Actinomycetes from the Red Sea Sponge Coscinoderma mathewsi: Isolation, Diversity, and Potential for Bioactive Compounds Discovery

 ,
,  , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Area for Sponge Collection
2.2. Chemicals and Reagents
2.3. Actinomycetes Isolation
2.4. Molecular Identification and Phylogenetic Analysis
2.5. Co-cultivation and Extract Preparation
2.6. Metabolic Profiling
2.7. Mycolic Acid Detection
2.8. Antibacterial Activity
2.9. Antifungal Activity
2.10. Anti-Trypanosomal Activity
2.11. Statistical Analysis
3. Results and Discussion
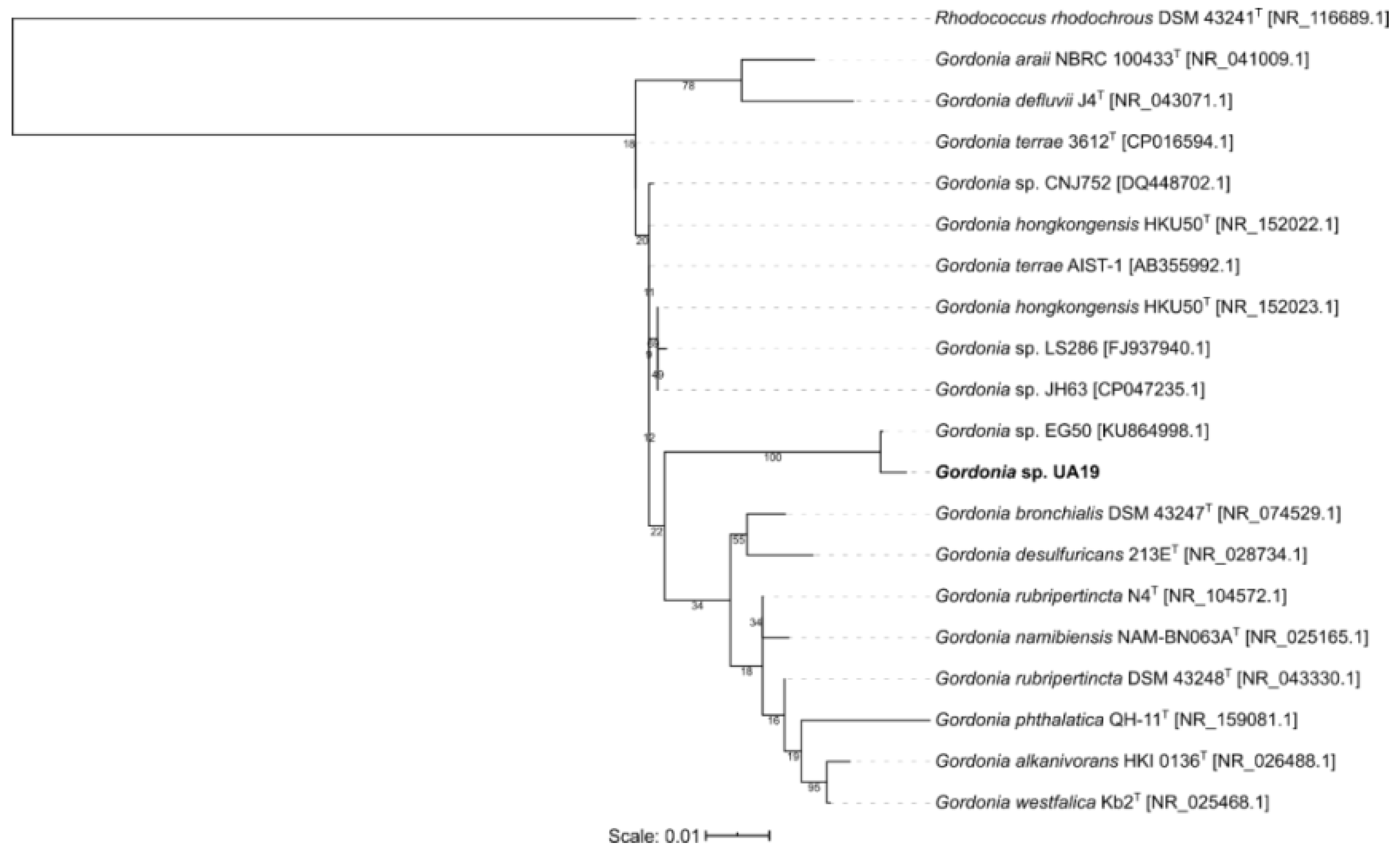
3.1. Molecular Identification and Phylogenetic Analysis
3.2. Metabolomic Profiling of Monoculture and Co-Culture Crude Extracts
3.2.1. Chemical Dereplication of Micromonospora sp. UA17
3.2.2. Chemical Dereplication of Gordonia sp. UA19
3.2.3. Chemical Dereplication of Nocardia sp. UA 23
3.2.4. Chemical Dereplication of Strains UA17 + UA19
3.2.5. Chemical Dereplication of Strains UA17 + UA23
3.2.6. Chemical Dereplication of Strain UA17 with Mycolic Acid
3.3. Antibacterial, Antifungal, and Anti-Trypanosomal Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hameş-Kocabaş, E.E.; Ataç, U.Z.E.L. Isolation strategies of marine-derived actinomycetes from sponge and sediment samples. J. Microbiol. Methods 2012, 88, 342–347. [Google Scholar]
- Adegboye, M.F.; Babalola, O.O. Taxonomy and ecology of antibiotic producing actinomycetes. Afr. J. Agric. Res. 2012, 15, 2255–2261. [Google Scholar]
- Zhao, K.; Penttinen, P.; Guan, T.W.; Xiao, J.; Chen, Q.A.; Xu, J.; Lindstrom, K.; Zhang, L.L.; Zhang, X.P.; Strobel, G.A. The diversity and antimicrobial activity of endophytic actinomycetes isolated from medicinal plants in Panxi Plateau China. Curr. Microbiol. 2011, 62, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukhari, M.; Thomas, A.; Wong, N. Culture Conditions for Optimal Growth of Actinomycetes from Marine Sponges. In Developments in Sustainable Chemical and Bioprocess Technology; Springer: New York, NY, USA, 2013; pp. 203–210. [Google Scholar] [CrossRef]
- Vicente, J.; Stewart, A.; Song, B.; Hill, R.T.; Wright, J.L. Biodiversity of Actinomycetes associated with Caribbean sponges and their potential for natural product discovery. Mar. Biotechnol. 2013, 15, 413–424. [Google Scholar] [CrossRef]
- Sun, W.; Peng, C.S.; Zhao, Y.Y.; Li, Z.Y. Functional gene–guided discovery of type II polyketides from culturable actinomycetes associated with soft coral Scleronephthya sp. PLoS ONE 2012, 7, e42847. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; MacIntyre, L.; Abdelmohsen, U.R.; Horn, H.; Polymenakou, P.; Edrada-Ebel, R.; Hentschel, U. Biodiversity, anti–trypanosomal activity screening, and metabolomics profiling of actinomycetes isolated from Mediterranean sponges. PLoS ONE 2015, 10, e0138528. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Bayer, K.; Hentschel, U. Diversity, abundance, and natural products of marine sponge–associated actinomycetes. Nat. Prod. Rep. 2014, 31, 381–399. [Google Scholar] [CrossRef]
- Muller, R.; Wink, J. Future potential for anti–infectives from bacteria–how to exploit biodiversity and genomic potential. Int. J. Med. Microbiol. 2014, 304, 3–13. [Google Scholar] [CrossRef]
