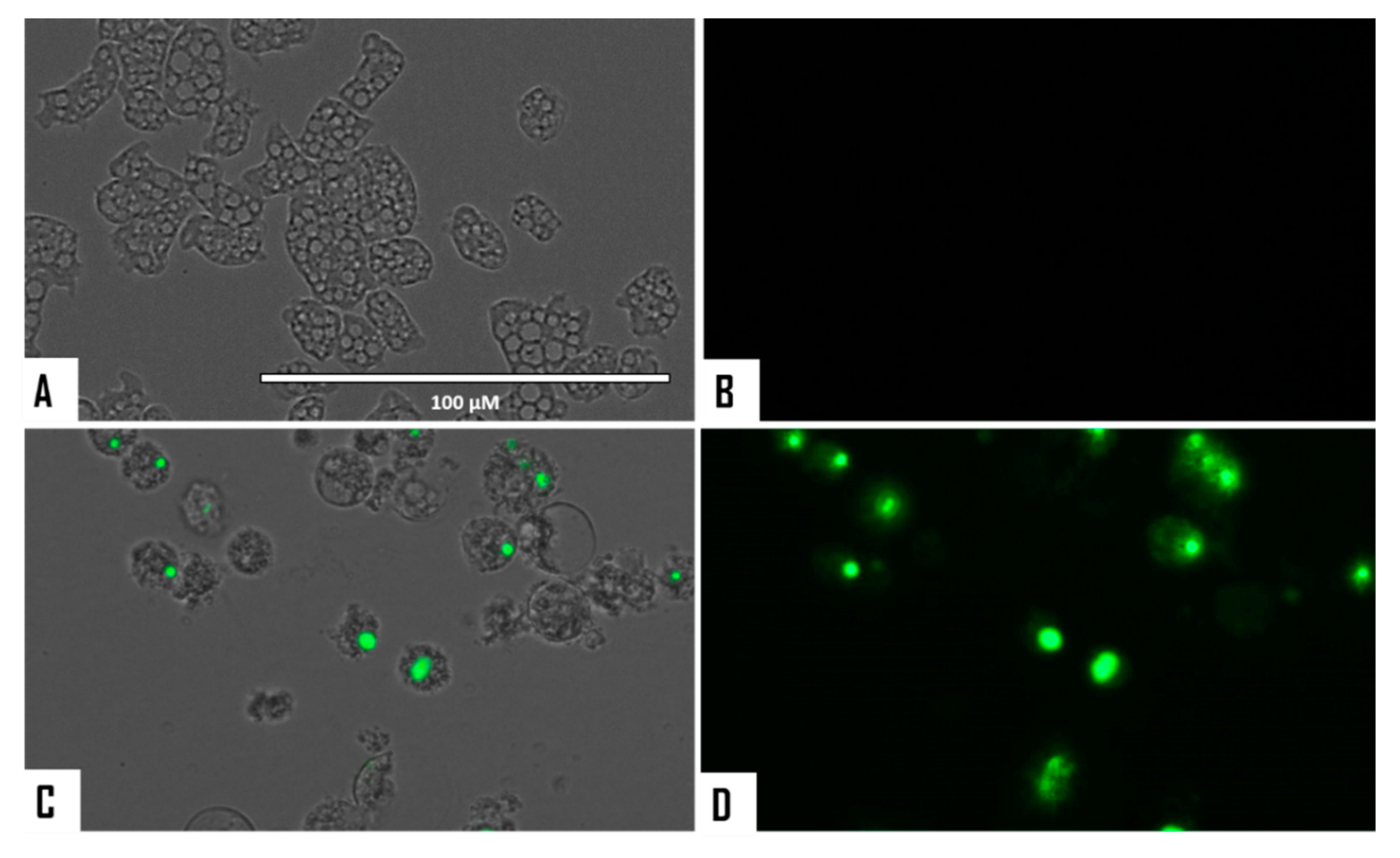
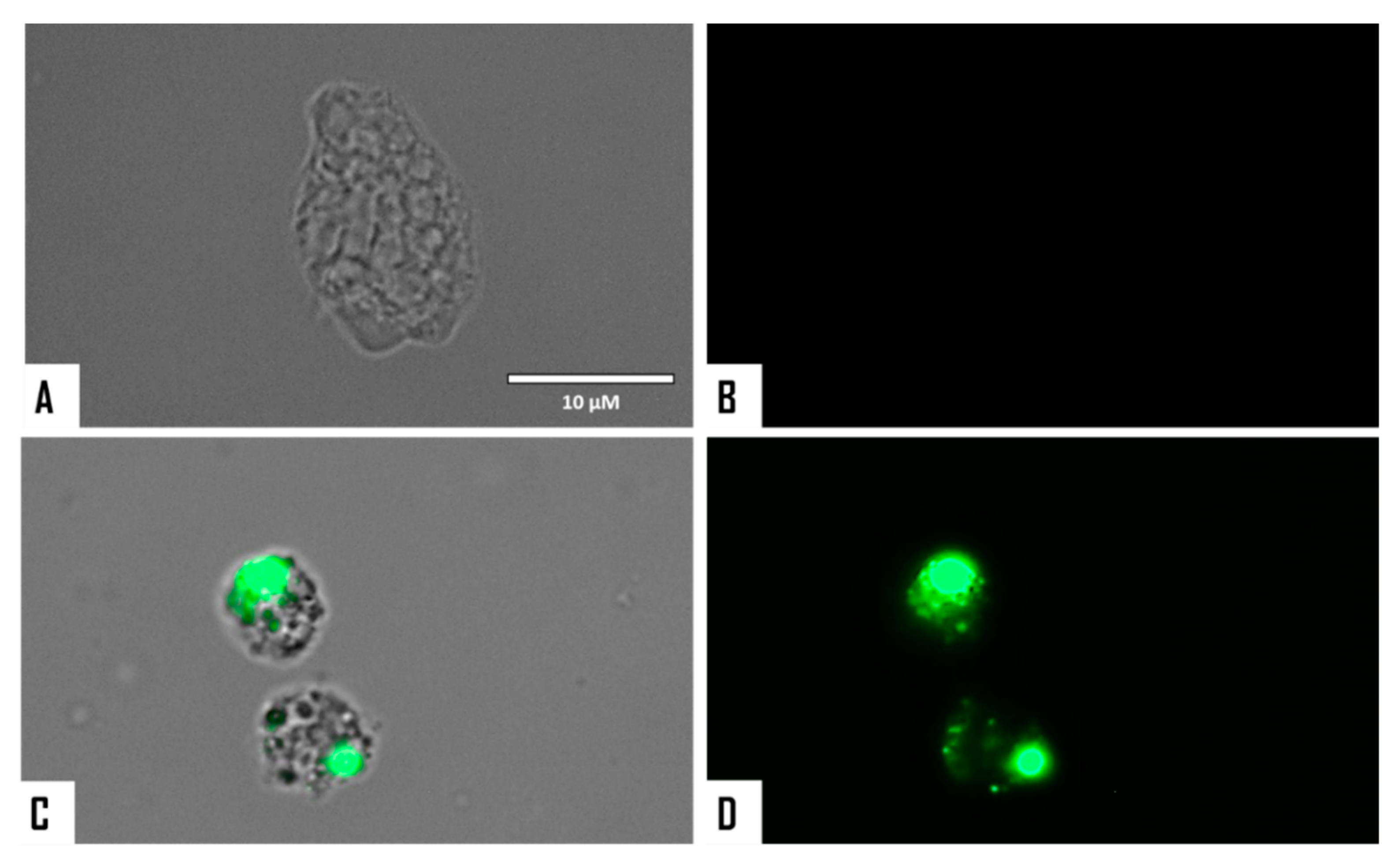
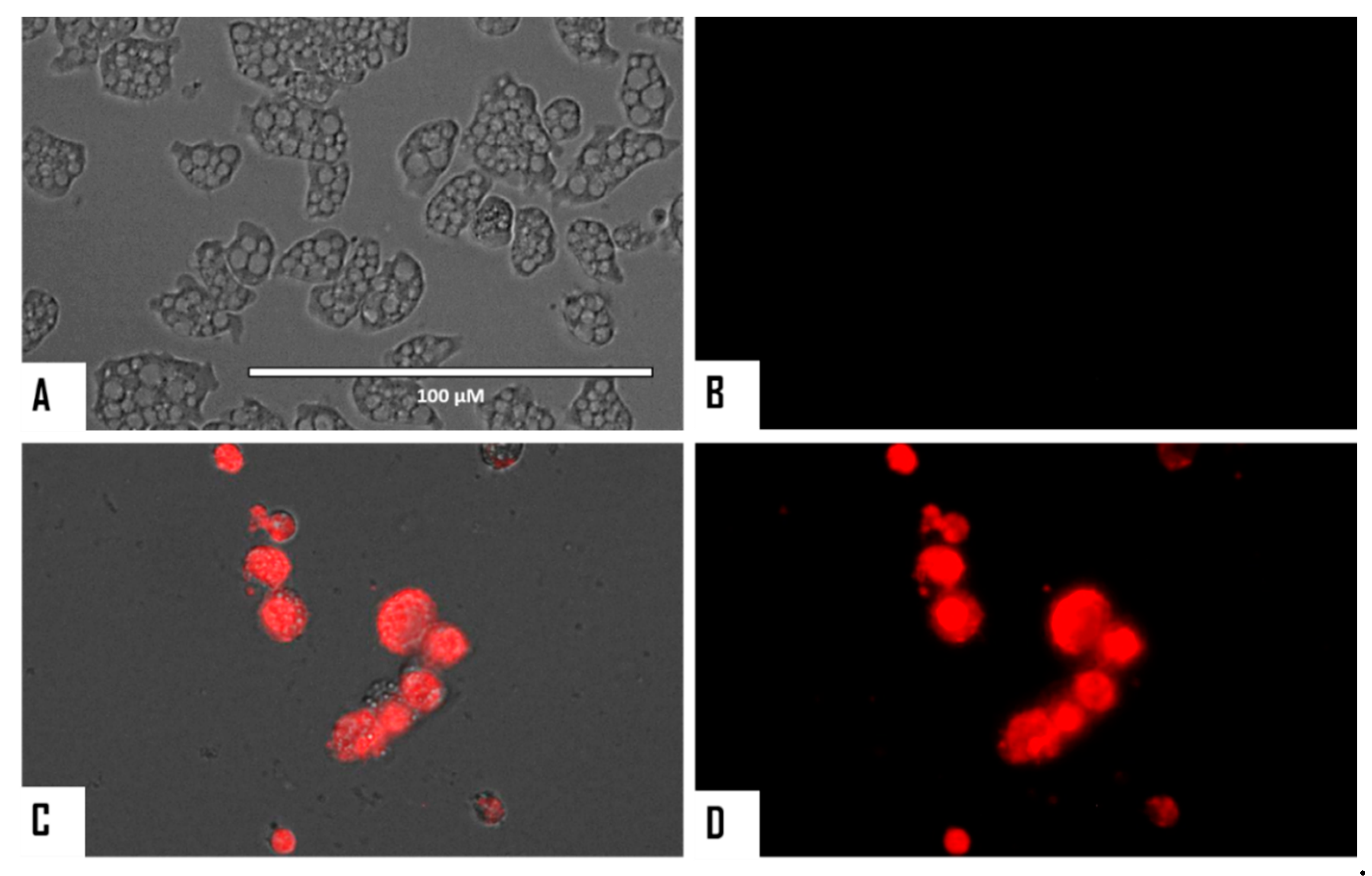
2.5. Evaluation of Programmed Cell Death (PCD) Induction in Both Naegleria fowleri Strains
2.5.1. Detection of DNA Condensation
2.5.2. Evaluation of Cellular Membrane Damage
2.5.3. Assessment of Mitochondrial Function Disruption in Treated Cells
ATP Detection Levels Using the Celltiter-Glo® Luminescent Cell Viability Assay
JC-1 Assay for the Detection of Alteration in the Mitochondrial Membrane Potential
2.6. Generation of Intracellular Reactive Oxygen Species (ROS)
3. Results
3.1. In Vitro Activity and Toxicity of the Tested Natural ICZs against Naegleria fowleri
3.2. Evaluation of PCD Induction in STS-Treated Amoebae
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, R.; Ali, I.K.M.; Cope, J.R.; Khan, N.A. Biology and pathogenesis of Naegleria fowleri. Acta Trop 2016, 164, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Pinero, J.E.; Chavez-Munguia, B.; Omana-Molina, M.; Lorenzo-Morales, J. Naegleria fowleri . Trends Parasitol. 2019, 35, 848–849. [Google Scholar] [CrossRef] [PubMed]
- Bellini, N.K.; Fonseca, A.L.M.D.; Reyes-Batlle, M.; Lorenzo-Morales, J.; Rocha, O.; Thiemann, O.H. Isolation of Naegleria spp. from a Brazilian water source. Pathogens 2020, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Visvesvara, G.S. Infections with free-living amebae. Handb. Clin. Neurol. 2013, 114, 153–168. [Google Scholar]
- Graciaa, D.S.; Cope, J.R.; Roberts, V.A.; Cikesh, B.L.; Kahler, A.M.; Vigar, M.; Hilborn, E.D.; Wade, T.J.; Backer, L.C.; Montgomery, S.P.; et al. Outbreaks associated with untreated recreational water-United States, 2000–2014. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 701–706. [Google Scholar] [CrossRef]
- Lopez, C.; Budge, P.; Chen, J.; Bilyeu, S.; Mirza, A.; Custodio, H.; Irazuzta, J.; Visvesvara, G.; Sullivan, K.J. Primary amebic meningoencephalitis: A case report and literature review. Pediatr. Emerg. Care 2012, 28, 272–276. [Google Scholar] [CrossRef]
