1. Introduction
Fungi of the genus
Fusarium are known as plurivorous pathogens of various crops worldwide.
Fusarium diseases are caused by several co-occurring species which infect small-grain cereals like wheat, barley, oat, rye and triticale as well as maize. Many species attack a range of plants parts and stages, such as seedlings, roots, stems and heads, causing
Fusarium head blight (FHB) of small-grain cereals (also known as scab or ear blight) and ear rot of maize [
1,
2,
3,
4].
FHB of wheat (
Triticum aestivum L.) is one of the most important diseases in wheat-growing regions around the world causing quantitative and qualitative losses. Symptoms of FHB infections usually include premature bleaching of the entire head or just a few spikelets (
Figure 1a,b), pinkish-red mycelium and spores on infected spikelets (
Figure 1b), inhibited grain formation and grain that are shrivelled, light weighted and discoloured from white to pink as a result of the mycelial outgrowths from the
Fusarium colonized grain (
Figure 1c). If the rachis is infected, the head above the point of infection senesces prematurely due to the undersupply with water and nutrients with corresponding shrivelled, uninfected grain. Together this results in yield losses, reductions of the thousand grain weight (TGW) and germination ability of the harvested grain [
1,
2,
5,
6]. Furthermore, the baking quality is adversely affected by the reduction of starch content and degradation of different protein fractions [
7].
In addition, FHB causes quality losses of grain by the formation of mycotoxins, which causes a potential health risk for animals and humans, because their occurrence in feed and food is often associated with chronic or acute mycotoxicosis. Several
Fusarium species are capable to produce a range of mycotoxins including zearalenone (ZEA), moniliformin (MON), beauvericin (BEA), enniatins (ENs) and trichothecenes such as deoxynivalenol (DON), nivalenol (NIV), 3- and 15-acetyl-DON (3-AcDON, 15-AcDON), HT-2 and T-2 toxin. The often co-occurring mycotoxins DON and ZEA, mainly produced by the
Fusarium species
F. graminearum and
F. culmorum, are the most common mycotoxins associated with FHB in wheat. Exposure to DON induces chronic effects such as anorexia and reduced growth, as well as acute intoxication leading to vomiting and depressed immune function. ZEA has been proven to be hepatotoxic, immunotoxic and carcinogenic to a number of mammalian species and is able to bind to oestrogen receptors causing reproductive disorders (e.g., hyper-oestrogenism and infertility in livestock) [
2,
4,
8]. Within the EU, limit thresholds have already been defined for DON and ZEA in unprocessed grain and human food based on cereal grain. Limits for unprocessed wheat grain intended for use as food are 1250 µg DON/kg and 100 µg ZEA/kg [
9]. Likewise, in other countries of the world, maximum levels for DON and other mycotoxins were established, e.g., Canada, China, Russia and USA [
4].
FHB of wheat is caused by a complex of different toxigenic
Fusarium species. In Europe, the major causal agents of FHB are
F. graminearum,
F. culmorum,
F. avenaceum and
F. poae. However,
F. equiseti,
F. langsethiae,
F. sporotrichioides and
F. tricinctum are also frequently found [
1,
2,
10,
11,
12]. Regional and year-specific differences in the qualitative and quantitative species profile are particularly dependent on meteorological conditions and the cropping system (e.g., crop rotation, previous crop, tillage practices, cultivar susceptibility) [
10,
13]. Crop residues in the field are the major reservoir on which FHB pathogens saprophytically survive and generally produce asexual conidia, but some also produce sexual ascospores, e.g.,
F. graminearum (teleomorph
Gibberella zeae) and
F. avenaceum (teleomorph
Gibberella avenacea) [
2,
14]. Conidia are dispersed from crop residues by rain splashing, but wind dispersal is also important, especially for ascospores, which can travel meters to kilometres [
15,
16,
17]. In general, wheat heads are most susceptible to FHB infections at flowering, when the flower is directly exposed to the environment. Warm and moist conditions during this period encourage the development of the disease [
1,
18,
19]. Spores are deposited on or inside wheat florets where they germinate and initiate infection. The fungus infects extruded anthers and then ramifies throughout florets, the developing caryopsis and rachis spreading then to other flowers in the spikelet [
20]. Besides flowering, significant precipitation and high relative humidity during the periods of several days before (heading) and after flowering during grain filling increase the occurrence of FHB and mycotoxin contamination [
21,
22,
23,
24]. The composition and predominance of
Fusarium species in wheat is, to a large extent, determined by meteorological factors like moisture (precipitation, relative humidity) and temperature. Traditionally
F. culmorum and
F. avenaceum are associated with wet and cool maritime conditions,
F. poae with relatively drier and warmer conditions, while
F. graminearum dominates in regions with warm and humid environments [
18,
19]. Furthermore, FHB infections of wheat are determined by agronomic factors like crop rotation, previous crop, and soil cultivation. FHB epidemics are favoured when wheat follows host crops including small-grain cereals and especially silage maize and grain maize together with reduced cultivation tillage systems that preserve organic residues from the previous crop on the soil surface, suitable for saprophytic survival of the fungi and serving as a source of inoculum for
Fusarium spores [
14,
20,
25,
26,
27]. Different wheat cultivars are known to express different levels of susceptibility towards FHB and therefore, the choice of cultivar is currently the most effective agronomic method to reduce FHB infections and mycotoxin contamination [
27,
28,
29].
