Cytomegalovirus Strain TB40/E Restrictions and Adaptations to Growth in ARPE-19 Epithelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
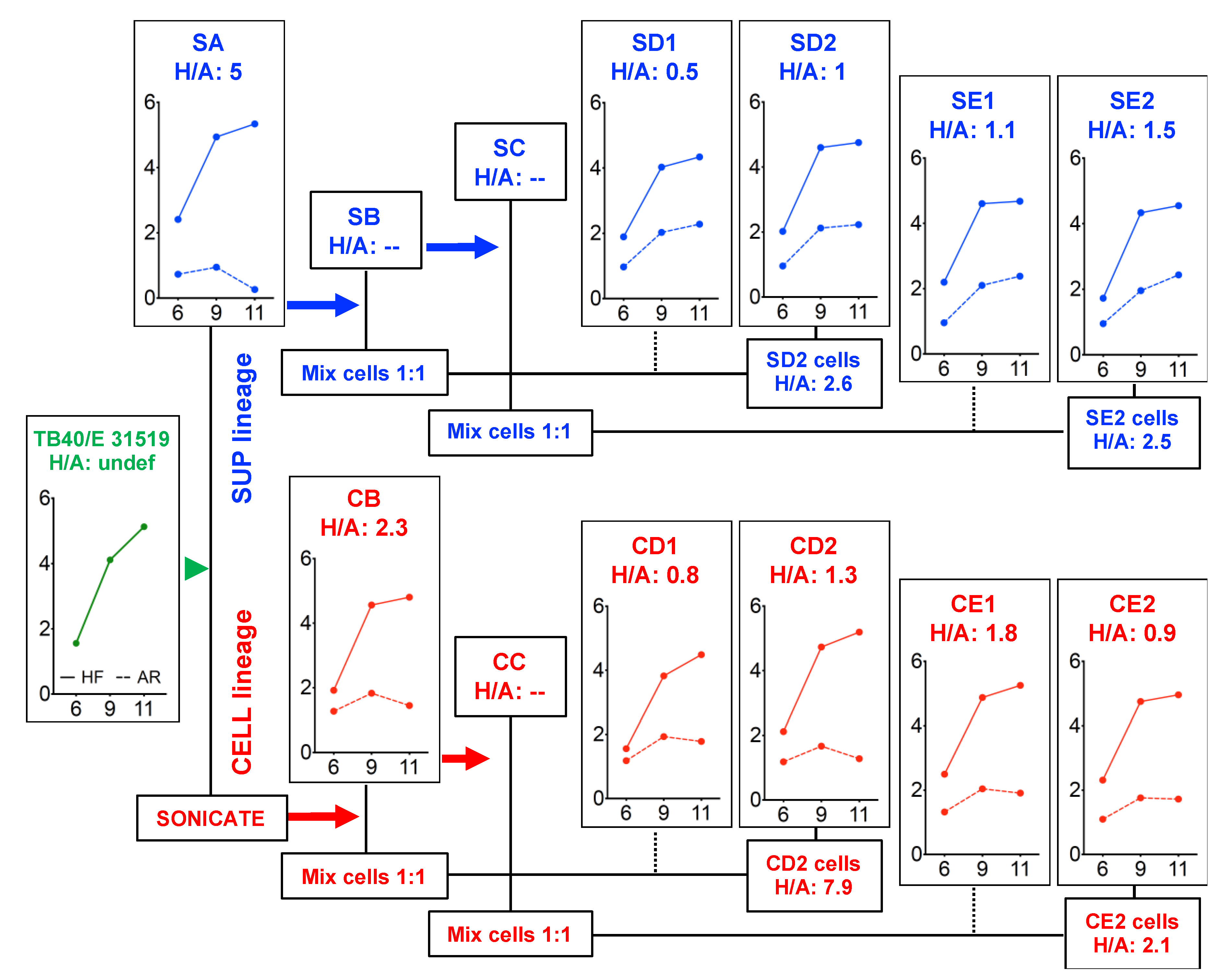
2.2. TB40/E Stock Adaptation in ARPE-19 Cells
2.3. Immunofluorescence Staining Analyses (IFA)
2.4. Virus Titrations
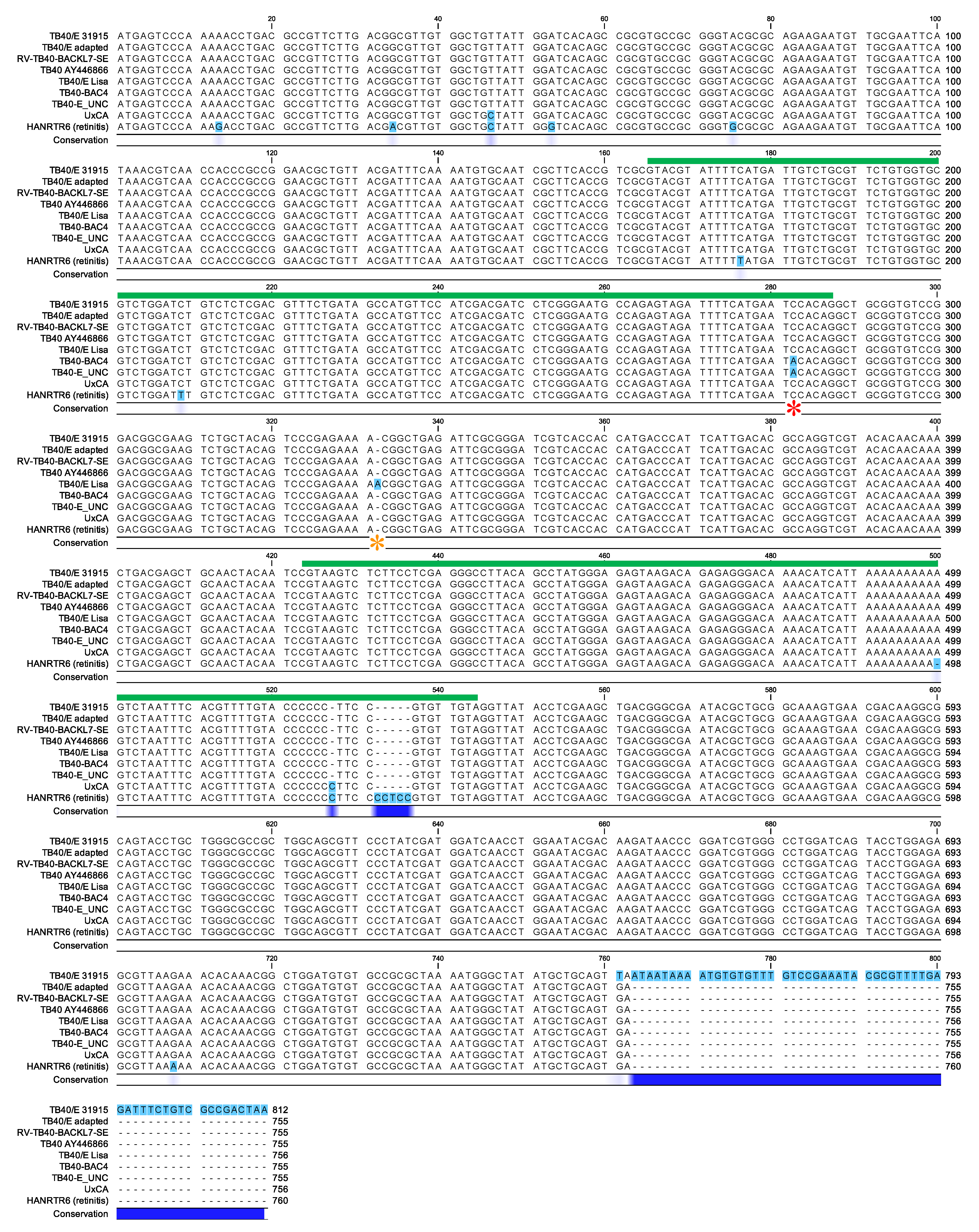
2.5. Polymerase Chain Reaction (PCR) Amplifications, Sequencing and Sequence Alignments
2.6. Real-Time Quantitative Genomic PCR
2.7. Statistical Analysis
3. Results
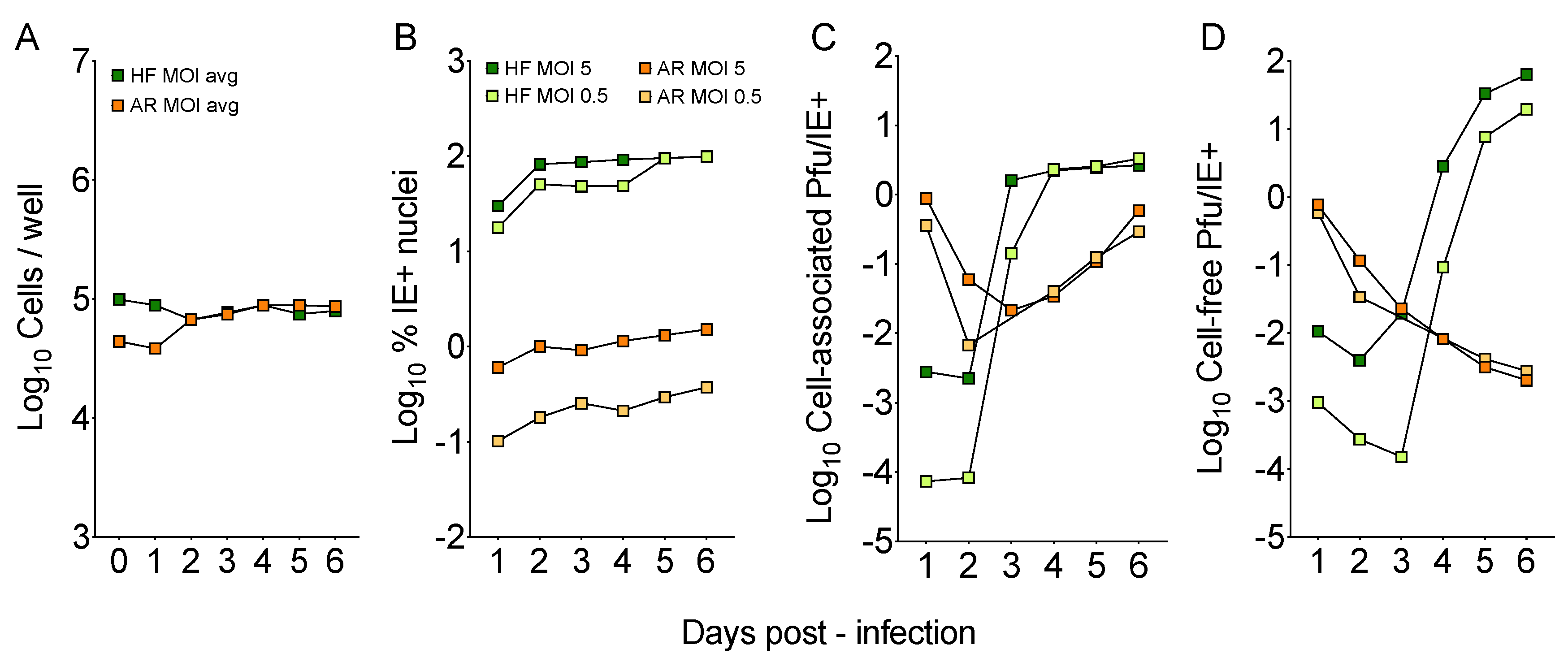
3.1. TB40/E Grown of Human Foreskin Fibroblasts (HFF) Shows Poor Tropism for ARPE-19 Epithelial Cells
3.2. The Ability to Initiate Infection in ARPE-19 Cells is Rapidly Gained upon Passaging on Epithelial Cells
3.3. Efficient Entry into ARPE-19 Cells is Associated with the Selection of TB40/E Variants Carrying a TGA (STOP) Codon at the 3′ End of UL128
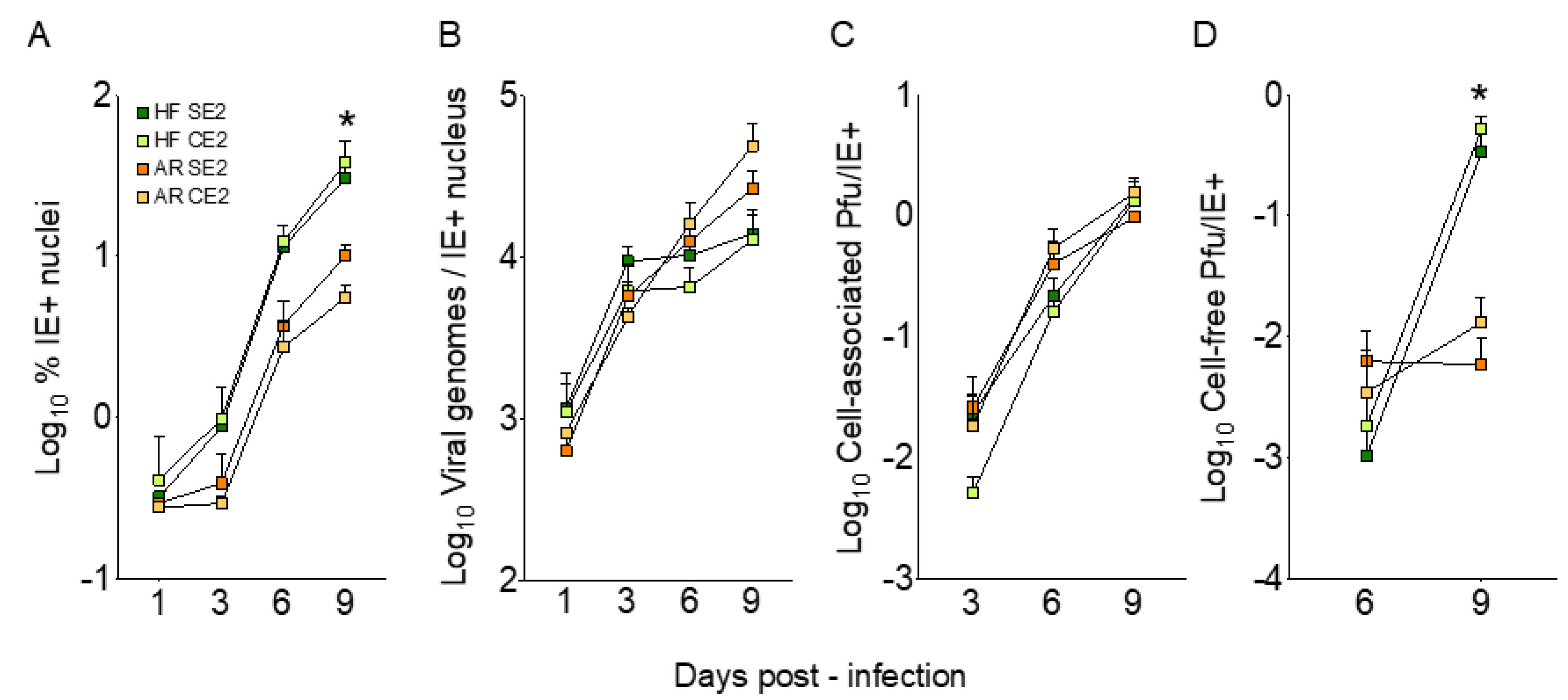
3.4. Cell-Free Progeny Release from ARPE-19 Cells Infected with Adapted Viruses Remains Hampered
3.5. Viral Genome Replication and Cell-Associated Progeny Production are Similarly Efficient in ARPE-19 Cells and HFF Infected with Adapted Viruses
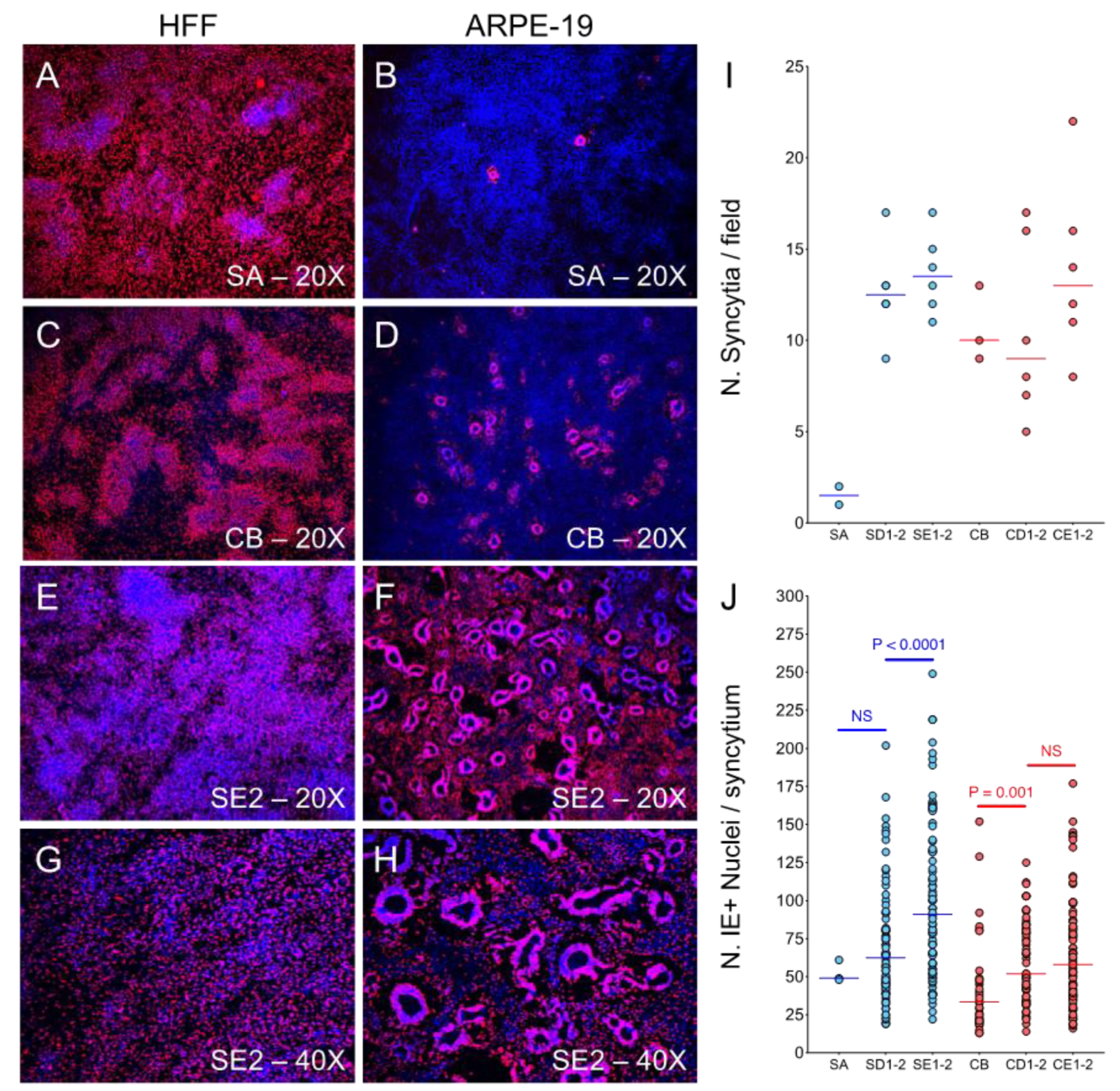
3.6. ARPE-19 Cultures Infected with Adapted Viruses Contain Numerous Large Syncytia at Late Times Post-Infection
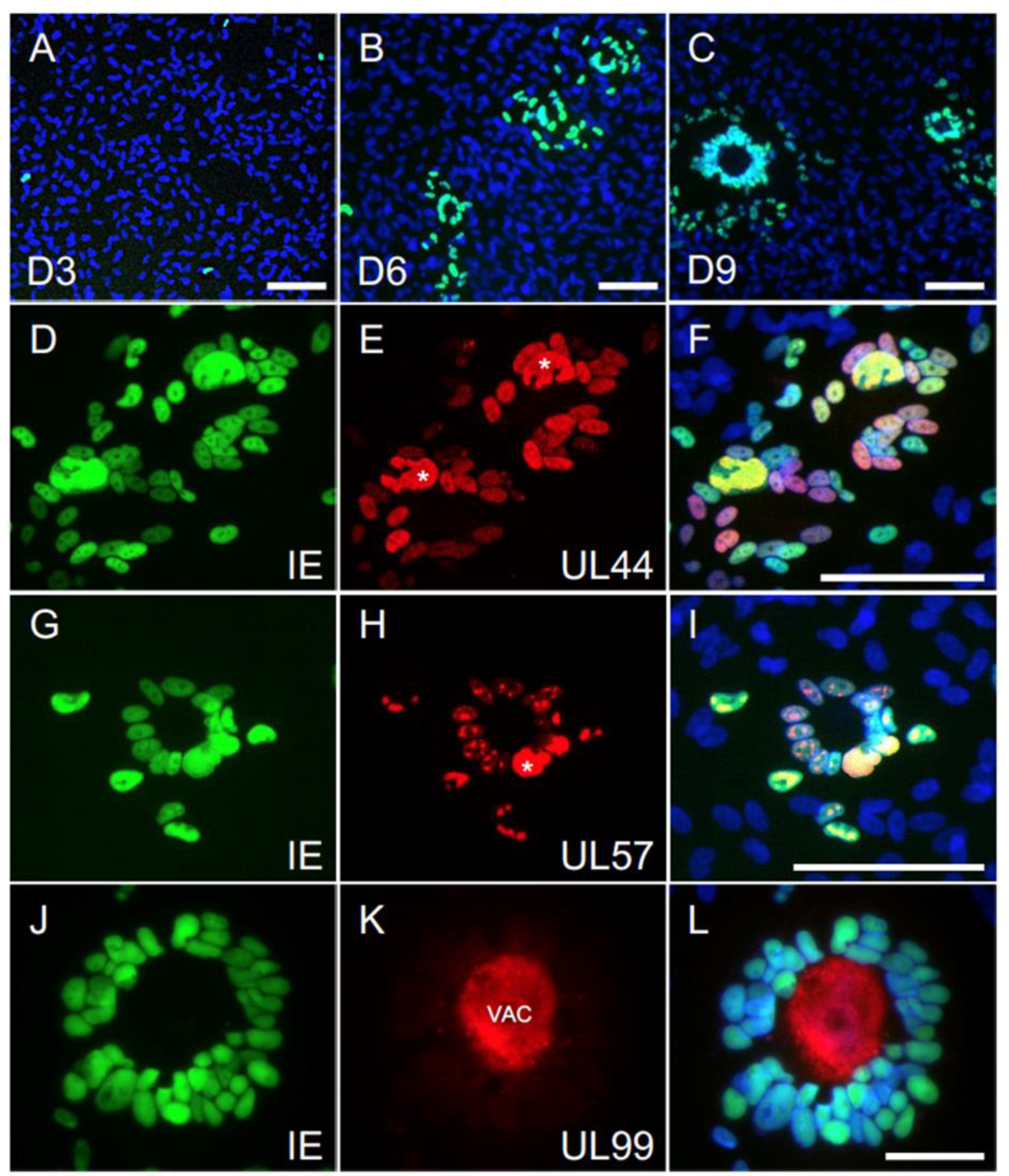
3.7. The Majority of Syncytia Contain Multiple IE+, UL44+ and UL57+ Nuclei but a Single UL99+ VAC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sinzger, C.; Digel, M.; Jahn, G. Cytomegalovirus cell tropism. Curr. Top. Microbiol. Immunol. 2008, 325, 63–83. [Google Scholar] [PubMed]
- Cui, X.; Freed, D.C.; Wang, D.; Qiu, P.; Li, F.; Fu, T.M.; Kauvar, L.M.; McVoy, M.A. Impact of antibodies and strain polymorphisms on cytomegalovirus entry and spread in fibroblasts and epithelial cells. J. Virol. 2017, 91, e01650-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerna, G.; Percivalle, E.; Sarasini, A.; Revello, M.G. Human cytomegalovirus and human umbilical vein endothelial cells: Restriction of primary isolation to blood samples and susceptibilities of clinical isolates from other sources to adaptation. J. Clin. Microbiol. 