- Dalisay, D.S.; Williams, D.E.; Wang, X.L.; Centko, R.; Chen, J.; Andersen, R.J. Marine sediment–derived Streptomyces bacteria from British Columbia, Canada are a promising microbiota resource for the discovery of antimicrobial natural products. PLoS ONE 2013, 8, e77078. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Abdelmohsen, U.R.; Ibrahim, A.K.; Hassanean, H.A.; Hentschel, U.; Ahmed, S.A. New antibacterial xanthone from the marine sponge–derived Micrococcus sp. EG45. Bioorg. Med. Chem. Lett. 2014, 24, 4939–4942. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Zhang, G.L.; Philippe, A.; Schmitz, W.; Pimentel-Elardo, S.M.; Hertlein-Amslinger, B.; Hentschel, U.; Bringmann, G. Cyclodysidins A–D, cyclic lipopeptides from the marine sponge–derived Streptomyces strain RV15. Tetrahedron Lett. 2012, 53, 23–29. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Szesny, M.; Othman, E.M.; Schirmeister, T.; Grond, S.; Stopper, H.; Hentschel, U. Antioxidant and anti–Protease activities of diazepinomicin from the sponge–associated Micromonospora strain RV115. Mar. Drugs 2012, 10, 2208–2221. [Google Scholar] [CrossRef] [PubMed]
- Subramani, R.; Aalbersberg, W. Marine actinomycetes: An ongoing source of novel bioactive metabolites. Microbiol. Res. 2012, 167, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Solanki, R.; Khanna, M.; Lal, R. Bioactive compounds from marine actinomycetes. Indian J. Microbiol. 2008, 48, 410–431. [Google Scholar] [CrossRef] [Green Version]
- Abdelmohsen, U.R.; Yang, C.; Horn, H.; Hajjar, D.; Ravasi, T.; Hentschel, U. Actinomycetes from Red Sea sponges: Sources for chemical and phylogenetic diversity. Mar. Drugs 2014, 12, 2771–2789. [Google Scholar] [CrossRef] [Green Version]
- Grkovic, T.; Abdelmohsen, U.R.; Othman, E.M.; Stopper, H.; Edrada-Ebel, R.; Hentschel, U.; Quinn, R.J. Two new antioxidant actinosporin analogues from the calcium alginate beads culture of sponge associated Actinokineospora sp. strain EG49. Bioorg. Med. Chem. Lett. 2014, 24, 5089–5092. [Google Scholar] [CrossRef]
- Ziemert, N.; Lechner, A.; Wietz, M.; Millan-Aguinaga, N.; Chavarria, K.L.; Jensen, P.R. Diversity, and evolution of secondary metabolism in the marine actinomycete genus Salinispora. Proc. Natl. Acad. Sci. USA 2014, 111, 1130–1139. [Google Scholar] [CrossRef] [Green Version]
- Udwary, D.W.; Zeigler, L.; Asolkar, R.N.; Singan, V.; Lapidus, A.; Fenical, W.; Jensen, P.R.; Moore, B.S. Genome sequencing reveals complex secondary metabolome in the marine actinomycete Salinispora tropica. Proc. Natl. Acad. Sci. USA 2007, 104, 10376–10381. [Google Scholar] [CrossRef] [Green Version]
- Marmann, A.; Aly, A.H.; Lin, W.H.; Wang, B.G.; Proksch, P. Co–Cultivation–A Powerful Emerging Tool for Enhancing the Chemical Diversity of Microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [Green Version]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Brown, L.C.W.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Abdelmohsen, U.R.; Grkovic, T.; Balasubramanian, S.; Kamel, M.S.; Quinn, R.J.; Hentschel, U. Elicitation of secondary metabolism in actinomycetes. Biotechnol. Adv. 2015, 33, 798–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.Q.; Tan, H.R. Molecular regulation of antibiotic biosynthesis in streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochi, K.; Hosaka, T. New strategies for drug discovery: Activation of silent or weakly expressed microbial gene clusters. Appl. Microbiol. Biotechnol. 2013, 97, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef]
- Letzel, A.C.; Pidot, S.J.; Hertweck, C. A genomic approach to the cryptic secondary metabolome of the anaerobic world. Nat. Prod. Rep. 2013, 30, 392–428. [Google Scholar] [CrossRef]
- Luo, Y.; Huang, H.; Liang, J.; Wang, M.; Lu, L.; Shao, Z.; Cobb, R.E.; Zhao, H. Activation, and characterization of a cryptic polycyclic tetramate macrolactam biosynthetic gene cluster. Nat. Commun. 2013, 4, 2894. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Sandiford, S.K.; van Wezel, G.P. Triggers and cues that activate antibiotic production by actinomycetes. J. Ind. Microbiol. Biotechnol. 2014, 41, 371–386. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Hofs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Paranagama, P.A.; Wijeratne, E.M.K.; Gunatilaka, A.A.L. Uncovering biosynthetic potential of plant–associated fungi: Effect of culture conditions on metabolite production by Paraphaeosphaeria quadriseptata and Chaetomium chiversii. J. Nat. Prod. 2007, 70, 1939–1945. [Google Scholar] [CrossRef]