Due to differences in pathogenicity, toxigenicity, and fungicide sensitivity, it is important to obtain detailed knowledge of the qualitative and quantitative occurrence of individual
Fusarium species [
11]. The visual assessment of disease severities and incidences of affected heads, spikelets or damaged grain at harvest only estimate disease symptoms, but not pathogen populations. In contrast, mycological methods based on the stimulation of fungal growth from grain (or heads) infected by
Fusarium on agar plates and subsequent morphological analysis determine the spectrum and the incidence of the different FHB-causing species. The standard parameter is therefore the percentage of infected grain by the different FHB-causing fungi. These methods require taxonomical expertise, are time expensive and risk the overestimating of fast-growing species [
30]. Compared to the aforementioned methods, polymerase chain reaction (PCR)-based methods using species-specific primers have the advantage of species specificity, sensitivity and speed without the need for isolation and cultivation of the fungi from grain or heads. Furthermore, quantitative PCR (qPCR) offers the opportunity for quantifying DNA amounts and reveals the true dominance of the individual species in the FHB complex [
11,
31].
In the present study, harvested winter wheat grain of different susceptible cultivars from maize-free crop rotations was collected in a five-year survey from 2013 to 2017 at seven locations in northern Germany, a suitable growing area for wheat. Besides the determination of mycotoxin contamination, the Fusarium community in wheat grain was characterized by species-specific qPCR assays. The aims of the study were (i) to characterize the year-specific composition and quantitative predominance of species belonging to the genus Fusarium by quantifying DNA amounts as well as DON and ZEA concentrations, (ii) to analyse interactions between the occurring species, (iii) to determine differences in the qualitative and quantitative species complex and mycotoxin concentrations between different susceptible wheat cultivars, and (iv) to evaluate the effects of regional meteorological factors before, during and after flowering on DNA amounts of the different species in the Fusarium community and DON and ZEA concentrations in order to characterize the most crucial stages of wheat development towards FHB infections and mycotoxin contamination.
4. Discussion
The present study reports the results of a five-year survey from 2013 to 2017 on the composition and quantitative predominance of species belonging to the genus Fusarium by quantifying DNA amounts as well as DON and ZEA concentrations in harvested grain samples of winter wheat, depending on cultivar susceptibility and meteorological factors at seven locations in the northernmost federal state of Germany, Schleswig-Holstein. This state is a suitable growing area with large acreages of winter wheat and maritime weather conditions, which are conducive for FHB infections, being a representative region for these analyses.
In our study, it characterized the
Fusarium community in 315 grain samples. Using traditional culturing methods for this large number of samples is extremely laborious, requires taxonomical expertise, is time expensive, and increases the risk of overestimating fast-growing species. Culturing methods based on the stimulation of fungal growth from grain on agar plates and subsequent analysis of the different FHB species by morphological determination or qualitative PCR of mycelium outgrowing from the
Fusarium colonized grain only provide the incidence of the different FHB species by determination of the percentage of grain infected by the different
Fusarium species [
11,
30,
31]. Furthermore, the visual assessment of damaged grain at harvest or the quantitative detection of mycotoxins only estimate disease symptoms or mycotoxin contamination produced by several
Fusarium species, but not pathogen populations [
30]. Many studies analysing the regional and year-specific occurrence of FHB or the effects of meteorological and agronomic factors focus on parameters such as DON contamination [
22,
25,
26,
27,
28,
35,
45,
46,
47], FHB-damaged grain [
28,
46,
47], incidence and severity of FHB-affected heads and spikelets [
25,
26,
28,
45,
46,
47] or incidence of individual species in wheat grain or heads by culturing methods with the subsequent morphological [
12,
45,
46,
48,
49] or molecular (qualitative PCR) [
50,
51,
52] determination. The quantitative PCR (qPCR) method used in our study allowed the detection and quantification of species present in very low amounts in wheat grain, in contrast to traditional culturing methods, where the presence of especially slow-growing species will be underestimated. Furthermore, this method reveals the true dominance of the individual species in the FHB complex in wheat grain, which is of paramount importance, due to differences in pathogenicity, environmental requirements, toxin-production abilities, and fungicide sensitivity [
10,
11,
31].
Regarding FHB populations found in wheat grain, the situation was very similar to other European regions from a qualitative point of view [
1,
2,
10,
11,
12,
13,
31,
48,
49,
51,
52,
53,
54,
55,
56,
57,
58,
59,
60], with the FHB complex mainly composed of
F. graminearum,
F. culmorum,
F. avenaceum and
F. poae. In addition,
F. tricinctum and
F. langsethiae were detected, which were also found in other European surveys.
F. sporotrichioides and
F. equiseti were not detected, although a sporadic occurrence could be observed in wheat in some regions of Europe [
2,
11,
12,
54,
55,
56,
57]. Within the five-year survey
F. graminearum,
F. culmorum,
F. avenaceum and
F. poae were found in ≥ 80% of all samples through the entire period of study in the highly susceptible cultivar. High percentages of infected grain samples were also detected for
F. tricinctum and
F. langsethiae. In contrast to this qualitative assertion, DNA amounts of the abovementioned species differed considerably between years and especially between species within a year.
