Unraveling Heterogeneity of Coral Microbiome Assemblages in Tropical and Subtropical Corals in the South China Sea


Abstract
:1. Introduction
2. Materials and Methods
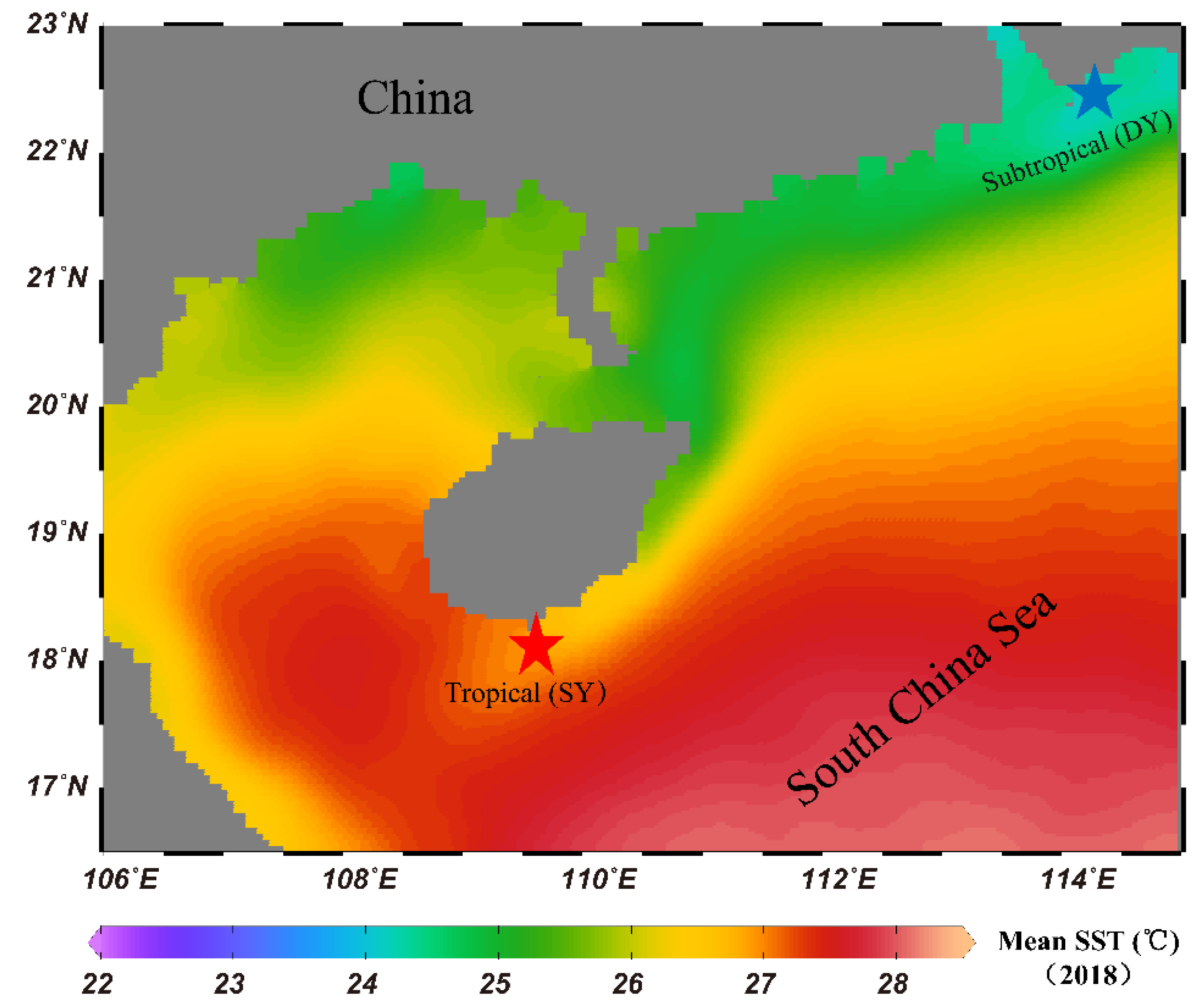
2.1. Sampling Regions and Sample Collection
2.2. Environmental Data Collection
2.3. DNA Extraction, Amplification, Pyrosequencing and Data Processing
2.4. Network Analysis and Functional Profiling of Coral Microbiome
2.5. Multivariate Statistics Analyses
2.6. Availability of Data and Materials
3. Results
3.1. Environmental Parameters at the Study Regions
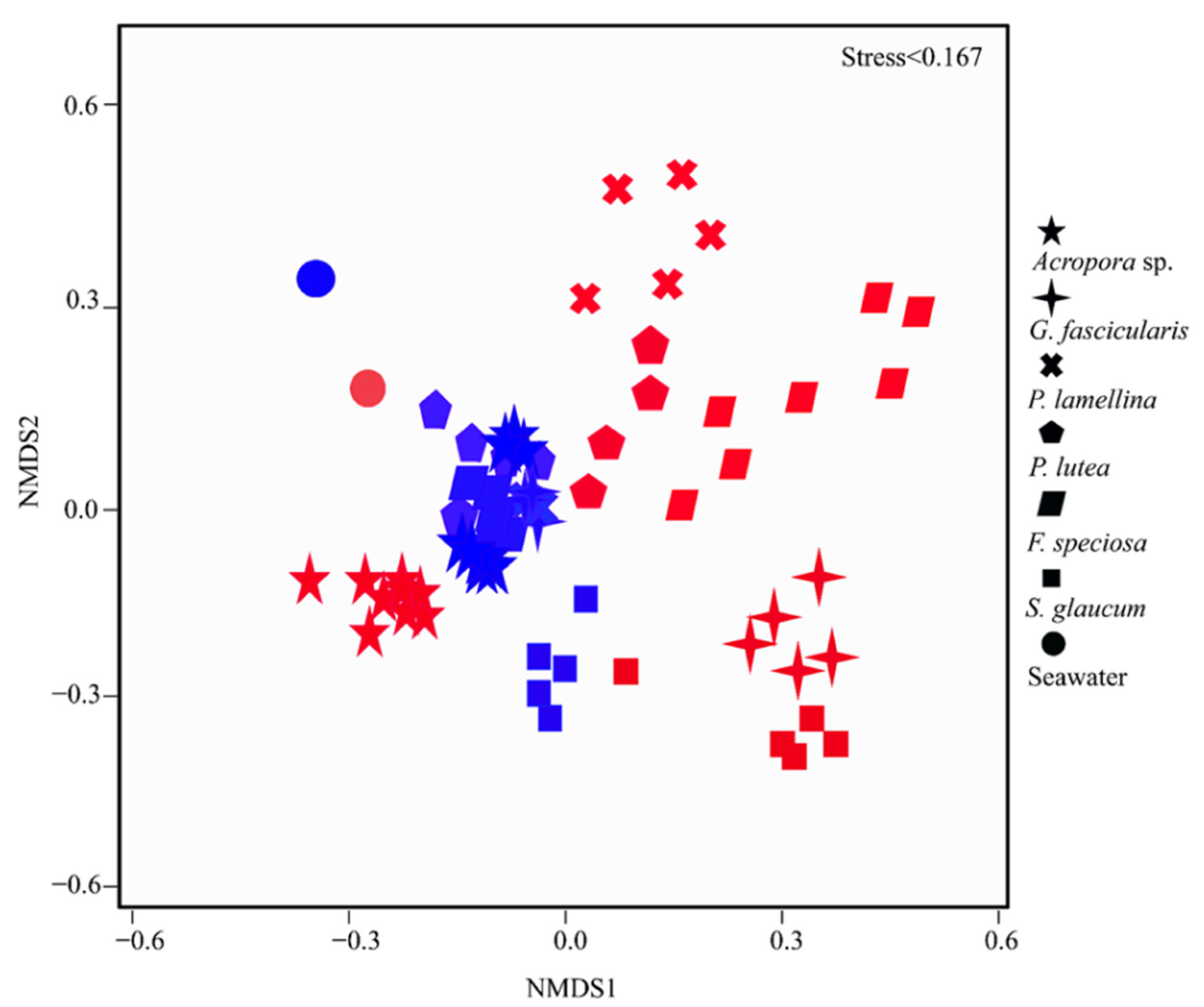
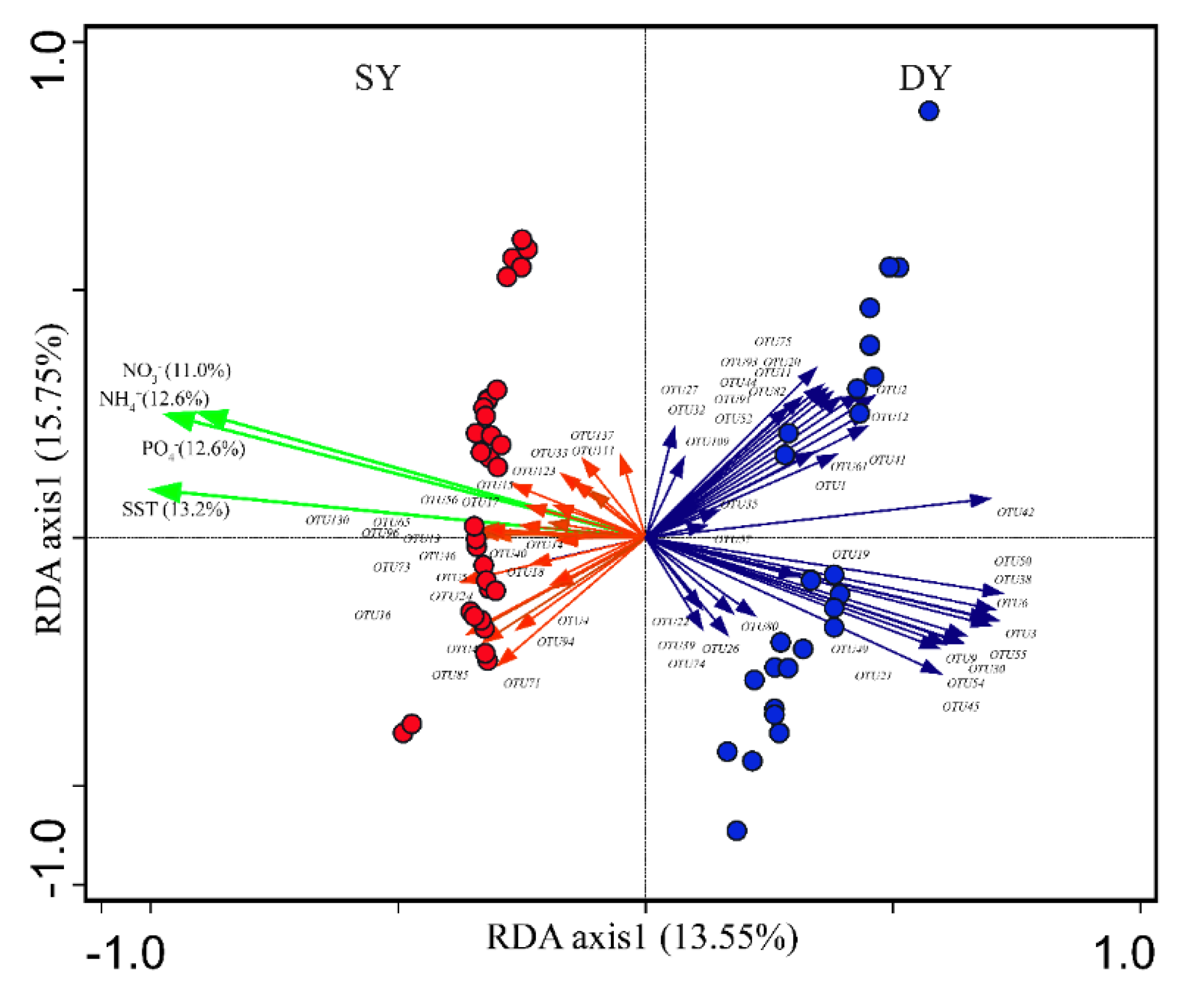
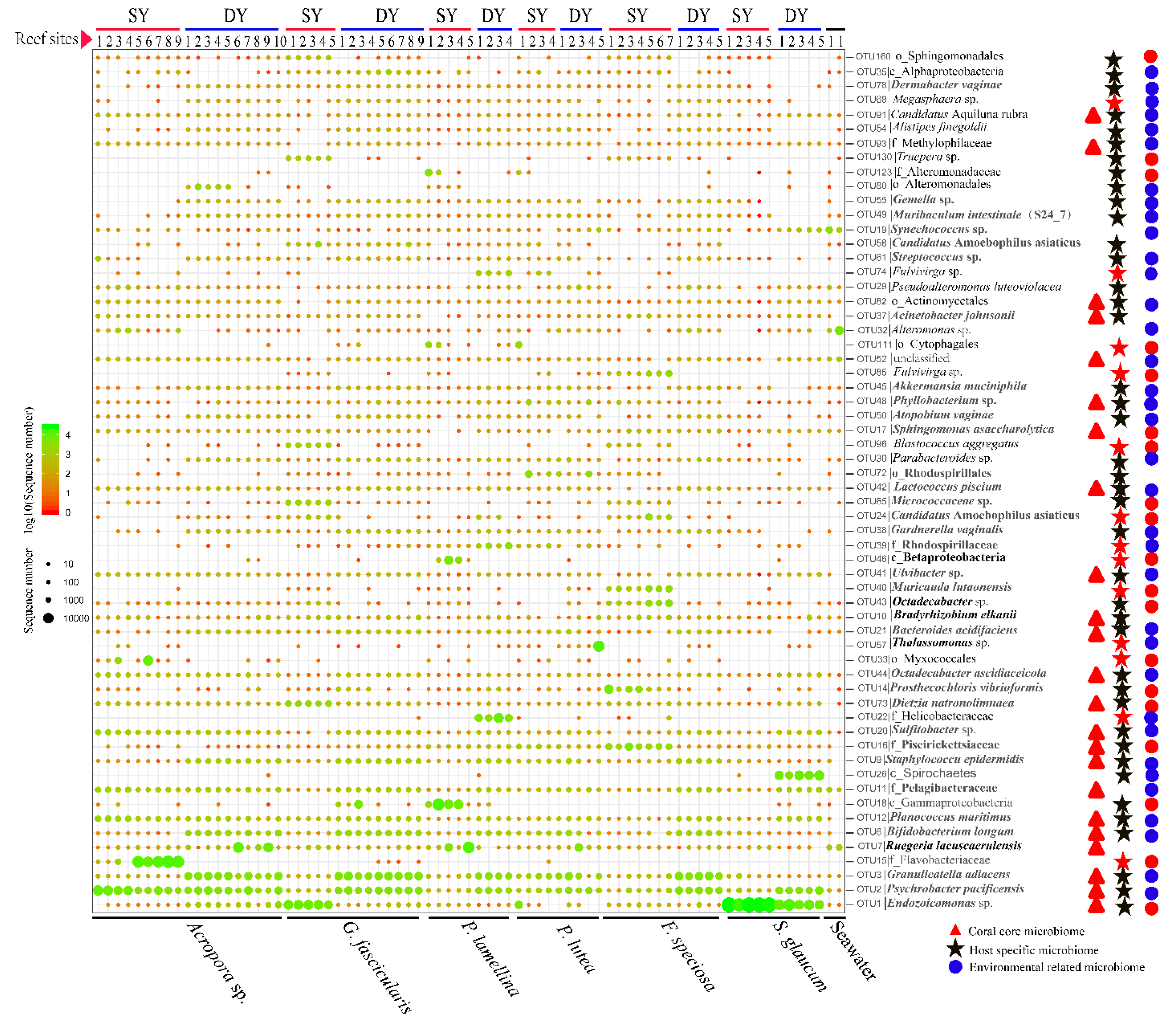
3.2. Coral Microbiomes and Their Relationships with the Host and Environment
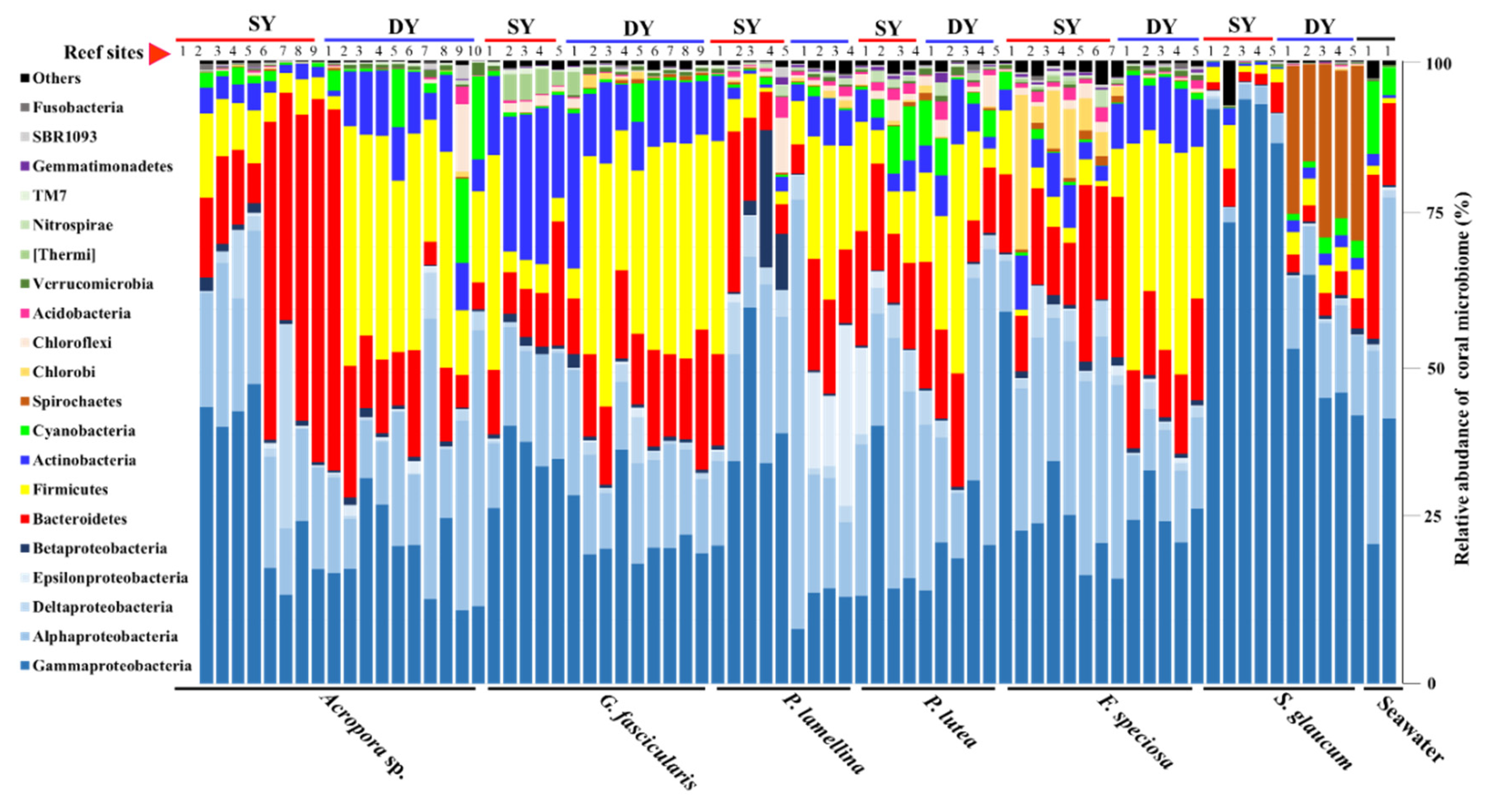
3.3. Diversity and Functional Groups of Coral Microbiome
3.4. Microbial Functional Profiles Change Between the Tropical and Subtropical Reefs
4. Discussion
4.1. Effects of Hosts and Thermal Regimes on Coral Microbiome Assemblages
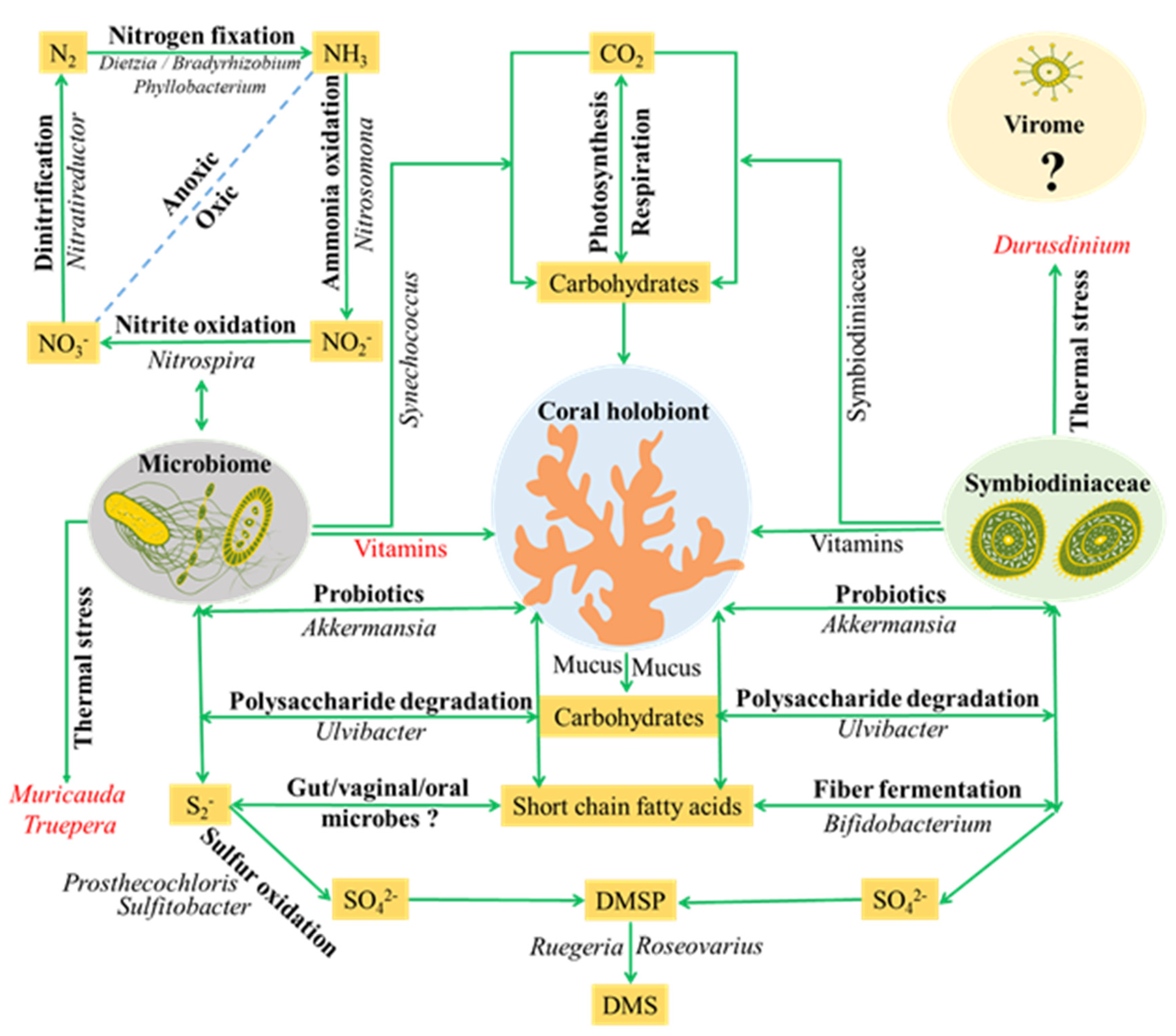
4.2. Major Members and Potential Roles of Coral Microbiome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Coral reef ecosystems and anthropogenic climate change. Reg. Environ. Change 2010, 11, 215–227. [Google Scholar] [CrossRef]
- Carpenter, K.E.; Abrar, M.; Aeby, G.; Aronson, R.B.; Banks, S.; Bruckner, A.; Chiriboga, A.; Cortés, J.; Delbeek, J.C.; DeVantieret, L.; et al. One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 2008, 321, 560–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowlton, N.; Rohwer, F. Multispecies microbial mutualisms on coral reefs: The host as a habitat. Am. Nat. 2003, 162, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Bosch, T.C.; McFall-Ngai, M.J. Metaorganisms as the new frontier. Zoology 2011, 114, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Pantos, O.; Bongaerts, P.; Dennis, P.G.; Tyson, G.W.; Hoegh-Guldberg, O. Habitat-specific environmental conditions primarily control the microbiomes of the coral Seriatopora hystrix. ISME. J. 2015, 9, 1916–1927. [Google Scholar] [CrossRef]
- Trench, R.K. The physiology and biochemistry of zooxanthellae symbiotic with marine coelenterates. ii. liberation of fixed formula C by zooxanthellae in vitro. Proc. Biol. Sci. 1971, 177, 237–250. [Google Scholar]