- Wei, H.; Lin, Z.; Li, D.; Gu, Q.; Zhu, T. OSMAC (One Strain Many Compounds) approach in the research of microbial metabolites a review. Wei Sheng Wu Xue Bao 2010, 50, 701–709. [Google Scholar] [PubMed]
- Abdelmohsen, U.R.; Cheng, C.; Viegelmann, C.; Zhang, T.; Grkovic, T.; Ahmed, S.; Quinn, R.J.; Hentschel, U.; Edrada-Ebel, R. Dereplication strategies for targeted isolation of new antitrypanosomal actinosporins A and B from a marine sponge associated–Actinokineospora sp. EG49. Mar. Drugs 2014, 12, 1220–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, W.R.; Guthertz, L.S. Mycolic acid analysis by high–performance liquid chromatography for identification of Mycobacterium species. Clin. Microbiol. Rev. 2001, 14, 704–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Betancourt, O.E.; Karls, R.; Grosse-Siestrup, B.; Helms, S.; Quinn, F.; Dluhy, R.A. Identification of mycobacteria based on spectroscopic analyses of mycolic acid profiles. Analyst 2013, 138, 6774–6785. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.R.; Floyd, M.M.; Brown, J.M.; Toney, S.R.; Daneshvar, M.I.; Cooksey, R.C.; Carr, J.; Steigerwalt, A.G.; Charles, N. Novel mycolic acid–containing bacteria in the family Segniliparaceae fam. nov., including the genus Segniliparus gen. nov., with descriptions of Segniliparus rotundus sp nov and Segniliparus rugosus sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1615–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrakchi, H.; Laneelle, M.A.; Daffe, M. Mycolic Acids: Structures, Biosynthesis, and Beyond. Chem. Biolog. 2014, 21, 67–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamet, S.; Slama, N.; Domingues, J.; Laval, F.; Texier, P.; Eynard, N.; Quemard, A.; Peixoto, A.; Lemassu, A.; Daffe, M.; et al. The Non–Essential Mycolic Acid Biosynthesis Genes hadA and hadC Contribute to the Physiology and Fitness of Mycobacterium smegmatis. PLoS ONE 2015, 10, e0145883. [Google Scholar] [CrossRef]
- Glickman, M.S.; Cox, J.S.; Jacobs, W.R. A novel mycolic acid cyclopropane synthetase is required for cording, persistence, and virulence of Mycobacterium tuberculosis. Mol. Cell 2000, 5, 717–727. [Google Scholar] [CrossRef]
- Onaka, H.; Mori, Y.; Igarashi, Y.; Furumai, T. Mycolic Acid–Containing Bacteria Induce Natural–Product Biosynthesis in Streptomyces Species. Appl. Environ. Microbiol. 2011, 77, 400–406. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, S.; Okada, M.; Wakimoto, T.; Zhang, H.; Hayashi, F.; Onaka, H.; Abe, I. Niizalactams A–C, Multicyclic Macrolactams Isolated from Combined Culture of Streptomyces with Mycolic Acid–Containing Bacterium. J. Nat. Prod. 2015, 78, 3011–3017. [Google Scholar] [CrossRef]
- Hoshino, S.; Zhang, L.; Awakawa, T.; Wakimoto, T.; Onaka, H.; Abe, I. Arcyriaflavin E, a new cytotoxic indolocarbazole alkaloid isolated by combined–culture of mycolic acid–containing bacteria and Streptomyces cinnamoneus Nbrc 13823. J. Antibiotics 2015, 68, 342–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, S.; Wakimoto, T.; Onaka, H.; Abe, I. Chojalactones A–C, cytotoxic butanolides isolated from Streptomyces sp. cultivated with mycolic acid–containing bacterium. Org. Lett. 2015, 17, 1501–1504. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high–throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post–analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Tawfike, A.; Attia, E.Z.; Desoukey, S.Y.; Hajjar, D.; Makki, A.A.; Schupp, P.J.; Edrada-Ebel, R.; Abdelmohsen, U.R. New bioactive metabolites from the elicited marine sponge–derived bacterium Actinokineospora spheciospongiae sp. nov. AMB Express 2019, 9, 12. [Google Scholar] [CrossRef]