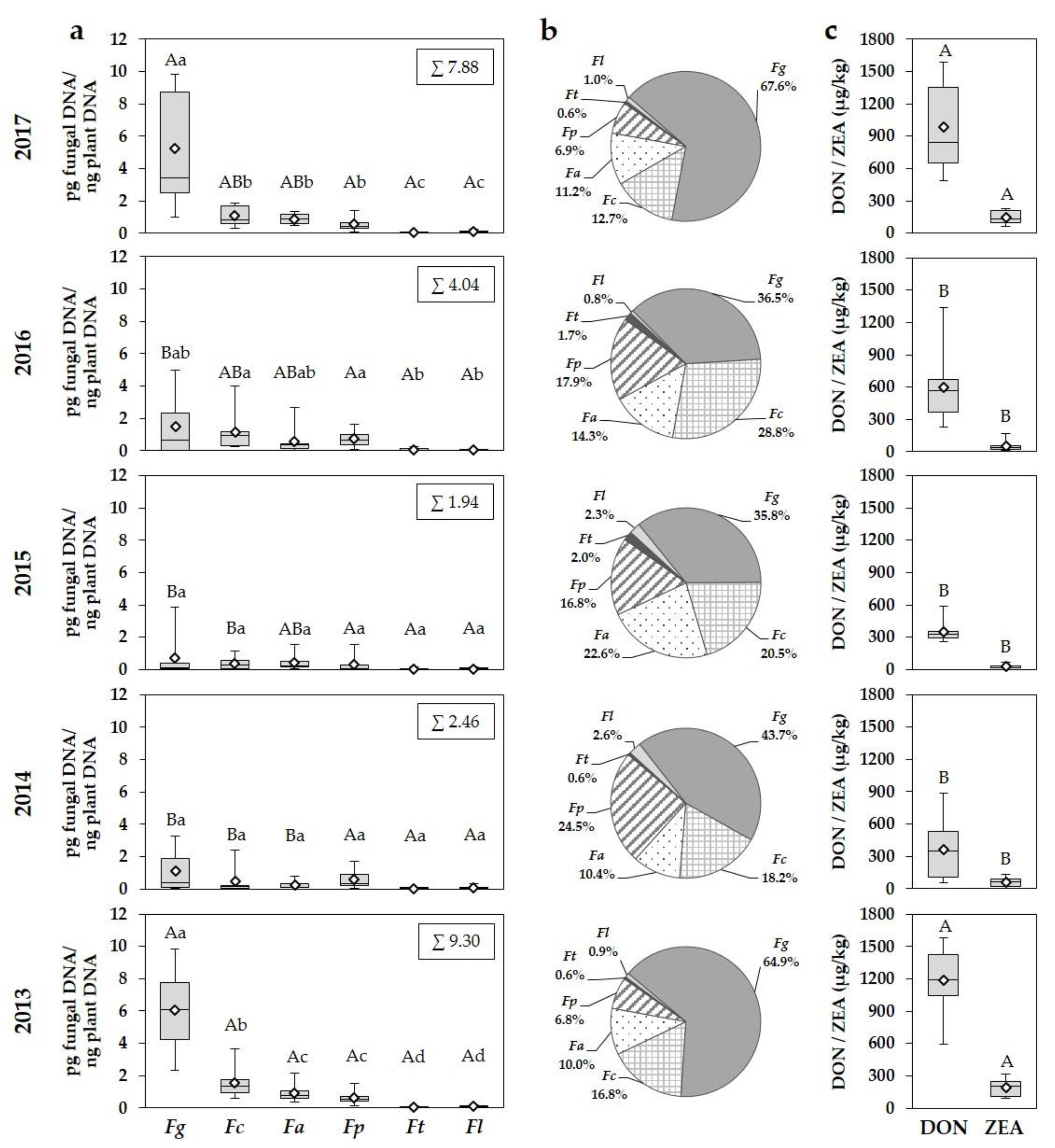
F. graminearum was the most dominant species in all years followed by
F. culmorum,
F. avenaceum and
F. poae, while
F. tricinctum and
F. langsethiae played only a very subordinate role in the FHB complex.
F. graminearum reached the highest percentages of the total
Fusarium DNA of all detected species, with maximum percentages of up to 70% in years favourable for the development of the FHB disease, averaging nearly 60% for the entire period of study.