- Hernandez-Agreda, A.; Leggat, W.; Bongaerts, P.; Ainsworth, T.D. The microbial signature provides insight into the mechanistic basis of coral success across reef habitats. mBio. 2016, 7, e00560-16. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the core microbiome in corals’ microbial soup. Trends. Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef]
- Tremblay, P.; Weinbauer, M.G.; Rottier, C.; Guerardel, Y.; Nozais, C.; Ferrier-Pages, C. Mucus composition and bacterial communities associated with the tissue and skeleton of three scleractinian corals maintained under culture conditions. J. Mar. Biol. Assoc. UK. 2011, 91, 649–657. [Google Scholar] [CrossRef]
- Lee, O.O.; Yang, J.; Bougouffa, S.; Wang, Y.; Qian, P.Y. Spatial and species variations in bacterial communities associated with corals from the red sea as revealed by pyrosequencing. Appl. Environ. Microbiol. 2012, 78, 7173–7184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, K.M.; Moss, A.G.; Chadwick, N.E.; Liles, M.R. Bacterial associates of two Caribbean coral species reveal species-specific distribution and geographic variability. Appl. Environ. Microbiol. 2012, 78, 6438–6449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlos, C.; Torres, T.T.; Ottoboni, L.M.M. Bacterial communities and species-specific associations with the mucus of Brazilian coral species. Sci. Rep. 2013, 3, 1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.D.; Brown, B.E.; Putchim, L.; Sweet, M.J. Age-related shifts in bacterial diversity in a reef coral. PLoS One 2015, 10, e0144902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hester, E.R.; Barott, K.L.; Nulton, J.; Vermeij, M.J.; Rohwer, F.L. Stable and sporadic symbiotic communities of coral and algal holobionts. ISME J. 2016, 10, 1157–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, D.; Iida, Y.; Uthicke, S.; Smith-Keune, C. Changes in coral-associated microbial communities during a bleaching event. ISME J. 2007, 2, 350–363. [Google Scholar] [CrossRef]
- Roder, C.; Arif, C.; Daniels, C.; Weil, E.; Voolstra, C.R. Bacterial profiling of White Plague Disease across corals and oceans indicates a conserved and distinct disease microbiome. Mol. Ecol. 2014, 23, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Roder, C.; Bayer, T.; Aranda, M.; Kruse, M.; Voolstra, C.R. Microbiome structure of the fungid coral Ctenactis echinata aligns with environmental differences. Mol. Ecol. 2015, 24, 3501–3511. [Google Scholar] [CrossRef] [Green Version]