- Liu, M.; Lu, J.; Müller, P.; Turnbull, L.; Burke, C.M.; Schlothauer, R.C.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Antibiotic–specific differences in the response of Staphylococcus aureus to treatment with antimicrobials combined with manuka honey. Fron. Microbiol. 2015, 5, 779. [Google Scholar] [CrossRef] [Green Version]
- Lum, K.Y.; Tay, S.T.; Le, C.F.; Lee, V.S.; Sabri, N.H.; Velayuthan, R.D.; Hassan, H.; Sekaran, S.D. Activity of novel synthetic peptides against Candida albicans. Sci. Rep. 2015, 5, 9657. [Google Scholar] [CrossRef]
- Kaewkla, O.; Thamchaipinet, A.; Franco, C.M.M. Micromonospora terminaliae sp. nov., an endophytic actinobacterium isolated from the surface–sterilized stem of the medicinal plant Terminalia mucronata. Int. J. Syst. Evol. Microbiol. 2017, 67, 225–230. [Google Scholar] [CrossRef]
- Carro, L.; Nouioui, I.; Sangal, V.; Meier-Kolthoff, J.P.; Trujillo, M.E.; del Carmen Montero-Calasanz, M.; Sahin, N.; Smith, D.L.; Kim, K.E.; Peluso, P. Genome–based classification of micromonosporae with a focus on their biotechnological and ecological potential. Sci. Rep. 2018, 8, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Carro, L.; Pukall, R.; Spröer, C.; Kroppenstedt, R.M.; Trujillo, M.E. Micromonospora cremea sp. nov. and Micromonospora zamorensis sp. nov., isolated from the rhizosphere of Pisum sativum. Int. J. Syst. Evol. Microbiol. 2012, 62, 2971–2977. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H.; Tamura, T.; Harayama, S. Intrageneric relationships among Micromonospora species deduced from gyrB–based phylogeny and DNA relatedness. Int. J. Syst. Evol. Microbiol. 2000, 50, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, B.; Liu, C.; Guan, X.; Song, J.; Zhao, J.; Liu, H.; Li, C.; Ning, W.; Wang, X.; Xiang, W. Two new species of the genus Micromonospora: Micromonospora palomenae sp. nov. and Micromonospora harpali sp. nov. isolated from the insects. Antonie Van Leeuwenhoek 2015, 108, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.-C.; Xiong, L.; Poon, R.W.; Chen, J.H.; Leung, K.-W.; Lam, J.Y.; Wu, A.K.; Chan, J.F.; Lau, S.K.; Woo, P.C. Gordonia hongkongensis sp. nov., isolated from blood culture and peritoneal dialysis effluent of patients in Hong Kong. Int. J. Syst. Evol. Microbiol. 2016, 66, 3942–3950. [Google Scholar] [CrossRef]
- Russell, D.A.; Bustamante, C.A.G.; Garlena, R.A.; Hatfull, G.F. Complete genome sequence of Gordonia terrae 3612. Genome Announc. 2016, 4, e01058-16. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, N.; Sikorski, J.; Jando, M.; Lapidus, A.; Nolan, M.; Lucas, S.; Del Rio, T.G.; Tice, H.; Copeland, A.; Cheng, J.-F. Complete genome sequence of Gordonia bronchialis type strain (3410 T). Stand. Genomic Sci. 2010, 2, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.B.; Brown, R.; Oldfield, C.; Gilbert, S.C.; Iliarionov, S.; Goodfellow, M. Gordonia amicalis sp. nov., a novel dibenzothiophene–desulphurizing actinomycete. Int. J. Syst. Evol. Microbiol. 2000, 50, 2031–2036. [Google Scholar] [CrossRef] [Green Version]
- Shen, F.-T.; Lu, H.-L.; Lin, J.-L.; Huang, W.-S.; Arun, A.; Young, C.-C. Phylogenetic analysis of members of the metabolically diverse genus Gordonia based on proteins encoding the gyrB gene. Res. Microbiol. 2006, 157, 367–375. [Google Scholar] [CrossRef]
- Thawai, C.; Rungjindamai, N.; Klanbut, K.; Tanasupawat, S. Nocardia xestospongiae sp. nov., isolated from a marine sponge in the Andaman Sea. Int. J. Syst. Evol. Microbiol. 2017, 67, 1451–1456. [Google Scholar] [CrossRef]