- Cope, J.R.; Ali, I.K. Primary Amebic Meningoencephalitis: What have we learned in the last 5 Years? Curr. Infect. Dis. Rep. 2016, 18, 31–34. [Google Scholar] [CrossRef]
- Betanzos, A.; Banuelos, C.; Orozco, E. Host invasion by pathogenic amoebae: Epithelial Disruption by Parasite Proteins. Genes 2019, 10, 618. [Google Scholar] [CrossRef] [Green Version]
- Maciver, S.K.; Pinero, J.E.; Lorenzo-Morales, J. Is Naegleria fowleri an Emerging Parasite? Trends Parasitol. 2020, 36, 19–28. [Google Scholar] [CrossRef]
- Trabelsi, H.; Dendana, F.; Sellami, A.; Sellami, H.; Cheikhrouhou, F.; Neji, S.; Makni, F.; Ayadi, A. Pathogenic free-living amoebae: Epidemiology and clinical review. Pathol. Biol. (Paris) 2012, 60, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Jahangeer, M.; Mahmood, Z.; Munir, N.; Waraich, U.E.; Tahir, I.M.; Akram, M.; Ali Shah, S.M.; Zulfqar, A.; Zainab, R. Naegleria fowleri: Sources of infection, pathophysiology, diagnosis, and management; a review. Clin. Exp. Pharmacol. Physiol. 2020, 47, 199–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizo-Liendo, A.; Sifaoui, I.; Reyes-Batlle, M.; Chiboub, O.; Rodriguez-Exposito, R.L.; Bethencourt-Estrella, C.J.; San Nicolas-Hernandez, D.; Hendiger, E.B.; Lopez-Arencibia, A.; Rocha-Cabrera, P.; et al. In vitro activity of statins against Naegleria fowleri. Pathogens 2019, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Grace, E.; Asbill, S.; Virga, K. Naegleria fowleri: Pathogenesis, diagnosis, and treatment options. Antimicrob. Agents Chemother. 2015, 59, 6677–6681. [Google Scholar] [CrossRef] [Green Version]
- Capewell, L.G.; Harris, A.M.; Yoder, J.S.; Cope, J.R.; Eddy, B.A.; Roy, S.L.; Visvesvara, G.S.; Fox, L.M.; Beach, M.J. Diagnosis, clinical course, and treatment of Primary Amoebic Meningoencephalitis in the United States, 1937–2013. J. Pediatric Infect. Dis. Soc. 2015, 4, e68–e75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellini, N.K.; Santos, T.M.; da Silva, M.T.A.; Thiemann, O.H. The therapeutic strategies against Naegleria fowleri. Exp. Parasitol. 2018, 187, 1–11. [Google Scholar] [CrossRef]