Despite the fact that the wet and cool maritime conditions that are generally encountered in the area surveyed would seem to favour infections by
F. culmorum,
F. graminearum was generally the most prevalent species in wheat grain samples in terms of DNA amounts over the entire period of study. Traditionally,
F. culmorum is associated with cool, wet and humid conditions, while
F. graminearum dominates in regions with warm and humid environments [
18,
19]. Not only in Germany, but also in many other countries of Europe,
F. graminearum becomes the major FHB constituent expanding towards regions with cooler and maritime conditions, replacing
F. culmorum as the main causal agent of FHB in the past [
10,
11,
49,
53,
54,
56,
58,
59]. The emergence of
F. graminearum has been often linked to an increased production of maize, another primary host of
F. graminearum in contrast to
F. culmorum [
10,
11,
13,
48]. Maize crop residues promote the mass production of ascospores of
F. graminearum, which appear to greatly outnumber conidia and which can travel over long distances [
14,
15,
16,
17,
61]. In contrast,
F. culmorum only produce asexual conidia, which are mainly dispersed by rain splashing [
2,
14]. Additionally, in the area surveyed a dramatic increase in maize (for bioenergy) was observed by 147%, from 79,200 ha in 2000 [
62] to 195,600 ha in 2011, with an unchanged cultivation area of wheat with 210,300 ha (arable crop area in 2011: 673,300 ha) [
63]. Although wheat grain samples within our survey originated from maize-free crop rotations,
F. graminearum was the predominant species in all years, indicating that the increased production area of maize is likely to be responsible for an increased infection potential of wheat with
F. graminearum in this area. Furthermore, the shift toward more
F. graminearum may involve an adaption to cooler climate, changes in climate toward warmer and more humid springs/summers and a superior competitiveness over other
Fusarium species [
10]. Although
F. graminearum dominated over
F. culmorum, a positive correlation of DNA amounts was found between these two species, indicating that they were favoured by similar meteorological conditions. This is in contrast to other reports [
49,
51], which found no or a negative interaction between
F. graminearum and
F. culmorum.
In addition to
F. graminearum and
F. culmorum,
F. avenaceum and
F. poae are often represented in the
Fusarium complex as causative agents of FHB in Europe, reaching partially high frequencies of infected grain or DNA amounts in wheat grain [
2,
10,
11,
12,
31,
48,
49,
52,
54,
56,
57,
58]. Like
F. culmorum,
F. avenaceum is associated with cool, wet and humid conditions, and is therefore often found as one of the predominant species in the FHB complex, mainly in northern Europe [
11,
19,
31,
57,
58].
F. poae has been described to be associated with warmer and drier conditions with an increasing occurrence in some years and regions of Europe [
19,
51,
54,
56]. Both species were not found in greater extent in our five-year survey and DNA amounts were apparently stable over the entire period of study and significantly lower than those of
F. graminearum. Fernandez and Chen [
64] and Xu et al. [
10] suggest that
F. graminearum is probably the most pathogenic FHB species, which rapidly infects wheat heads and developing grain under favourable meteorological conditions, which may preclude the infection by other FHB species. Xu et al. [
65,
66] described
F. poae and
F. avenaceum as less pathogenic than the more aggressive species
F. culmorum and especially
F. graminearum. They found that in mixed inoculations,
F. graminearum was most competitive, whereas
F. poae was the least competitive of the four species including