- Brener-Raffalli, K.; Clerissi, C.; Vidaldupiol, J.; Adjeroud, M.; Bonhomme, F.; Pratlong, M.; Aurelle, D.; Mitta, G.; Toulza, E. Thermal regime and host clade, rather than geography, drive Symbiodinium and bacterial assemblages in the scleractinian coral Pocillopora damicornis sensu lato. Microbiome. 2018, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.F. Coral reefs in the South China Sea: Their response to and records on past environmental changes. Science China Earth Sciences. 2012, 55, 1217–1229. [Google Scholar] [CrossRef]
- Huang, D.; Licuanan, W.Y.; Hoeksema, B.W.; Chen, C.A.; Ang, P.O.; Huang, H.; Lane, D.J.M.; Vo, S.T.; Waheed, Z.; Affendi, Y.M.; et al. Extraordinary diversity of reef corals in the South China Sea. Mar. Biodivers. 2015, 45, 157–168. [Google Scholar] [CrossRef]
- Li, S.; Yu, K.; Shi, Q.; Chen, T.; Zhao, M.; Zhao, J. Interspecies and spatial diversity in the symbiotic zooxanthellae density in corals from northern South China Sea and its relationship to coral reef bleaching. Chin. Sci. Bull. 53, 295–303. [CrossRef]
- Clark, T.R.; Chen, X.F.; Leonard, N.D.; Liu, F.Y.; Guo, Y.R.; Zeng, T.; Wei, G.J.; Zhao, J.X. Episodic coral growth in China’s subtropical coral communities linked to broad-scale climatic change. Geology 2018, 47, 79–82. [Google Scholar] [CrossRef]
- Beman, J.M.; Roberts, K.J.; Wegley, L.; Rohwer, F.; Francis, C.A. Distribution and diversity of archaeal ammonia monooxygenase genes associated with corals. Appl. Environ. Microbiol. 2007, 73, 5642–5647. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Tian, R.M.; Zhou, G.W.; Tong, H.Y.; Wong, Y.H.; Zhang, W.P.; Chui, A.P.Y.; Xie, J.Y.; Qiu, J.W.; Ang, P.O.; et al. Exploring coral microbiome assemblages in the South China Sea. Sci. Rep. 2018, 8, 2428. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Meth. 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 2010, 26, 2460. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; Desantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics. 2010, 26, 266–267. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Tu, Q.; Zhi, X. Functional Molecular Ecological Networks. mBio 2010, 1, e00169-10. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J. Finding community structure in networks using the eigenvectors of matrices. Phys. Rev. E 2006, 74, 036104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 17 April 2020).