- Linam, W.M.; Ahmed, M.; Cope, J.R.; Chu, C.; Visvesvara, G.S.; da Silva, A.J.; Qvarnstrom, Y.; Green, J. Successful treatment of an adolescent with Naegleria fowleri primary amebic meningoencephalitis. Pediatrics 2015, 135, E744–E748. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Davis, S.; Sifaoui, I.; Reyes-Batlle, M.; Viveros-Valdez, E.; Pinero, J.E.; Lorenzo-Morales, J.; Fernandez, J.J.; Diaz-Marrero, A.R. Anti-Acanthamoeba activity of brominated sesquiterpenes from Laurencia johnstonii. Mar. Drugs 2018, 16, 443. [Google Scholar] [CrossRef] [Green Version]
- Chiboub, O.; Sifaoui, I.; Lorenzo-Morales, J.; Abderrabba, M.; Mejri, M.; Fernandez, J.J.; Pinero, J.E.; Diaz-Marrero, A.R. Spiralyde A, an antikinetoplastid dolabellane from the brown alga Dictyota spiralis. Mar. Drugs 2019, 17, 192. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Marrero, A.R.; Lopez-Arencibia, A.; Bethencout-Estrella, C.J.; Cen-Pacheco, F.; Sifaoui, I.; Hernandez Creus, A.; Duque-Ramirez, M.C.; Souto, M.L.; Hernandez Daranas, A.; Lorenzo-Morales, J.; et al. Antiprotozoal activities of marine polyether triterpenoids. Bioorg. Chem. 2019, 92, 103276. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Diaz-Marrero, A.R.; Cen-Pacheco, F.; Sifaoui, I.; Reyes-Batlle, M.; Souto, M.L.; Hernandez Daranas, A.; Pinero, J.E.; Fernandez, J.J. Evaluation of oxasqualenoids from the red alga Laurencia viridis against Acanthamoeba. Mar. Drugs 2019, 17, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartuche, L.; Sifaoui, I.; Lopez-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolas-Hernandez, D.; Lorenzo-Morales, J.; Pinero, J.E.; Diaz-Marrero, A.R.; Fernandez, J.J. Antikinetoplastid activity of Indolocarbazoles from Streptomyces sanyensis. Biomolecules 