2002, 40, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prichard, M.N.; Penfold, M.E.; Duke, G.M.; Spaete, R.R.; Kemble, G.W. A review of genetic differences between limited and extensively passaged human cytomegalovirus strains. Rev. Med. Virol. 2001, 11, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Cha, T.A.; Tom, E.; Kemble, G.W.; Duke, G.M.; Mocarski, E.S.; Spaete, R.R. Human cytomegalovirus clinical isolates carry at least 19 genes not found in laboratory strains. J. Virol. 1996, 70, 78–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldman, W.J.; Sneddon, J.M.; Stephens, R.E.; Roberts, W.H. Enhanced endothelial cytopathogenicity induced by a cytomegalovirus strain propagated in endothelial cells. J. Med. Virol. 1989, 28, 223–230. [Google Scholar] [CrossRef]
- Waldman, W.J.; Roberts, W.H.; Davis, D.H.; Williams, M.V.; Sedmak, D.D.; Stephens, R.E. Preservation of natural endothelial cytopathogenicity of cytomegalovirus by propagation in endothelial cells. Arch. Virol. 1991, 117, 143–164. [Google Scholar] [CrossRef] [Green Version]
- Sinzger, C.; Schmidt, K.; Knapp, J.; Kahl, M.; Beck, R.; Waldman, J.; Hebart, H.; Einsele, H.; Jahn, G. Modification of human cytomegalovirus tropism through propagation in vitro is associated with changes in the viral genome. J. Gen. Virol. 1999, 80, 2867–2877. [Google Scholar] [CrossRef]
- Dolan, A.; Cunningham, C.; Hector, R.D.; Hassan-Walker, A.F.; Lee, L.; Addison, C.; Dargan, D.J.; McGeoch, D.J.; Gatherer, D.; Emery, V.C.; et al. Genetic content of wild-type human cytomegalovirus. J. Gen. Virol. 2004, 85, 1301–1312. [Google Scholar] [CrossRef]
- Tomasec, P.; Wang, E.C.; Davison, A.J.; Vojtesek, B.; Armstrong, M.; Griffin, C.; McSharry, B.P.; Morris, R.J.; Llewellyn-Lacey, S.; Rickards, C.; et al. Downregulation of natural killer cell-activating ligand CD155 by human cytomegalovirus UL141. Nat. Immunol. 2005, 6, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Sinzger, C.; Hahn, G.; Digel, M.; Katona, R.; Sampaio, K.L.; Messerle, M.; Hengel, H.; Koszinowski, U.; Brune, W.; Adler, B. Cloning and sequencing of a highly productive, endotheliotropic virus strain derived from human cytomegalovirus TB40/E. J. Gen. Virol. 