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; Mcdonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchinet, P.R.; O’Hara, R.B. Vegan: Community Ecology Package. R Package Version 2.3--5. Available online: http://CRAN.R-project.org/package=vegan (accessed on 17 April 2020).
- Poulin, R.; Krasnov, B.R.; Shenbrot, G.I.; Mouillot, D.; Khokhlova, I.S. Evolution of host specificity in fleas: Is it directional and irreversible? Int. J. Parasitol. 2006, 36, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, M.; Seneca, F.O.; Yum, L.K.; Palumbi, S.R.; Voolstra, C.R. Bacterial community dynamics are linked to patterns of coral heat tolerance. Nat. Commun. 2017, 8, 14213. [Google Scholar] [CrossRef] [Green Version]
- Lajeunesse, T.C.; Bhagooli, R.; Hidaka, M.; Devantier, L.; Done, T.; Schmidt, G.W.; Fitt, W.K.; Hoegh-Guldberg, O. Closely related Symbiodinium spp. differ in relative dominance in coral reef host communities across environmental, latitudinal and biogeographic gradients. Mar. Eco. Prog. Ser. 2004, 284, 147–161. [Google Scholar] [CrossRef]
- Stat, M.; Pochon, X.; Cowie, R.O.M.; Gates, R.D. Specificity in communities of Symbiodinium in corals from johnston atoll. Mar. Eco. Prog. Ser. 2009, 386, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.Q.; Chai, G.J.; Xiao, Y.L.; Xu, L.J.; Yu, K.F.; Li, J.L.; Liu, F.; Cheng, H.; Zhang, F.L.; Liao, B.L.; et al. Flexible symbiotic associations of Symbiodinium with five typical coral species in tropical and subtropical reef regions of the northern South China Sea. Front. Microbiol. 2018, 9, 2485. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshwater. Res. 1999, 50, 839. [Google Scholar] [CrossRef] [Green Version]
- Zaneveld, J.R.; Mcminds, R.; Vega Thurber, R. Stress and stability: Applying the anna karenina principle to animal microbiomes. Nat. Microbiol. 2017, 2, 17121. [Google Scholar] [CrossRef]