2020, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Cartuche, L.; Reyes-Batlle, M.; Sifaoui, I.; Arberas-Jimenez, I.; Pinero, J.E.; Fernandez, J.J.; Lorenzo-Morales, J.; Diaz-Marrero, A.R. Antiamoebic activities of indolocarbazole metabolites isolated from Streptomyces sanyensis cultures. Mar. Drugs 2019, 17, 588. [Google Scholar] [CrossRef] [Green Version]
- Cartuche, L.; Sifaoui, I.; Cruz, D.; Reyes-Batlle, M.; Lopez-Arencibia, A.; Javier Fernandez, J.; Diaz-Marrero, A.R.; Pinero, J.E.; Lorenzo-Morales, J. Staurosporine from Streptomyces sanyensis activates programmed cell death in Acanthamoeba via the mitochondrial pathway and presents low in vitro cytotoxicity levels in a macrophage cell line. Sci. Rep. 2019, 9, 11651. [Google Scholar] [CrossRef] [PubMed]
- Sordet, O.; Khan, Q.A.; Plo, I.; Pourquier, P.; Urasaki, Y.; Yoshida, A.; Antony, S.; Kohlhagen, G.; Solary, E.; Saparbaev, M.; et al. Apoptotic topoisomerase I-DNA complexes induced by staurosporine-mediated oxygen radicals. J. Biol. Chem. 2004, 279, 50499–50504. [Google Scholar] [CrossRef] [Green Version]
- Sen, N.; Banerjee, B.; Das, B.B.; Ganguly, A.; Sen, T.; Pramanik, S.; Mukhopadhyay, S.; Majumder, H.K. Apoptosis is induced in leishmanial cells by a novel protein kinase inhibitor withaferin A and is facilitated by apoptotic topoisomerase I-DNA complex. Cell Death Differ. 2007, 14, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Braga, M.V.; de Souza, W. Effects of protein kinase and phosphatidylinositol-3 kinase inhibitors on growth and ultrastructure of Trypanosoma cruzi. FEMS Microbiol. Lett. 2006, 256, 209–216. [Google Scholar] [CrossRef]
- Osada, H.; Koshino, H.; Kudo, T.; Onose, R.; Isono, K. A new inhibitor of protein kinase C, RK-1409 (7-oxostaurosporine). I. Taxonomy and biological activity. J. Antibiot. (Tokyo) 1992, 45, 189–194. [Google Scholar] [CrossRef]
- Fabre, S.; Prudhomme, M.; Sancelme, M.; Rapp, M. Indolocarbazole protein kinase C inhibitors from rebeccamycin. Bioorg. Med. Chem. 1994, 2, 73–77. [Google Scholar] [CrossRef]