2008, 89, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, K.L.; Weyell, A.; Subramanian, N.; Wu, Z.; Sinzger, C. A TB40/E-derived human cytomegalovirus genome with an intact US-gene region and a self-excisable BAC cassette for immunological research. Biotechniques 2017, 63, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wille, P.T.; Knoche, A.J.; Nelson, J.A.; Jarvis, M.A.; Johnson, D.C. A human cytomegalovirus gO-null mutant fails to incorporate gH/gL into the virion envelope and is unable to enter fibroblasts and epithelial and endothelial cells. J. Virol. 2010, 84, 2585–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryckman, B.J.; Jarvis, M.A.; Drummond, D.D.; Nelson, J.A.; Johnson, D.C. Human cytomegalovirus entry into epithelial and endothelial cells depends on genes UL128 to UL150 and occurs by endocytosis and low-pH fusion. J. Virol. 2006, 80, 710–722. [Google Scholar] [CrossRef] [Green Version]
- Ryckman, B.J.; Rainish, B.L.; Chase, M.C.; Borton, J.A.; Nelson, J.A.; Jarvis, M.A.; Johnson, D.C. Characterization of the human cytomegalovirus gH/gL/UL1281–31 complex that mediates entry into epithelial and endothelial cells. J. Virol. 2008, 82, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Ryckman, B.J.; Chase, M.C.; Johnson, D.C. HCMV gH/gL/UL1281–31 interferes with virus entry into epithelial cells: Evidence for cell type-specific receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 14118–14123. [Google Scholar] [CrossRef] [Green Version]
- Adler, B.; Scrivano, L.; Ruzcics, Z.; Rupp, B.; Sinzger, C.; Koszinowski, U. Role of human cytomegalovirus UL131A in cell type-specific virus entry and release. J. Gen. Virol. 2006, 87, 2451–2460. [Google Scholar] [CrossRef]
- Schuessler, A.; Sampaio, K.L.; Scrivano, L.; Sinzger, C. Mutational mapping of UL130 of human cytomegalovirus defines peptide motifs within the C-terminal third as essential for endothelial cell infection. J. Virol. 2010, 84, 9019–9026. [Google Scholar] [CrossRef] [Green Version]
- Schuessler, A.; Sampaio, K.L.; Sinzger, C. Charge cluster-to-alanine scanning of UL128 for fine tuning of the endothelial cell tropism of human cytomegalovirus. J. Virol. 2008, 82, 11239–11246. [Google Scholar] [CrossRef] [Green Version]