- Venn, A.A.; Loram, J.E.; Douglas, A.E. Photosynthetic symbioses in animals. J. Exp. Bot. 2008, 59, 1069–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, M.J.; Palenik, B. Niche adaptation in ocean cyanobacteria. Nature 1998, 396, 226–228. [Google Scholar] [CrossRef]
- Jeong, H.J.; Yoo, Y.D.; Kang, N.S.; Lim, A.S.; Seong, K.A.; Lee, S.Y.; Lee, M.J.; Lee, K.H.; Kim, H.S.; Shin, W.; et al. Heterotrophic feeding as a newly identified survival strategy of the dinoflagellate Symbiodinium. PNAS 2012, 109, 12604–12609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedashkovskaya, O.I.; Kukhlevskiy, A.D.; Zhukova, N.V.; Kim, S.B. Amylibacter ulvae sp. nov. a new alphaproteobacterium isolated from the pacific green alga Ulva fenestrata. Arch. Microbiol. 2016, 198, 251–256. [Google Scholar] [CrossRef]
- Antoun, H.; Beauchamp, C.J.; Goussard, N.; Chabot, R.; Lalande, R. Potential of rhizobium and bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effect on radishes (Raphanus sativus L.). Plant Soil. 1998, 204, 57–67. [Google Scholar] [CrossRef]
- Rädecker, N.; Pogoreutz, C.; Voolstra, C.R.; Wiedenmann, J.; Wild, C. Nitrogen cycling in corals: The key to understanding holobiont functioning? Trends Microbiol. 2015, 23, 490–497. [Google Scholar] [CrossRef] [Green Version]
- Krishnani, K.K.; Kathiravan, V.; Natarajan, M.; Kailasam, M.; Pillai, S.M. Diversity of sulfur-oxidizing bacteria in greenwater system of coastal aquaculture. Appl. Biochem. Biotech. 2010, 162, 1225–1237. [Google Scholar] [CrossRef]
- Miller, A.W.; Richardson, L.L. Fine structure analysis of black band disease (BBD) infected coral and coral exposed to the BBD toxins microcystin and sulfide. J. Invertebr. Pathol. 2012, 109, 27–33. [Google Scholar] [CrossRef]
- Raina, J.B.; Tapiolas, D.M.; Forêt, S.; Lutz, A.; Abrego, D.; Ceh, J.; Seneca, F.O.; Clode, P.L.; Bourne, D.G.; Willis, B.L.; et al. DMSP biosynthesis by an animal and its role in coral thermal stress response. Nature 2013, 502, 677–680. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Soutome, S.; Oho, T. Identification and characterization of a fibronectin-binding protein from Granulicatella adiacens. Oral. Microbiol. Immun. 2011, 26, 353–364. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature. 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Vaneechoutte, M. The human vaginal microbial community. Res. Microbiol. 2017, 168, 811–825. [Google Scholar] [CrossRef]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The human gut microbiome: From association to modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef]