- Fabre, S.; Prudhomme, M.; Rapp, M. Protein kinase C inhibitors; structure-activity relationships in K252c-related compounds. Bioorg. Med. Chem. 1993, 1, 193–196. [Google Scholar] [CrossRef]
- Karaman, M.W.; Herrgard, S.; Treiber, D.K.; Gallant, P.; Atteridge, C.E.; Campbell, B.T.; Chan, K.W.; Ciceri, P.; Davis, M.I.; Edeen, P.T.; et al. A quantitative analysis of kinase inhibitor selectivity. Nat. Biotechnol. 2008, 26, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Bruges, G.; Betancourt, M.; March, M.; Sanchez, E.; Mijares, A. Apoptotic-like activity of staurosporine in axenic cultures of Trypanosoma evansi. Rev. Inst. Med. Trop Sao Paulo 2012, 54, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basmaciyan, L.; Berry, L.; Gros, J.; Azas, N.; Casanova, M. Temporal analysis of the autophagic and apoptotic phenotypes in Leishmania parasites. Microb. Cell 2018, 5, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.J.; Kang, J.S.; Byun, J.O.; Han, K.S.; Kim, D.U.; Oh, S.M.; Kim, H.M.; Chae, S.W.; Kim, H.R. Molecular mechanism of staurosporine-induced apoptosis in osteoblasts. Pharmacol. Res. 2000, 42, 373–381. [Google Scholar] [CrossRef]
- Sifaoui, I.; Lopez-Arencibia, A.; Martin-Navarro, C.M.; Reyes-Batlle, M.; Mejri, M.; Valladares, B.; Lorenzo-Morales, J.; Abderabba, M.; Pinero, J.E. Selective activity of oleanolic and maslinic acids on the amastigote form of Leishmania spp. Iran. J. Pharm. Res. 2017, 16, 1190–1193. [Google Scholar]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- Sifaoui, I.; Rodriguez-Exposito, R.L.; Reyes-Batlle, M.; Rizo-Liendo, A.; Pinero, J.E.; Bazzocchi, I.L.; Lorenzo-Morales, J.; Jimenez, I.A. Ursolic acid derivatives as potential agents against Acanthamoeba spp. Pathogens 2019, 8, 130. [Google Scholar] [CrossRef] [Green Version]