- Schuessler, A.; Sampaio, K.L.; Straschewski, S.; Sinzger, C. Mutational mapping of pUL131A of human cytomegalovirus emphasizes its central role for endothelial cell tropism. J. Virol. 2012, 86, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shenk, T. Human cytomegalovirus virion protein complex required for epithelial and endothelial cell tropism. Proc. Natl. Acad. Sci. USA 2005, 102, 18153–18158. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yu, Q.C.; Schroer, J.; Murphy, E.; Shenk, T. Human cytomegalovirus uses two distinct pathways to enter retinal pigmented epithelial cells. Proc. Natl. Acad. Sci. USA 2007, 104, 20037–20042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, G.; Revello, M.G.; Patrone, M.; Percivalle, E.; Campanini, G.; Sarasini, A.; Wagner, M.; Gallina, A.; Milanesi, G.; Koszinowski, U.; et al. Human cytomegalovirus UL1311–28 genes are indispensable for virus growth in endothelial cells and virus transfer to leukocytes. J. Virol. 2004, 78, 10023–10033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerna, G.; Percivalle, E.; Lilleri, D.; Lozza, L.; Fornara, C.; Hahn, G.; Baldanti, F.; Revello, M.G. Dendritic-cell infection by human cytomegalovirus is restricted to strains carrying functional UL1311–28 genes and mediates efficient viral antigen presentation to CD8+ T cells. J. Gen. Virol. 2005, 86, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Straschewski, S.; Patrone, M.; Walther, P.; Gallina, A.; Mertens, T.; Frascaroli, G. Protein pUL128 of human cytomegalovirus is necessary for monocyte infection and blocking of migration. J. Virol. 2011, 85, 5150–5158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauron, E.J.; Yu, D.; Fehr, A.R.; Hertel, L. Human cytomegalovirus infection of langerhans-type dendritic cells does not require the presence of the gH/gL/UL1281–31A complex and is blocked after nuclear deposition of viral genomes in immature cells. J. Virol. 2014, 88, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronel, R.; Takayama, S.; Juwono, T.; Hertel, L. Dynamics of human cytomegalovirus infection in CD34+ hematopoietic cells and derived langerhans-type dendritic cells. J. Virol. 