- Ezeoke, I.; Klenk, H.-P.; Pötter, G.; Schumann, P.; Moser, B.D.; Lasker, B.A.; Nicholson, A.; Brown, J.M. Nocardia amikacinitolerans sp. nov., an amikacin–resistant human pathogen. Int. J. Syst. Evol. Microbiol. 2013, 63, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Conville, P.S.; Zelazny, A.M.; Witebsky, F.G. Analysis of secA1 gene sequences for identification of Nocardia species. J. Clin. Microbiol. 2006, 44, 2760–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conville, P.S.; Murray, P.R.; Zelazny, A.M. Evaluation of the Integrated Database Network System (IDNS) SmartGene software for analysis of 16S rRNA gene sequences for identification of Nocardia species. J. Clin. Microbiol. 2010, 48, 2995–2998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nett, M.; Ikeda, H.; Moore, B.S. Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep. 2009, 26, 1362–1384. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Sayed, A.M.; Mohammed, R.; Hassan, H.M.; Zaki, M.A.; Rateb, M.E.; Mohammed, T.A.; Amin, E.; Abdelmohsen, U.R. Epigenetic Modifiers Induce Bioactive Phenolic Metabolites in the Marine–Derived Fungus Penicillium brevicompactum. Mar. Drugs 2018, 16, 253. [Google Scholar] [CrossRef] [Green Version]
- Dashti, Y.; Grkovic, T.; Abdelmohsen, U.R.; Hentschel, U.; Quinn, R.J. Actinomycete Metabolome Induction/Suppression with N–Acetylglucosamine. J. Nat. Prod. 2017, 80, 828–836. [Google Scholar] [CrossRef]
- Dinesh, R.; Srinivasan, V.; Sheeja, T.E.; Anandaraj, M.; Srambikkal, H. Endophytic actinobacteria: Diversity, secondary metabolism, and mechanisms to unsilence biosynthetic gene clusters. Crit. Rev. Microbiol. 2017, 43, 546–566. [Google Scholar] [CrossRef]
- Fujioka, K.; Furihata, K.; Shimazu, A.; Hayakawa, Y.; Seto, H. Isolation and characterization of atramycin A and atramycin B, new isotetracenone type antitumor antibiotics. J. Antibiot. 1991, 44, 1025–1028. [Google Scholar] [CrossRef]
- Cai, Y.; Fredenhagen, A.; Hug, P.; Peter, H.H. A nitro analogue of staurosporine and other minor metabolites produced by a Streptomyces longisporoflavus strain. J. Antibiot. 1995, 48, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Maskey, R.P.; Grün-Wollny, I.; Laatsch, H. Resomycins AC: New Anthracyclinone Antibiotics Formed by a Terrestrial Streptomyces sp. J. Antibiot. 2003, 56, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Jin, W. Isolation and structure determination of mutactimycin A, a new anthracycline antibiotic. Kangshengsu 1990, 15, 399–406. [Google Scholar]
- Osada, H.; Koshino, H.; Kudo, T.; Onose, R.; Isono, K. A new inhibitor of protein kinase C, Rk–1409 (7–oxostaurosporine). J. Antibiot. 1992, 45, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Harindran, J.; Varde, A. Pimprinine, an extracellular alkaloid produced by Streptomyces CDRIL–312: Fermentation, isolation, and pharmacological activity. J. Biotechnol. 2001, 88, 1–10. [Google Scholar] [CrossRef]
- Wei, Y.; Fang, W.; Wan, Z.; Wang, K.; Yang, Q.; Cai, X.; Shi, L.; Yang, Z. Antiviral effects against EV71 of pimprinine and its derivatives isolated from Streptomyces sp. Virol. J. 2014, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.E.; Davies, J.; Patrick, B.O.; Bottriell, H.; Tarling, T.; Roberge, M.; Andersen, R.J. Cladoniamides A− G, tryptophan–derived alkaloids produced in culture by Streptomyces uncialis. Org. Lett. 2008, 10, 3501–3504. [Google Scholar] [CrossRef] [PubMed]
- Roscales, S.; Plumet, J. Biosynthesis and biological activity of carbasugars. Int. J. Carbohydr. Chem. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.Q.; Maul, C.; Höfs, R.; Sattler, I.; Grabley, S.; Feng, X.Z.; Zeeck, A.; Thiericke, R. Gabosines L, N and O: New Carba-Sugars from Streptomyces with DNA-Binding Properties. Eur. J. Org. Chem. 2000, 149–153. [Google Scholar] [CrossRef]