- Janosik, T.; Rannug, A.; Rannug, U.; Wahlstrom, N.; Slatt, J.; Bergman, J. Chemistry and properties of indolocarbazoles. Chem. Rev. 2018, 118, 9058–9128. [Google Scholar] [CrossRef]
- Gani, O.A.; Engh, R.A. Protein kinase inhibition of clinically important staurosporine analogues. Nat. Prod. Rep. 2010, 27, 489–498. [Google Scholar] [CrossRef]
- Pereira, E.R.; Belin, L.; Sancelme, M.; Prudhomme, M.; Ollier, M.; Rapp, M.; Severe, D.; Riou, J.F.; Fabbro, D.; Meyer, T. Structure-activity relationships in a series of substituted indolocarbazoles: Topoisomerase I and protein kinase C inhibition and antitumoral and antimicrobial properties. J. Med. Chem. 1996, 39, 4471–4477. [Google Scholar] [CrossRef]
- Lawrie, A.M.; Noble, M.E.; Tunnah, P.; Brown, N.R.; Johnson, L.N.; Endicott, J.A. Protein kinase inhibition by staurosporine revealed in details of the molecular interaction with CDK2. Nat. Struct. Biol. 1997, 4, 796–801. [Google Scholar] [CrossRef] [PubMed]
- John, D.T.; John, R.A. Viability of pathogenic Acanthamoeba and Naegleria and virulence of N. fowleri during long-term cryopreservation. Folia Parasitol. (Praha) 1996, 43, 43–46. [Google Scholar] [PubMed]
- John, D.T.; John, R.A. Viability of pathogenic Naegleria and Acanthamoeba isolates during 10 years of cryopreservation. Folia Parasitol. (Praha) 2006, 53, 311–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumidonming, W.; Koehsler, M.; Walochnik, J. Acanthamoeba strains show reduced temperature tolerance after long-term axenic culture. Parasitol. Res. 2010, 106, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.M.; Ferguson, T.J.; Marciano-Cabral, F. Protein kinase activation and protein phosphorylation in Naegleria fowleri amebae in response to normal human serum. J. Eukaryot. Microbiol. 2000, 47, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Han, K.L.; Lee, H.J.; Shin, M.H.; Shin, H.J.; Im, K.I.; Park, S.J. The involvement of an integrin-like protein and protein kinase C in amoebic adhesion to fibronectin and amoebic cytotoxicity. Parasitol. Res. 2004, 94, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Jamerson, M.; Schmoyer, J.A.; Park, J.; Marciano-Cabral, F.; Cabral, G.A. Identification of Naegleria fowleri proteins linked to primary amoebic meningoencephalitis. Microbiology 2017, 163, 322–332. [Google Scholar] [CrossRef]