2015, 89, 5615–5632. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Yu, Q.; Wechsler, A.; Ryckman, B.J. Comparative analysis of gO isoforms reveals that strains of human cytomegalovirus differ in the ratio of gH/gL/gO and gH/gL/UL1281–31 in the virion envelope. J. Virol. 2013, 87, 9680–9690. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, C.M.; Murphy, E.A. A myeloid progenitor cell line capable of supporting human cytomegalovirus latency and reactivation, resulting in infectious progeny. J. Virol. 2012, 86, 9854–9865. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Oberstein, A.; Wang, W.; Shenk, T. Role of PDGF receptor-alpha during human cytomegalovirus entry into fibroblasts. Proc. Natl. Acad. Sci. USA 2018, 115, E9889–E9898. [Google Scholar] [CrossRef] [Green Version]
- Murrell, I.; Tomasec, P.; Wilkie, G.S.; Dargan, D.J.; Davison, A.J.; Stanton, R.J. Impact of sequence variation in the UL128 locus on production of human cytomegalovirus in fibroblast and epithelial cells. J. Virol. 2013, 87, 10489–10500. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Nguyen, C.C.; Ryckman, B.J.; Britt, W.J.; Kamil, J.P. A viral regulator of glycoprotein complexes contributes to human cytomegalovirus cell tropism. Proc. Natl. Acad. Sci. USA 2015, 112, 4471–4476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.C.; Siddiquey, M.N.A.; Zhang, H.; Li, G.; Kamil, J.P. Human cytomegalovirus tropism modulator UL148 interacts with SEL1L, a cellular factor that governs endoplasmic reticulum-associated degradation of the viral envelope glycoprotein gO. J. Virol. 2018, 92, e00688-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luganini, A.; Cavaletto, N.; Raimondo, S.; Geuna, S.; Gribaudo, G. Loss of the human cytomegalovirus US16 protein abrogates virus entry into endothelial and epithelial cells by reducing the virion content of the pentamer. J. Virol. 2017, 91, e00205-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrell, I.; Wilkie, G.S.; Davison, A.J.; Statkute, E.; Fielding, C.A.; Tomasec, P.; Wilkinson, G.W.; Stanton, R.J. Genetic stability of bacterial artificial chromosome-derived human cytomegalovirus during culture In vitro. J. Virol. 