F. avenaceum and
F. culmorum, which was more competitive than
F. avenaceum. These findings suggest that
F. avenaceum and
F. poae can colonise wheat grain under meteorological conditions less favourable to
F. graminearum. The dominant and increased occurrence of
F. graminearum towards
F. avenaceum and
F. poae in the area surveyed was particularly observed in 2013 and 2017 under meteorological conditions favourable for
F. graminearum, while the remaining years 2014 to 2016 were, in general, not conductive for FHB infections.
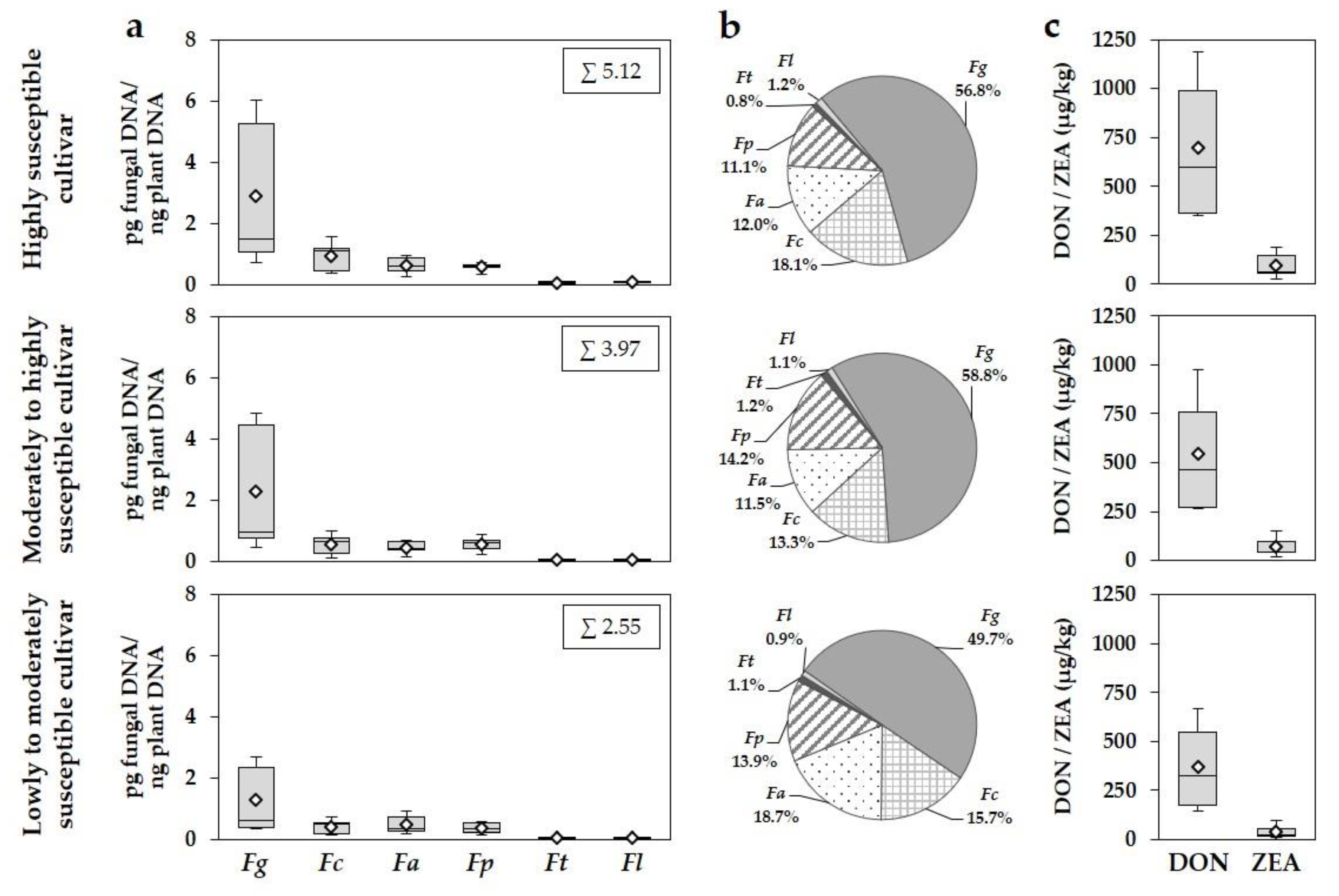
Our results show a differential susceptibility response of the three investigated cultivars to FHB infections and DON and ZEA concentrations. Most studies analysing the susceptibility against FHB focus on parameters such as DON content, FHB damaged grain, disease severity of affected heads and spikelets and frequency of individual
Fusarium species [
27,
28,
49,
50,
51,
67,
68], and were often conducted using artificial infections [
28,
67]. Beyer et al. [
27] noted that comparisons of different cultivars grown at different locations, in different years and after different pre-crops would bias a fair assessment of cultivar susceptibility. In our study, cultivar susceptibility was evaluated by measuring both DNA amounts of the co-occurring FHB species as well as DON and ZEA concentrations in wheat grain using natural field infections. Each of the three differently susceptible cultivars (highly, moderately to highly and lowly to moderately susceptible) was grown at each of the seven locations and in all years, which allowed a fair assessment between cultivars. Furthermore, soil cultivation and previous crop were the same at a location in each year and should have had no effect on FHB occurrence and mycotoxin contamination, which is in line with other authors [
14,
27,
50]. The FHB favourable reduced cultivation tillage was only applied after oilseed rape (non-host for FHB), ploughing was done after pre-crop wheat, burying the wheat residues for interrupting the saprophytic survival of the fungi on wheat stubbles. The ranking of the three cultivars in the five-year survey was similar between years and within a year, with the highly susceptible cultivar as the most susceptible and the lowly to moderately susceptible cultivar as the most tolerant cultivar. The same FHB species complex was found in all cultivars with a comparable year-specific quantitative occurrence of the six detected
Fusarium species as well as DON and ZEA concentrations, but with decreased DNA amounts and mycotoxin contents in the more tolerant cultivars. This was particularly pronounced in years with an increased disease pressure (2013, 2017). In all cultivars,
F. graminearum was the predominant species, followed by
F. culmorum,
F. avenaceum and
F. poae. The reduction in DNA amounts and mycotoxin concentrations between the three cultivars were mainly caused by the reduced DNA amounts of
F. culmorum and especially
F. graminearum in the more tolerant cultivars. Astonishingly, comparable percentages of DNA amounts of the six species to the total
Fusarium DNA amount of all detected species were found between the three cultivars for each species in all years. Increased cultivar resistance to FHB seems to limit the occurrence of almost all species of the FHB complex. Furthermore, no prevalence of a certain species to any of the tested cultivars was found. Mesterházy et al. [
28] also concluded that the degrees of FHB severity of wheat cultivars to different
Fusarium species, including
F. graminearum,
F. culmorum,
F. avenaceum and
F. poae, were very similar, indicating that the cultivar resistance to any
Fusarium species implies a similar level of resistance to other
Fusarium species.