- Fritz-Laylin, L.K.; Prochnik, S.E.; Ginger, M.L.; Dacks, J.B.; Carpenter, M.L.; Field, M.C.; Kuo, A.; Paredez, A.; Chapman, J.; Pham, J.; et al. The genome of Naegleria gruberi illuminates early eukaryotic versatility. Cell 2010, 140, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Debnath, A.; Tunac, J.B.; Galindo-Gomez, S.; Silva-Olivares, A.; Shibayama, M.; McKerrow, J.H. Corifungin, a new drug lead against Naegleria, identified from a high-throughput screen. Antimicrob. Agents Chemother. 2012, 56, 5450–5457. [Google Scholar] [CrossRef] [Green Version]
- Kaczanowski, S.; Sajid, M.; Reece, S.E. Evolution of apoptosis-like programmed cell death in unicellular protozoan parasites. Parasit Vectors 2011, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Cardenas-Zuniga, R.; Silva-Olivares, A.; Villalba-Magdaleno, J.A.; Sanchez-Monroy, V.; Serrano-Luna, J.; Shibayama, M. Amphotericin B induces apoptosis-like programmed cell death in Naegleria fowleri and Naegleria grubceri. Microbiology 2017, 163, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Sherer, C.; Snape, T.J. Heterocyclic scaffolds as promising anticancer agents against tumours of the central nervous system: Exploring the scope of indole and carbazole derivatives. Eur. J. Med. Chem. 2015, 97, 552–560. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
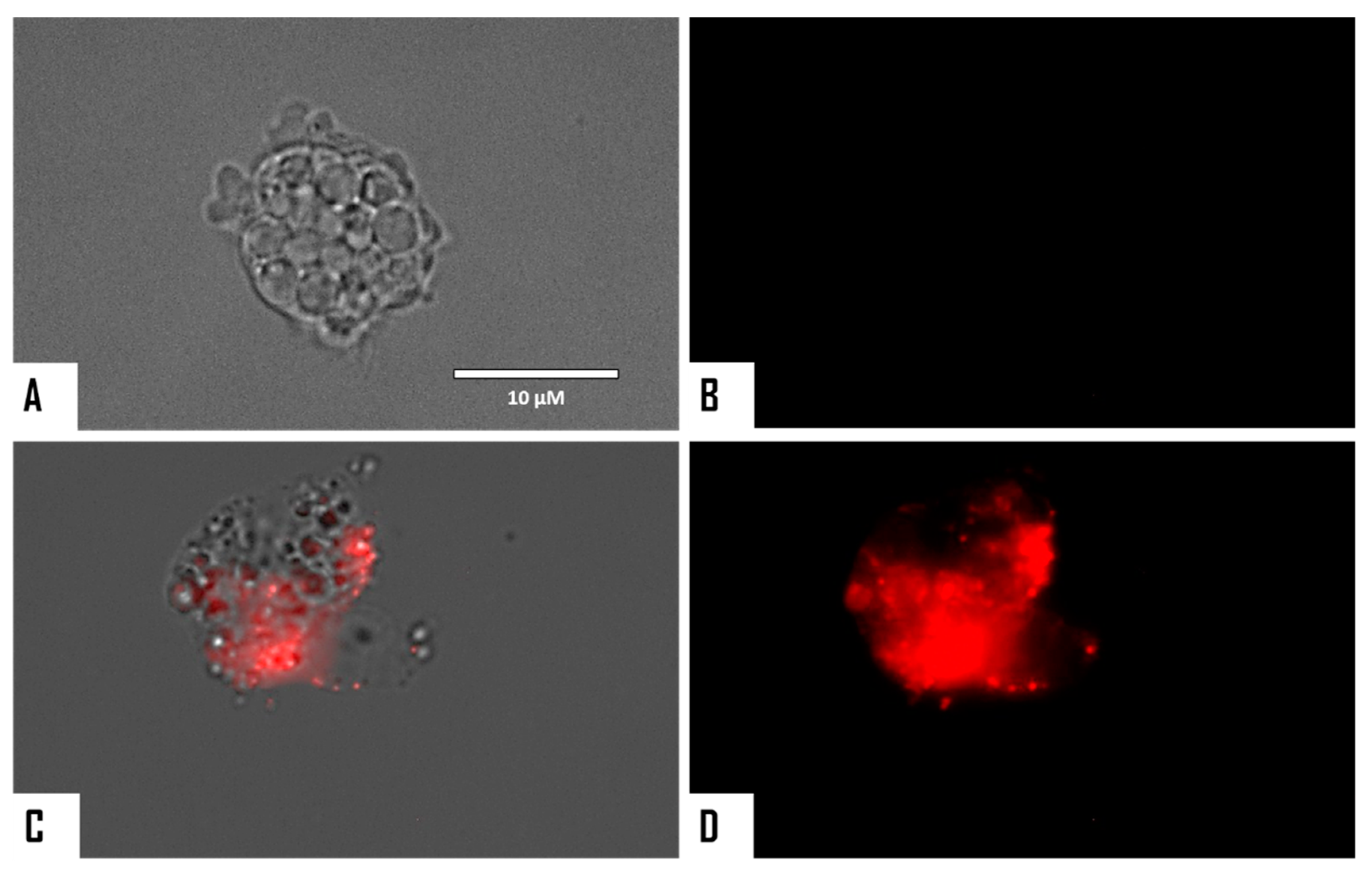{kind=link}
{kind=link}
| Compound | N. fowleri ATCC® 30808™ IC50 (µM) | N. fowleri ATCC® 30215™ IC50 (µM) | Murine Macrophage J774.A1 CC50 (µM) | SI * (CC50/IC50) | Log P ** |
|---|---|---|---|---|---|
| STS (1) | 0.08 *** ± 0.01 | 0.08 **** ± 0.02 | 8.74 ± 0.72 | 109.3 | 3.86 |