2016, 90, 3929–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerna, G.; Percivalle, E.; Perez, L.; Lanzavecchia, A.; Lilleri, D. Monoclonal antibodies to different components of the human cytomegalovirus (HCMV) pentamer gH/gL/pUL128L and trimer gH/gL/gO as well as antibodies elicited during primary HCMV infection prevent epithelial cell syncytium formation. J. Virol. 2016, 90, 6216–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertel, L.; Mocarski, E.S. Global analysis of host cell gene expression late during cytomegalovirus infection reveals extensive dysregulation of cell cycle gene expression and induction of Pseudomitosis independent of US28 function. J. Virol. 2004, 78, 11988–12011. [Google Scholar] [CrossRef] [Green Version]
- Suarez, N.M.; Wilkie, G.S.; Hage, E.; Camiolo, S.; Holton, M.; Hughes, J.; Maabar, M.; Vattipally, S.B.; Dhingra, A.; Gompels, U.A.; et al. Human cytomegalovirus genomes sequenced directly from clinical material: Variation, multiple-strain infection, recombination, and gene loss. J. Infect. Dis. 2019, 220, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Pari, G.S.; Anders, D.G. Eleven loci encoding trans-acting factors are required for transient complementation of human cytomegalovirus oriLyt-dependent DNA replication. J. Virol. 1993, 67, 6979–6988. [Google Scholar] [CrossRef] [Green Version]
- Bronzini, M.; Luganini, A.; Dell’Oste, V.; De Andrea, M.; Landolfo, S.; Gribaudo, G. The US16 gene of human cytomegalovirus is required for efficient viral infection of endothelial and epithelial cells. J. Virol. 2012, 86, 6875–6888. [Google Scholar] [CrossRef] [Green Version]
- Dargan, D.J.; Douglas, E.; Cunningham, C.; Jamieson, F.; Stanton, R.J.; Baluchova, K.; McSharry, B.P.; Tomasec, P.; Emery, V.C.; Percivalle, E.; et al. Sequential mutations associated with adaptation of human cytomegalovirus to growth in cell culture. J. Gen. Virol. 