Field surveys conducted elsewhere have indicated that the prevalence and severity of FHB varied strongly from year to year and also from location to location [
12,
48,
49,
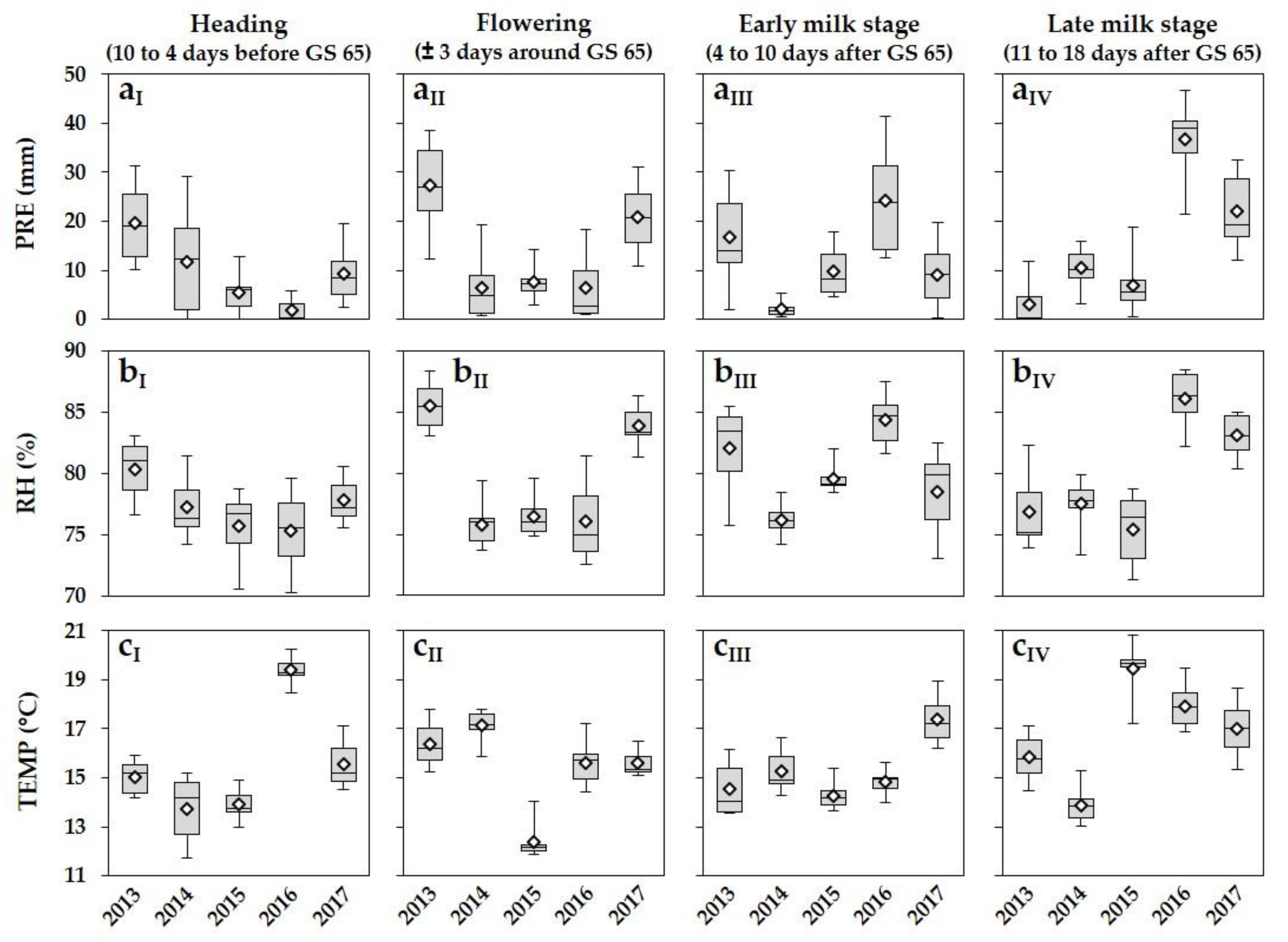
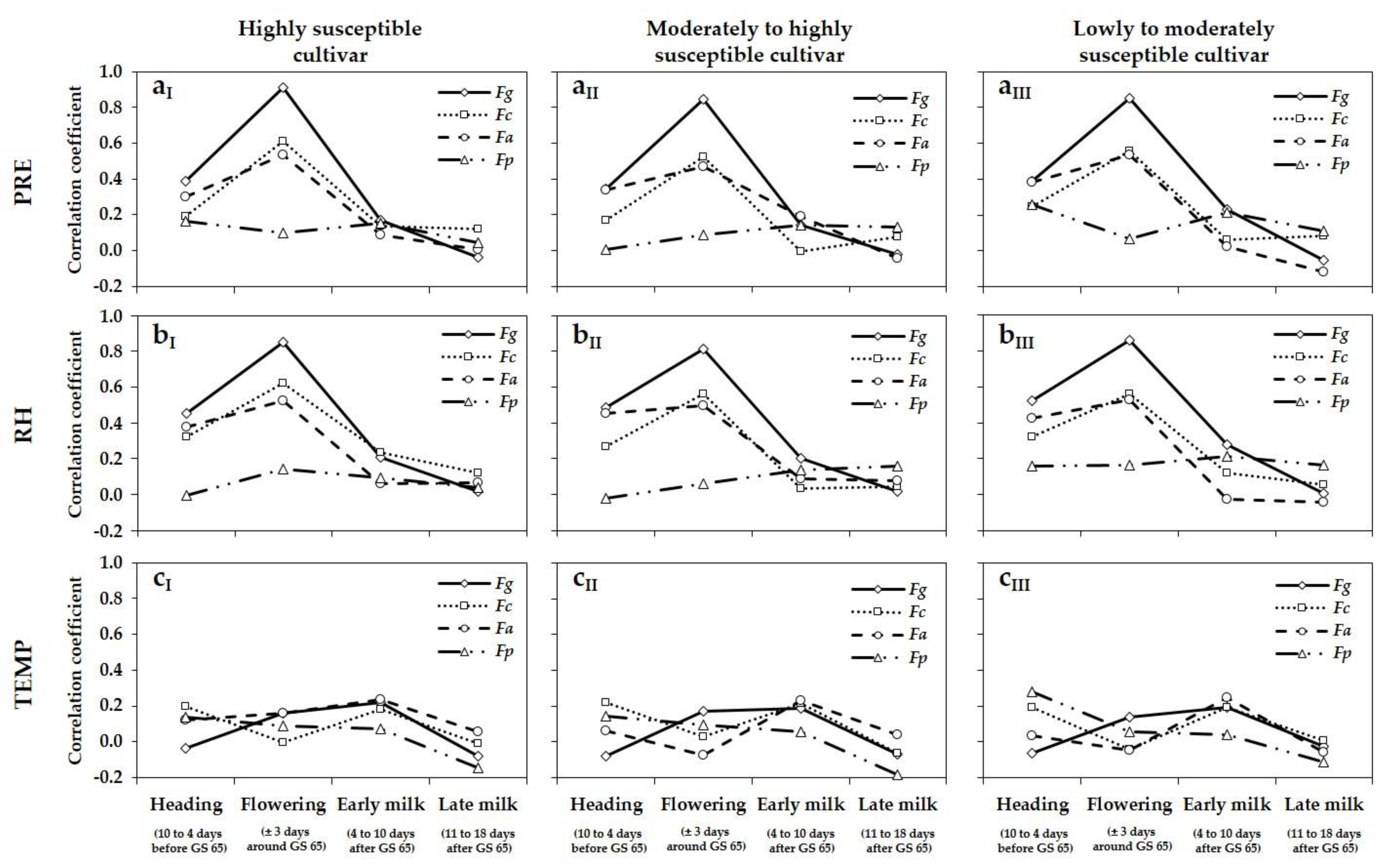
52]. In our five-year survey, DNA amounts as well as DON and ZEA concentrations also varied strongly from year to year and from location to location within a year. One of the major environmental factors influencing FHB occurrence are meteorological conditions, which vary often between growing seasons and locations within a season. Significant precipitation, high relative humidity and warm temperatures, coinciding with flowering of wheat, favour infection and development of FHB [
1,
18,
19]. Within our five-year survey, differences in DNA amounts, especially of
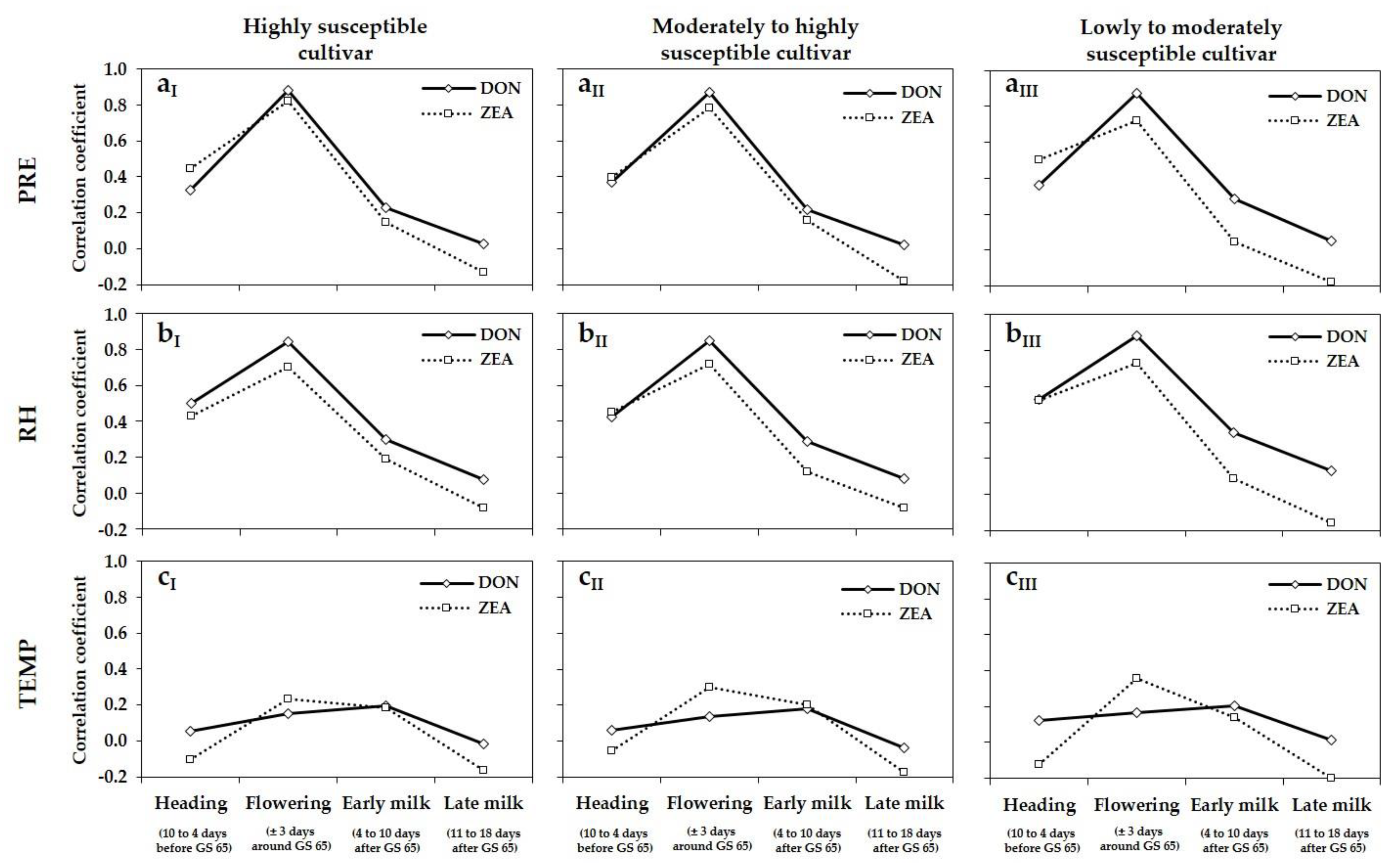
F. graminearum, and also DON and ZEA concentrations between growing seasons, were affected by the varying meteorological variables PRE and RH during flowering, shown by the highly positive correlations between these meteorological variables and DNA amounts of
F. graminearum and DON and ZEA concentrations. The highest DNA amounts of the predominant species
F. graminearum as well as mycotoxin concentrations were detected in 2013 and 2017 in the three different susceptible cultivars. These years were characterized by the highest values of PRE and RH at flowering. In 2014, 2015 and 2016, lower DNA amounts and mycotoxin concentrations were analysed, attributable to the lower values of PRE and RH during this period. The fluctuations of PRE and RH between locations within a year resulted in a large variation of DNA amounts and DON and ZEA concentrations within a year. Positive correlations were also observed between the meteorological variables PRE and RH, and DNA amounts of
F. culmorum and
F. avenaceum. Lacey et al. [
45] also showed that after artificial field inoculation with conidia of