| 7OSTS (2) | 1.28 ± 0.35 | 1.07 ± 0.23 | 5.20 ± 1.75 | 4.1 | 3.74 |
| 4′D4′OSTS (3) | 10.79 ± 2.92 | NA | >20 | >1.9 | 3.38 |
| SCZ B (4) | 11.51 ± 0.31 | NA | >20 | >1.7 | 3.43 |
| Amphotericin B | 0.12 ± 0.03 | 0.16 ± 0.02 | ≥200 | ≥1666.7 | - |
| Miltefosine | 38.74 ± 4.23 | 81.57 ± 7.23 | 127.88 ± 8.85 | 3.3 | - |
| Compound | N. fowleri ATCC® 30808™ IC50 (µM) | N. fowleri ATCC® 30215™ IC50 (µM) | Murine Macrophage J774.A1 CC50 (µM) | SI * (CC50/IC50) | Log P ** |
|---|---|---|---|---|---|
| Rebeccamycin (5) | NA at 20 | - | 1.42 ± 0.19 | - | 2.57 |
| K252a (6) | 0.34 ± 0.09 | 0.12 ± 0.03 | 1.07 ± 0.21 | 3.2 | 3.66 |
| K252b (7) | NA at 20 | - | ≥20 | - | 3.39 |
| K252c (8) | NA at 20 | - | 35.40 ± 2.47 | - | 3.09 |
| Arcyriflavin A (9) | NA at 20 | - | ≥20 | - | 2.96 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizo-Liendo, A.; Sifaoui, I.; Cartuche, L.; Arberas-Jiménez, I.; Reyes-Batlle, M.; Fernández, J.J.; Piñero, J.E.; Díaz-Marrero, A.R.; Lorenzo-Morales, J. Evaluation of Indolocarbazoles from Streptomyces sanyensis as a Novel Source of Therapeutic Agents against the Brain-Eating Amoeba Naegleria fowleri. Microorganisms 2020, 8, 789. https://doi.org/10.3390/microorganisms8050789
Rizo-Liendo A, Sifaoui I, Cartuche L, Arberas-Jiménez I, Reyes-Batlle M, Fernández JJ, Piñero JE, Díaz-Marrero AR, Lorenzo-Morales J. Evaluation of Indolocarbazoles from Streptomyces sanyensis as a Novel Source of Therapeutic Agents against the Brain-Eating Amoeba Naegleria fowleri. Microorganisms. 2020; 8(5):789. https://doi.org/10.3390/microorganisms8050789
Chicago/Turabian StyleRizo-Liendo, Aitor, Ines Sifaoui, Luis Cartuche, Iñigo Arberas-Jiménez, María Reyes-Batlle, José J. Fernández, José E. Piñero, Ana R. Díaz-Marrero, and Jacob Lorenzo-Morales. 2020. "Evaluation of Indolocarbazoles from Streptomyces sanyensis as a Novel Source of Therapeutic Agents against the Brain-Eating Amoeba Naegleria fowleri" Microorganisms 8, no. 5: 789. https://doi.org/10.3390/microorganisms8050789