- Balitz, D.; Bush, J.; Bradner, W.; Doyle, T.; O’herron, F.; Nettleton, D. Isolation of lavendamycin a new antibiotic from Streptomyces lavendulae. J. Antibiot. 1982, 35, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, E.; Beppu, T. Anew nucleosidic antibiotic AT–265. J. Antibiot. 1982, 35, 939–947. [Google Scholar] [CrossRef] [Green Version]
- Ates, N.; Ilbay, G.; Sahin, D. Suppression of generalized seizures activity by intrathalamic 2–chloroadenosine application. Exp. Biol. Med. 2005, 230, 501–505. [Google Scholar] [CrossRef]
- Takeuchi, M.; Inukai, M.; Enokita, R.; Iwado, S.; Takahashi, S.; Arai, M. Malioxamycin, a new antibiotic with spheroplast–forming activity. J. Antibiot. 1980, 33, 1213–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macgowan, A.P.; Bowker, K.E.; Wootton, M.; Holt, H.A. Activity of moxifloxacin, administered once a day, against Streptococcus pneumoniae in an in vitro pharmacodynamic model of infection. Antimicrob. Agents Chemother. 1999, 43, 1560–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vértesy, L.; Fehlhaber, H.W.; Kogler, H.; Schindler, P.W. Enkastines: Amadori Products with a Specific Inhibiting Action against Endopeptidase–24.11–from Streptomyces albus and by Synthesis. Liebigs Ann. 1996, 121–126. [Google Scholar] [CrossRef]
- Tsunakawa, M.; Kamei, H.; Konishi, M.; Miyaki, T.; Oki, T.; Kawaguchi, H. Porothramycin, a new antibiotic of the anthramycin group: Production, isolation, structure, and biological activity. J. Antibiot. 1988, 41, 1366–1373. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Hatori, M.; Tomita, K.; Sugawara, M.; Ikeda, C.; Nishiyama, Y.; Imanishi, H.; Miyaki, T.; Kawaguchi, H. Chicamycin, a new antitumor antibiotic. J. Antibiot. 1984, 37, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Osada, H.; Ishinabe, K.; Yano, T.; Kajikawa, K.; Isono, K. New Pyrrolobenzodiazepine Antibiotics, RK–1441A and B I. Biological Properties. Agric. Biol. Chem. 1990, 54, 2875–2881. [Google Scholar] [CrossRef]
- Huang, X.; He, J.; Niu, X.; Menzel, K.D.; Dahse, H.M.; Grabley, S.; Fiedler, H.P.; Sattler, I.; Hertweck, C. Benzopyrenomycin, a Cytotoxic Bacterial Polyketide Metabolite with a Benzo a pyrene-Type Carbocyclic Ring System. Angew. Chem. Int. Ed. 2008, 47, 3995–3998. [Google Scholar] [CrossRef]
- Pham, T.H.; Gardier, A.M. Fast–acting antidepressant activity of ketamine: Highlights on brain serotonin, glutamate, and GABA neurotransmission in preclinical studies. Pharmacol. Therapeut. 2019, 58–90. [Google Scholar] [CrossRef]
- Arai, T.; Uno, J.; Horimi, I.; Fukushima, K. Isolation of neocopiamycin A from Streptomyces hygroscopicus var. crystallogenes, the copiamycin source. J. Antibiot. 1984, 37, 103–109. [Google Scholar] [CrossRef]
- Seiga, K.; Yamaji, K. Microbiological study of copiamycin. Appl. Environ. Microbiol. 1971, 21, 986–989. [Google Scholar] [CrossRef] [Green Version]
- Butnariu, M.; Buțu, A. Functions of collateral metabolites produced by some actinomycetes. In Microbial Pathogens and Strategies for Combating them: Science, Technology, and Education; Formatex Research Center: Badajoz, Spain, 2013; pp. 1419–1425. [Google Scholar]