F. culmorum at different growth stages of wheat, most head infection and DON contamination was restricted to the period of flowering, whereas little or no symptoms as well as lower DON concentrations resulted from inoculations before (head emergence) and after flowering (early and late milk stage). Similar results were obtained by Rossi et al. [
30], who inoculated wheat heads with conidia of
F. graminearum and
F. culmorum under field conditions at six growth stages between heading and dough ripening. Inoculations at flowering resulted in the highest incidences of infected grain and DON concentrations compared to inoculations before and after flowering. In contrast, Del Ponte et al. [
46] and Yoshida and Nakajima [
47] showed in their studies with artificial inoculations at different growth stages in the greenhouse that heavy FHB infections associated with high mycotoxin concentrations are still possible during the milk stage and often resulted in higher values of FHB damaged grain and mycotoxin concentrations compared to inoculations during flowering. However, our results indicate that under natural field infections, the periods several days before (heading) and after flowering (early and late milk stage) were less important towards FHB infections than during flowering. For the pre- and post-flowering periods the relationships between the meteorological variables PRE and RH and DNA amounts of
F. graminearum,
F. culmorum and
F. avenaceum and mycotoxin concentrations were less correlated than for the period of wheat flowering. There was no evidence of correlations between TEMP and DNA amounts of any
Fusarium species and DON and ZEA concentrations during flowering. For wheat, wetness periods of at least 24 h and temperatures above 15 °C are required for significant infection by
F. graminearum,
F. culmorum,
F. avenaceum and
F. poae [