- Schmitzesser, S.; Tischler, P.; Arnold, R.; Montanaro, J.; Wagner, M.; Rattei, T.; Hornet, M. The genome of the amoeba symbiont “Candidatus Amoebophilus asiaticus” reveals common mechanisms for host cell interaction among amoeba-associated bacteria. J. Bacteriol. 2010, 192, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; Weber, L.G.; Santoro, A.E.; Webster, N.S. Distinguishing between microbial habitats unravels ecological complexity in coral microbiomes. mSystems. 2016, 1, e00143-16. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.O.; Park, I.S.; Park, S.; Nam, B.H.; Yoon, J.H. Octadecabacter ascidiaceicola sp. nov. isolated from sea squirt Halocynthia roretzi. Int. J. Syst. Evol. Microbiol. 2016, 66, 296–301. [Google Scholar] [CrossRef]
- Britton, A.P.; Zabek, E.; Schwantje, H. Pleural actinomycosis in a free-ranging river otter (Lontra canadensis). J. Zoo Wildl. Med. Off. Publ. Am. Assoc. Zoo Vet. 2011, 42, 751. [Google Scholar] [CrossRef]
- Kemp, D.W.; Hernandez-Pech, X.; Iglesias-Prieto, R.; Fitt, W.K.; Schmidt, G.W. Community dynamics and physiology of Symbiodinium spp. before, during, and after a coral bleaching event. Limnol. Oceanogr. 2014, 59, 788–797. [Google Scholar] [CrossRef]
- Arun, A.B.; Chen, W.M.; Lai, W.A.; Chao, J.H.; Rekha, P.D.; Shen, F.T.; Singh, S.; Young, C.C. Muricauda lutaonensis sp. nov. a moderate thermophile isolated from a coastal hot spring. Int. J. Syst. Evol. Micr. 2009, 59, 2738–2742. [Google Scholar] [CrossRef]
- Albuquerque, L.; Simões, C.; Nobre, M.F.; Pino, N.M.; Battista, J.R.; Silva, M.T.; Rainey, F.A.; da Costaa, M.S. Truepera radiovictrix gen. nov. sp. nov. a new radiation resistant species and the proposal of Trueperaceae fam. nov. FEMS. Microbiol. Lett. 2010, 247, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Ainsworth, T.D.; Krause, L.; Bridge, T.; Torda, G.; Raina, J.B.; Zakrzewski, M.; Gates, R.D.; Padilla-Gamiño, J.L.; Spalding, H.L.; Smith, C. The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. ISME. J. 2015, 9, 2261–2274. [Google Scholar] [CrossRef]