2010, 91, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Murrell, I.; Bedford, C.; Ladell, K.; Miners, K.L.; Price, D.A.; Tomasec, P.; Wilkinson, G.W.G.; Stanton, R.J. The pentameric complex drives immunologically covert cell-cell transmission of wild-type human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2017, 114, 6104–6109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calo, S.; Cortese, M.; Ciferri, C.; Bruno, L.; Gerrein, R.; Benucci, B.; Monda, G.; Gentile, M.; Kessler, T.; Uematsu, Y.; et al. The human cytomegalovirus UL116 gene encodes an envelope glycoprotein forming a complex with gH independently from gL. J. Virol. 2016, 90, 4926–4938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, J.C.; Beesley, J.E.; Stern, H. Syncytium formation caused by human cytomegalovirus in human embryonic lung fibroblasts. Arch. Virol. 1978, 57, 143–152. [Google Scholar] [CrossRef]
- Michelson-Fiske, S.; Arnoult, J.; Febvre, H. Cytomegalovirus infection of human lung epithelial cells in vitro. Intervirology 1975, 5, 354–363. [Google Scholar] [CrossRef]
- Vonka, V.; Anisimova, E.; Macek, M. Replication of cytomegalovirus in human epitheloid diploid cell line. Arch. Virol. 1976, 52, 283–296. [Google Scholar] [CrossRef]
- Knowles, W.A. In-vitro cultivation of human cytomegalovirus in thyroid epithelial cells. Arch. Virol. 1976, 50, 119–124. [Google Scholar] [CrossRef]
- Ho, K.L.; Gottlieb, C.; Zarbo, R.J. Cytomegalovirus infection of cerebral astrocytoma in an AIDS patient. Clin. Neuropathol. 1991, 10, 127–133. [Google Scholar] [CrossRef]
- Amidzic, J.; Vuckovic, N.; Capo, I.; Levakov, A.F. Congenital cytomegalic inclusion disease with disseminated Herpes simplex infection. Malays. J. Pathol. 2019, 41, 75–78. [Google Scholar]
- Patterson, J.W.; Broecker, A.H.; Kornstein, M.J.; Mills, A.S. Cutaneous cytomegalovirus infection in a liver transplant patient. Diagnosis by in situ DNA hybridization. Am. J. Dermatopathol. 1988, 10, 524–530. [Google Scholar] [CrossRef]
- Dancygier, H. Clinical Hepatology: Principles and Practice of Hepatobiliary Disease; Springer: Berlin/Heidelberg, Germany, 2009; Volume 2. [Google Scholar]
- Podolski, D.; Camilleri, M.; Fitz, J.; Kalloo, A.; Shanahan, F.; Wang, T. Yamada’s Textbook of Gastroenterology; John Wiley & Sons: Hoboken, NJ, USA, 2015; Volume 2. [Google Scholar]
- Rosa, R. Cytomegalovirus (CMV) Retinitis. 2020. Available online: https://www.aao.org/image/cytomegalovirus-cmv-retinitis (accessed on 23 April 2020).