- Nisbet, L.J.; Mehta, R.J.; Oh, Y.; Pan, C.H.; Phelen, C.G.; Polansky, M.J.; Shearer, M.C.; Giovenella, A.J.; Grappel, S.F. Chlorocardicin, a monocyclic β–lactam from a Streptomyces sp. J. Antibiot. 1985, 38, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikushima, H.; Iguchi, E.; Kohsaka, M.; Aoki, H.; Imanaka, H. Streptomyces auranticolor sp. nov., a new anticoccidial antibiotics producer. J. Antibiot. 1980, 33, 1103–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Yuan, G.; Li, P.; Cao, S. Guanidine–containing polyhydroxyl macrolides: Chemistry, biology, and structure–activity relationship. Molecules 2019, 24, 3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, A.; Kubo, M.; Hirose, T.; Yahagi, K.; Tsunoda, N.; Noguchi, Y.; Nakashima, T.; Takahashi, Y.; Welz, C.; Mueller, D. Jietacins, azoxy antibiotics with potent nematocidal activity: Design, synthesis, and biological evaluation against parasitic nematodes. Eur. J. Med. Chem. 2018, 145, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.; Reddy, P.; Reddy, D.R. The effect of C2–fluoro group on the biological activity of DC–81 and its dimers. Bioorg. Med. Chem. Lett. 2004, 14, 2669–2672. [Google Scholar] [CrossRef] [PubMed]
- Russotti, G.; Göklen, K.E.; Wilson, J.J. Development of a pilot–scale microfiltration harvest for the isolation of physostigmine from Streptomyces griseofuscus broth. J. Chem. Technol. Biotechnol. 1995, 63, 37–47. [Google Scholar] [CrossRef]
- Mynderse, J.S.; O’Connor, S.C. Quiuolizine and indolizine enzyme inhibitors. U.S. Patent 4,508,901, 2 April 1985. [Google Scholar]
- Mayer, M.; Thiericke, R. Biosynthesis of streptazolin. J. Org. Chem. 1993, 58, 3486–3489. [Google Scholar] [CrossRef]
- Chaudhary, H.S.; Soni, B.; Shrivastava, A.R.; Shrivastava, S. Diversity and versatility of actinomycetes and its role in antibiotic production. J. Appl. Pharm. Sci. 2013, 3, S83–S94. [Google Scholar]
- Beck, J.; Guminski, Y.; Long, C.; Marcourt, L.; Derguini, F.; Plisson, F.; Grondin, A.; Vandenberghe, I.; Vispé, S.; Brel, V. Semisynthetic neoboutomellerone derivatives as ubiquitin–proteasome pathway inhibitors. Bioorg. Med. Chem. 2012, 20, 819–831. [Google Scholar] [CrossRef]
- Blum, S.; Groth, I.; Rohr, J.; Fiedler, H.P. Biosynthetic capacities of actinomycetes. 5. Dioxolides, novel secondary metabolites from Streptomyces tendae. J. Basic Microbiol. 1996, 36, 19–25. [Google Scholar] [CrossRef]
- Koyama, Y.; Yokose, K.; Dolby, L.J. Isolation, characterization, and synthesis of pimprinine, pimprinethine and pimprinaphine, metabolites of Streptoverticillium olivoreticuli. Agric. Biol. Chem. 1981, 45, 1285–1287. [Google Scholar] [CrossRef]
- Lee, H.-B.; Choi, Y.-C.; Kim, S.-U. Isolation and identification of maculosins from Streptomyces rochei 87051–3. Appl. Biol. Chem. 1994, 37, 339–342. [Google Scholar]
- Ortiz-López, F.; Alcalde, E.; Sarmiento-Vizcaíno, A.; Díaz, C.; Cautain, B.; García, L.; Blanco, G.; Reyes, F. New 3–Hydroxyquinaldic Acid Derivatives from Cultures of the Marine Derived Actinomycete Streptomyces cyaneofuscatus M–157. Mar. Drugs 2018, 16, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massoud, M.A.; el Bialy, S.A.; Bayoumi, W.A.; el Husseiny, W.M. Synthesis of new 2–and 3–hydroxyquinoline–4–carboxylic acid derivatives as potential antioxidants. Heterocycl. Commun. 2014, 20, 81–88. [Google Scholar] [CrossRef]
- Mori, M.; Uozumi, Y.; Kimura, M.; Ban, Y. Total syntheses of prothracarcin and tomaymycin by use of palladium catalyzed carbonylation. Tetrahedron 1986, 42, 3793–3806. [Google Scholar] [CrossRef]
- Shimizu, K.-i.; Kawamoto, I.; Tomita, F.; Morimoto, M.; Fujimoto, K. Prothracarcin, a novel antitumor antibiotic. J. Antibiot. 1982, 35, 972–978. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.C.; White, R.L.; Le, Y.; Vining, L.C. Isolation of N–acetyl–3, 4–dihydroxy–L–phenylalanine from Streptomyces akiyoshiensis. J. Nat. Prod. 1995, 58, 1274–1277. [Google Scholar] [CrossRef]
- Kannan, S. Screening for antiviral activity of Actinomycetes isolated from soil sediments. Ph.D. Thesis, Nandha College of Pharmacy, Erode, India, 2009. [Google Scholar]
- Hurley, L.H. Pyrrolo (1, 4) benzodiazepine antitumor antibiotics. Comparative aspects of anthramycin, tomaymycin and sibiromycin. J. Antibiot. 1977, 30, 349–370. [Google Scholar] [CrossRef]