- Neave, M.J.; Rachmawati, R.; Xun, L.; Michell, C.T.; Bourne, D.G.; Apprill, A.; Voolstra, C.R. Differential specificity between closely related corals and abundant Endozoicomonas endosymbionts across global scales. ISME J. 2016, 11, 186–200. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | Climate | Coral | Reef Sites (n) a | Date |
|---|---|---|---|---|
| Sanya Bay (SY) E109.430°–109.490°, N18.210°–18.221° | Tropical | A. sp.b | 9 | 2018-08-25 |
| G. fascicularis | 5 | 2018-08-25 | ||
| P. lamellina | 5 | 2018-08-25 | ||
| P. lutea | 4 | 2018-08-26 | ||
| F. speciosa | 7 | 2018-08-26 | ||
| S. glaucum | 5 | 2018-08-25 | ||
| Daya Bay (DY) E114.411°–114.650°, N22.841°–22.780° | Subtropical | A. sp. | 10 | 2018-08-27 |
| G. fascicularis | 9 | 2018-08-27 | ||
| P. lamellina | 4 | 2018-08-27 | ||
| P. lutea | 5 | 2018-08-27 | ||
| F. speciosa | 5 | 2018-08-27 | ||
| S. glaucum | 5 | 2018-08-27 |
| Nodes | Links | R2 of Power-Law | AvgK a | AvgCC | AvgGD | |
|---|---|---|---|---|---|---|
| Tropical (SY) | 101 | 274 | 0.833 | 5.426 | 0.128 | 3.148 |
| Subtropical (DY) | 184 | 1421 | 0.800 | 15.446 | 0.547 | 4.125 |
| PathwayID | Pathway Description | Variation Trends (SY vs DY) | P Values |
|---|---|---|---|
| PWY-5896 | Vitamin K2 synthesis | ↑ | 6.84 × 10−5 |
| PWY-6897 | Vitamin B1 synthesis | ↓ | 3.32 × 10−4 |
| PYRIDOXSYN-PWY | Vitamin B6 synthesis | ↓ | 1.21 × 10−7 |
| PWY-6519 | Biotin synthesis | ↓ | 3.32 × 10−4 |
| FOLSYN-PWY | Folate synthesis | ↓ | 1.28 × 10−5 |
| HEMESYN2-PWY | Heme b biosynthesis by anaerobic | ↓ | 3.32 × 10−4 |
| PWY-5920 | Heme b biosynthesis from glycine | ↓ | 1.89 × 10−10 |
| NAD-BIOSYNTHESIS-II | NAD biosynthesis | ↓ | 1.36 × 10−8 |
| POLYISOPRENSYN-PWY | Isopenteny biosynthesis (IPP) | ↓ | 1.94 × 10−7 |
| PANTO-PWY | Phosphopantothenate biosynthesis I (coenzyme) | ↓ | 1.90 × 10−11 |
| PWY-5198 | Factor 420 biosynthesis (coenzyme) | ↓ | 4.45 × 10−7 |
| P23-PWY | CO2 fixation by reductive TCA | ↓ | 3.24 × 10−8 |
| PWY-7024 | CO2 fixation by 3-hydroxypropanoate cycle | ↑ | 8.20 × 10−6 |
| PENTOSE-P-PWY | Pentose phosphate pathway (PPP) | ↓ | 2.12 × 10−6 |
| PWY-5659 | GDP-mannose biosynthesis (LPS) | ↑ | 1.13 × 10−5 |
| GALACTARDEG-PWY | D-galactarate (sugar acid) degradation | ↓ | 9.68 × 10−5 |
| PWY-5100 | Pyruvate fermentation to acetate and lactate | ↓ | 5.19 × 10−4 |