- Tugizov, S.; Navarro, D.; Paz, P.; Wang, Y.; Qadri, I.; Pereira, L. Function of human cytomegalovirus glycoprotein B: Syncytium formation in cells constitutively expressing gB is blocked by virus-neutralizing antibodies. Virology 1994, 201, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Tugizov, S.; Wang, Y.; Qadri, I.; Navarro, D.; Maidji, E.; Pereira, L. Mutated forms of human cytomegalovirus glycoprotein B are impaired in inducing syncytium formation. Virology 1995, 209, 580–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maidji, E.; Tugizov, S.; Jones, T.; Zheng, Z.; Pereira, L. Accessory human cytomegalovirus glycoprotein US9 in the unique short component of the viral genome promotes cell-to-cell transmission of virus in polarized epithelial cells. J. Virol. 1996, 70, 8402–8410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanarsdall, A.L.; Chase, M.C.; Johnson, D.C. Human cytomegalovirus glycoprotein gO complexes with gH/gL, promoting interference with viral entry into human fibroblasts but not entry into epithelial cells. J. Virol. 2011, 85, 11638–11645. [Google Scholar] [CrossRef] [Green Version]
- Vanarsdall, A.L.; Ryckman, B.J.; Chase, M.C.; Johnson, D.C. Human cytomegalovirus glycoproteins gB and gH/gL mediate epithelial cell-cell fusion when expressed either in cis or in trans. J. Virol. 2008, 82, 11837–11850. [Google Scholar] [CrossRef] [Green Version]
- Vanarsdall, A.L.; Pritchard, S.R.; Wisner, T.W.; Liu, J.; Jardetzky, T.S.; Johnson, D.C. CD147 promotes entry of pentamer-expressing human cytomegalovirus into epithelial and endothelial cells. MBio 2018, 9, e00781-18. [Google Scholar] [CrossRef] [Green Version]
- Ciferri, C.; Chandramouli, S.; Donnarumma, D.; Nikitin, P.A.; Cianfrocco, M.A.; Gerrein, R.; Feire, A.L.; Barnett, S.W.; Lilja, A.E.; Rappuoli, R.; et al. Structural and biochemical studies of HCMV gH/gL/gO and Pentamer reveal mutually exclusive cell entry complexes. Proc. Natl. Acad Sci. USA 2015, 112, 1767–1772. [Google Scholar] [CrossRef] [Green Version]
- Kinzler, E.R.; Compton, T. Characterization of human cytomegalovirus glycoprotein-induced cell-cell fusion. J. Virol. 2005, 79, 7827–7837. [Google Scholar] [CrossRef] [Green Version]
- Noriega, V.M.; Gardner, T.J.; Redmann, V.; Bongers, G.; Lira, S.A.; Tortorella, D. Human cytomegalovirus US28 facilitates cell-to-cell viral dissemination. Viruses 2014, 6, 1202–1218. [Google Scholar] [CrossRef]
- Wang, D.; Shenk, T. Human cytomegalovirus UL131 open reading frame is required for epithelial cell tropism. J. Virol. 2005, 79, 10330–10338. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vo, M.; Aguiar, A.; McVoy, M.A.; Hertel, L. Cytomegalovirus Strain TB40/E Restrictions and Adaptations to Growth in ARPE-19 Epithelial Cells. Microorganisms 2020, 8, 615. https://doi.org/10.3390/microorganisms8040615
Vo M, Aguiar A, McVoy MA, Hertel L. Cytomegalovirus Strain TB40/E Restrictions and Adaptations to Growth in ARPE-19 Epithelial Cells. Microorganisms. 2020; 8(4):615. https://doi.org/10.3390/microorganisms8040615
Chicago/Turabian StyleVo, Mai, Alexis Aguiar, Michael A. McVoy, and Laura Hertel. 2020. "Cytomegalovirus Strain TB40/E Restrictions and Adaptations to Growth in ARPE-19 Epithelial Cells" Microorganisms 8, no. 4: 615. https://doi.org/10.3390/microorganisms8040615