- Deepika, L.; Kannabiran, K. Antagonistic activity of streptomyces vitddk1 spp.(gu223091) isolated from the coastal region of tamil nadu, India. Pharmacologyoline 2010, 1, 17–29. [Google Scholar]
- Harada, S. New Cephalosporins. J. Chromatogr. Libr. 1989, 43, 233–257. [Google Scholar]
- Lee, H.C.; Liou, K.; Kim, D.H.; Kang, S.-Y.; Woo, J.-S.; Sohng, J.K. Cystocin, a novel antibiotic, produced byStreptomyces sp. GCA0001: Biological activities. Arch. Pharm. Res. 2003, 26, 446–448. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Accession ID | Identity [%] | Source | Ref |
|---|---|---|---|---|
| Micromonospora terminaliae DSM 101760 | CP045309.1 | 96.678 | Surface sterilized stem of Thai medicinal plant Terminalia mucronata | [51] |
| Micromonospora inositola DSM 43819 | LT607754.1 | 96.263 | forest soil | [52] |
| Micromonospora cremea CR30 | NR_108478.1 | 96.258 | rhizosphere of Pisum sativum | [53] |
| Micromonospora rosaria DSM 803 | NR_026282.1 | 96.125 | unknown | [54] |
| Micromonospora palomenae NEAU-CX1 | NR_136848.1 | 96.055 | Nymphs of stinkbug (Palomena viridissima Poda) | [55] |
| Isolate | Accession ID | Identity [%] | Source | Ref |
|---|---|---|---|---|
| Gordonia hongkongensis HKU50 | NR_152023.1 | 95.386 | human blood culture | [56] |
| Gordonia terrae 3612 | CP016594.1 | 95.320 | soil | [57] |
| Gordonia bronchialis DSM 43247 | NR_074529.1 | 94.470 | human sputum | [58] |
| Gordonia desulfuricans 213E | NR_028734.1 | 94.412 | soil | [59] |
| Gordonia rubripertincta DSM 43248 | NR_043330.1 | 94.345 | soil | [60] |
| Isolate | Accession ID | Identity [%] | Source | Ref |
|---|---|---|---|---|
| Nocardia xestospongiae ST01-07 | NR_156866.1 | 96.972 | Xestospongia sp. | [61] |
| Nocardia amikacinitolerans NBRC 108937 | NR_117564.1 | 96.972 | human eye (clinical isolate) | [62] |
| Nocardia arthritidis DSM 44731 | NR_115824.1 | 96.898 | human sputum | [63] |
| Nocardia araoensis NBRC 100135 | NR_118199.1 | 96.677 | human | [64] |
| Nocardia beijingensis DSM 44636 | NR_118618.1 | 96.529 | mud from a sewage ditch | NA |
| Sample Code | MIC (µg/mL) | MIC (µg/mL, 72 h.) Trypanosoma brucei TC 221 | ||
|---|---|---|---|---|
| Staphylococcus aureus NCTC 8325 | Enterococcus faecalis | Candida albicans 5314 | ||
| Micromonospora sp. UA17 | 15.6 | 14.3 | 13.2 | >100 |
| Gordonia sp. UA19 | 35.7 | 31.9 | 16.8 | >100 |
| Nocardia sp. UA 23 | 38.9 | 39.2 | 25.7 | 7.2 * |
| UA17 + UA19 | 8.6 * | 7.4 * | 6.4 * | >100 |
| UA17 + UA23 | 4.2 * | 3.9 * | 3.8 * | >100 |
| UA17 + Myc | 4.7 * | 3.8 * | 5.9 * | >100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamikh, Y.I.; El Shamy, A.A.; Gaber, Y.; Abdelmohsen, U.R.; Madkour, H.A.; Horn, H.; Hassan, H.M.; Elmaidomy, A.H.; Alkhalifah, D.H.M.; Hozzein, W.N. Actinomycetes from the Red Sea Sponge Coscinoderma mathewsi: Isolation, Diversity, and Potential for Bioactive Compounds Discovery. Microorganisms 2020, 8, 783. https://doi.org/10.3390/microorganisms8050783
Shamikh YI, El Shamy AA, Gaber Y, Abdelmohsen UR, Madkour HA, Horn H, Hassan HM, Elmaidomy AH, Alkhalifah DHM, Hozzein WN. Actinomycetes from the Red Sea Sponge Coscinoderma mathewsi: Isolation, Diversity, and Potential for Bioactive Compounds Discovery. Microorganisms. 2020; 8(5):783. https://doi.org/10.3390/microorganisms8050783
Chicago/Turabian StyleShamikh, Yara I., Aliaa A. El Shamy, Yasser Gaber, Usama Ramadan Abdelmohsen, Hashem A. Madkour, Hannes Horn, Hossam M. Hassan, Abeer H. Elmaidomy, Dalal Hussien M. Alkhalifah, and Wael N. Hozzein. 2020. "Actinomycetes from the Red Sea Sponge Coscinoderma mathewsi: Isolation, Diversity, and Potential for Bioactive Compounds Discovery" Microorganisms 8, no. 5: 783. https://doi.org/10.3390/microorganisms8050783