| GOLPDLCAT-PWY | Glycerol degradation: propanediol | ↑ | 5.40 × 10−5 |
| PWY-5677 | Succinate fermentation to butanoate | ↓ | 6.37 × 10−8 |
| P122-PWY | Heterolactic fermentation: lactate/CO2/CH3OH | ↑ | 3.37 × 10−5 |
| FASYN-ELONG-PWY | Fatty acid elongation-saturated | ↑ | 2.60 × 10−6 |
| PWY-5971 | Fatty acid biosynthesis: palmitate | ↓ | 2.50 × 10−7 |
| PWY-5529 | Bacteriochlorophyll a biosynthesis | ↓ | 1.21 × 10−8 |
| PWY-5531 | Chlorophyll a biosynthesis | ↑ | 3.24 × 10−4 |
| PWY-6545 | Pyrimidine biosynthesis | ↑ | 6.12 × 10−4 |
| THREOCAT-PWY | L-threonine degradation | ↑ | 4.33 × 10−7 |
| ORNDEG-PWY | L-ornithine degradation (putrescine biosynthesis) | ↓ | 1.05 × 10−6 |
| PWY-5347 | L-methionine biosynthesis (transsulfuration) | ↓ | 4.89 × 10−7 |
| PWY-5345 | L-methionine biosynthesis (by sulfhydrylation) | ↑ | 4.86 × 10−5 |
| HOMOSER-METSYN-PWY | L-methionine biosynthesis I | ↓ | 3.46 × 10−4 |
| PWY-5028 | L-histidine degradation II | ↓ | 3.46 × 10−4 |
| ASPASN-PWY | L-aspartate and L-asparagine biosynthesis | ↓ | 3.28 × 10−8 |
| PWY0-781 | L-aspartate biosynthesis | ↓ | 3.29 × 10−8 |
| PWY0-1338 | Polymyxin resistance (antibiotic resistance) | ↑ | 3.24 × 10−4 |
| PWY-6071 | Aromatic compound degradation: phenylethylamine | ↓ | 5.15 × 10−7 |
| PWY-6210 | Aromatic compound degradation:2-aminophenol degradation | ↓ | 1.36 × 10−4 |
| PWY-5419 | Aromatic compound degradation: catechol | ↑ | 7.74 × 10−6 |
| P621-PWY | Nylon-6 oligomer degradation | ↓ | 3.76 × 10−6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, S.; Jin, X.; Ren, L.; Tan, Y.; Xia, X. Unraveling Heterogeneity of Coral Microbiome Assemblages in Tropical and Subtropical Corals in the South China Sea. Microorganisms 2020, 8, 604. https://doi.org/10.3390/microorganisms8040604
Gong S, Jin X, Ren L, Tan Y, Xia X. Unraveling Heterogeneity of Coral Microbiome Assemblages in Tropical and Subtropical Corals in the South China Sea. Microorganisms. 2020; 8(4):604. https://doi.org/10.3390/microorganisms8040604
Chicago/Turabian StyleGong, Sanqiang, Xuejie Jin, Lijuan Ren, Yehui Tan, and Xiaomin Xia. 2020. "Unraveling Heterogeneity of Coral Microbiome Assemblages in Tropical and Subtropical Corals in the South China Sea" Microorganisms 8, no. 4: 604. https://doi.org/10.3390/microorganisms8040604