18]. In almost all years, TEMP during flowering was higher than this critical value and does not seem to be a limiting factor, although higher optimal temperature requirements were described for several
Fusarium species [
18,
69], which will never be reached under field conditions in the area surveyed.
The often co-occurring mycotoxins DON and ZEA are the most common mycotoxins in wheat, associated with
F. graminearum and
F. culmorum [
2,
4,
8]. In our five-year survey, high levels of DON and ZEA were detected in wheat grain samples in years with a strong occurrence of these two species, especially with
F. graminearum. Correlation results between DNA amounts and DON and ZEA concentrations indicate that
F. graminearum is the main producer of DON and ZEA in wheat grain samples. Grain samples were not analysed for DON derivates 3-AcDON, 15-AcDON and NIV, which can be produced by the two aforementioned species [
2,
4]. In addition to
F. graminearum and
F. culmorum, other toxigenic species present in grain samples are capable to produce several mycotoxins.
F. avenaceum is known to produce MON, BEA and ENS,
F. poae to produce NIV and BEA.
F. langsethiae has been described as a producer of the highly toxic HT-2 and T-2 toxins, which can also be synthesized by a few isolates of
F. poae.
F. tricinctum is able to produce MON and ENS [
2,
4,
8,
54,
70]. We did not measure the concentrations of MON, BEA, ENS, NIV, HT-2 and T-2 toxin, but the range of
Fusarium species present in grain samples indicates a potential for the contamination with these mycotoxins as shown in others studies [
54,
56]. It must be noticed that
F. avenaceum and
F. poae often, and
F. tricinctum and
F. langsethiae in general occurred with very small DNA amounts in wheat grain, indicating that these species only produce small or very small amounts of the abovementioned mycotoxins. Due to the strong occurrence of
F. graminearum in wheat growing areas, a contamination with DON and ZEA in wheat grain can be expected.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}