Biodegradation of Allethrin by a Novel Fungus Fusarium proliferatum Strain CF2, Isolated from Contaminated Soils

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Enrichment and Isolation of Allethrin-Degrading Fungi
2.3. Identification of the Strain CF2
2.4. Preparation of Fungal Inoculum
2.5. Optimization of the Degradation Conditions for Strain CF2
2.6. Degradation Kinetics of Allethrin with Strain CF2
2.7. Extraction and HPLC Analysis
2.8. Kinetic Analysis
2.9. Statistical Analysis
3. Results
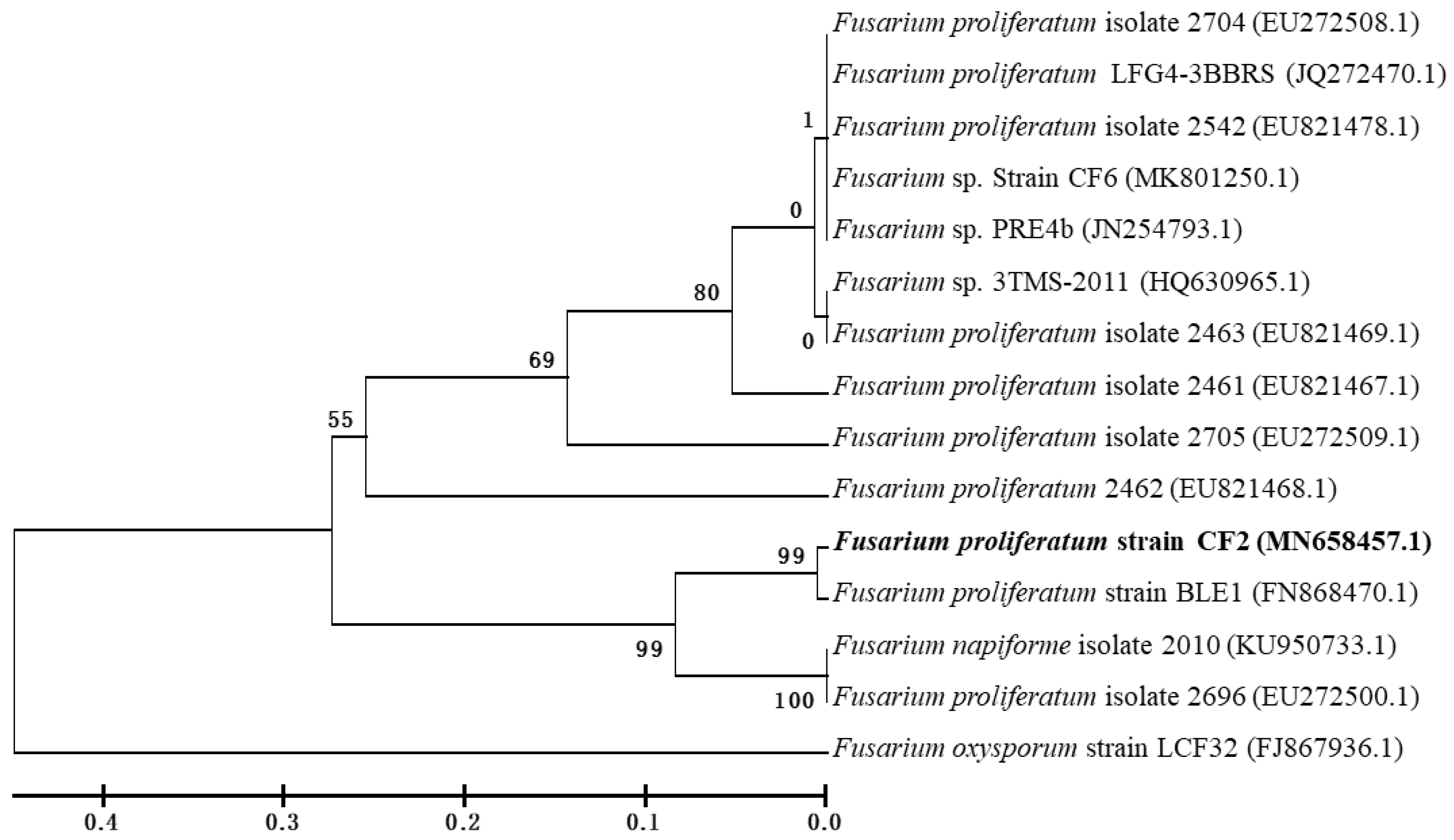
3.1. Isolation and Identification of Strain CF2
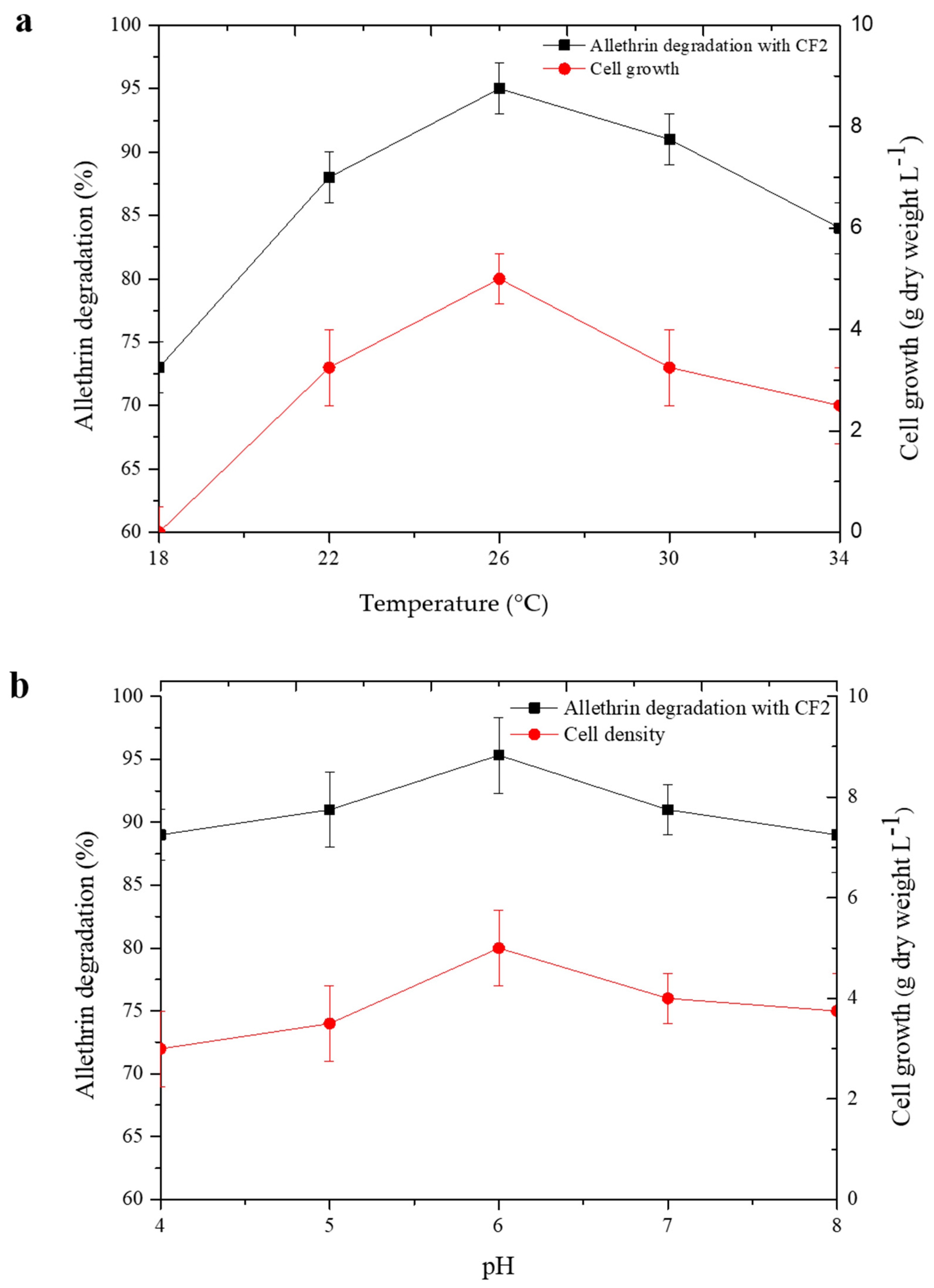
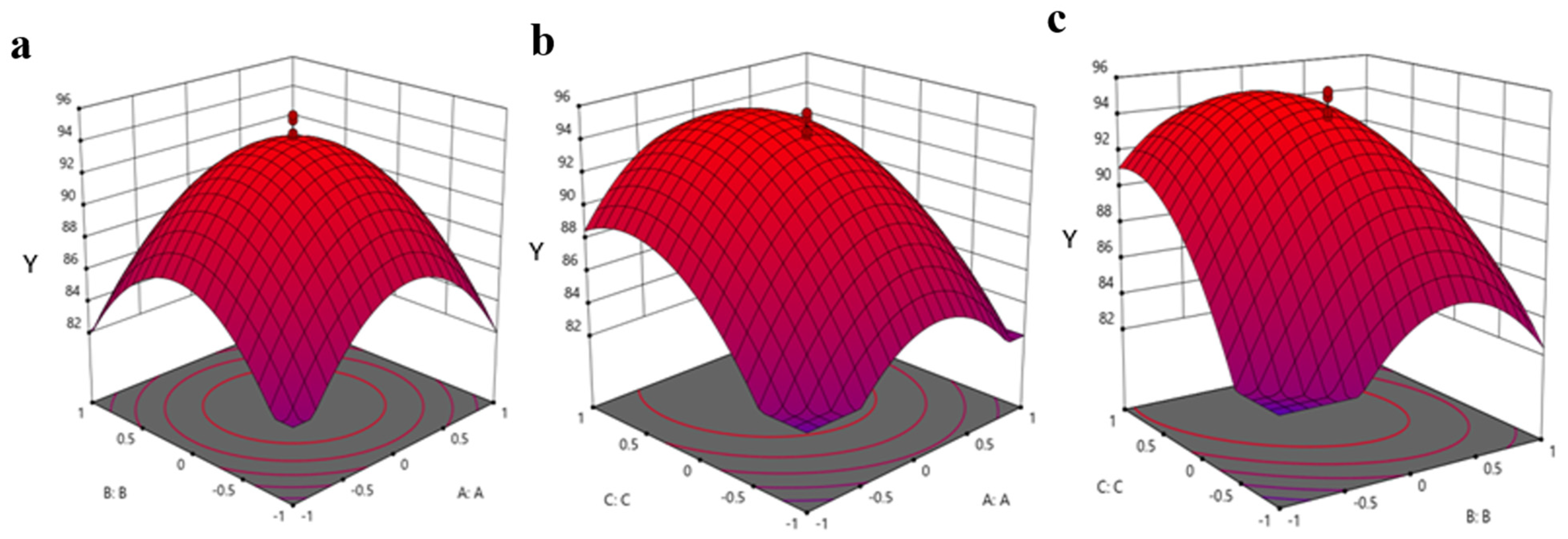
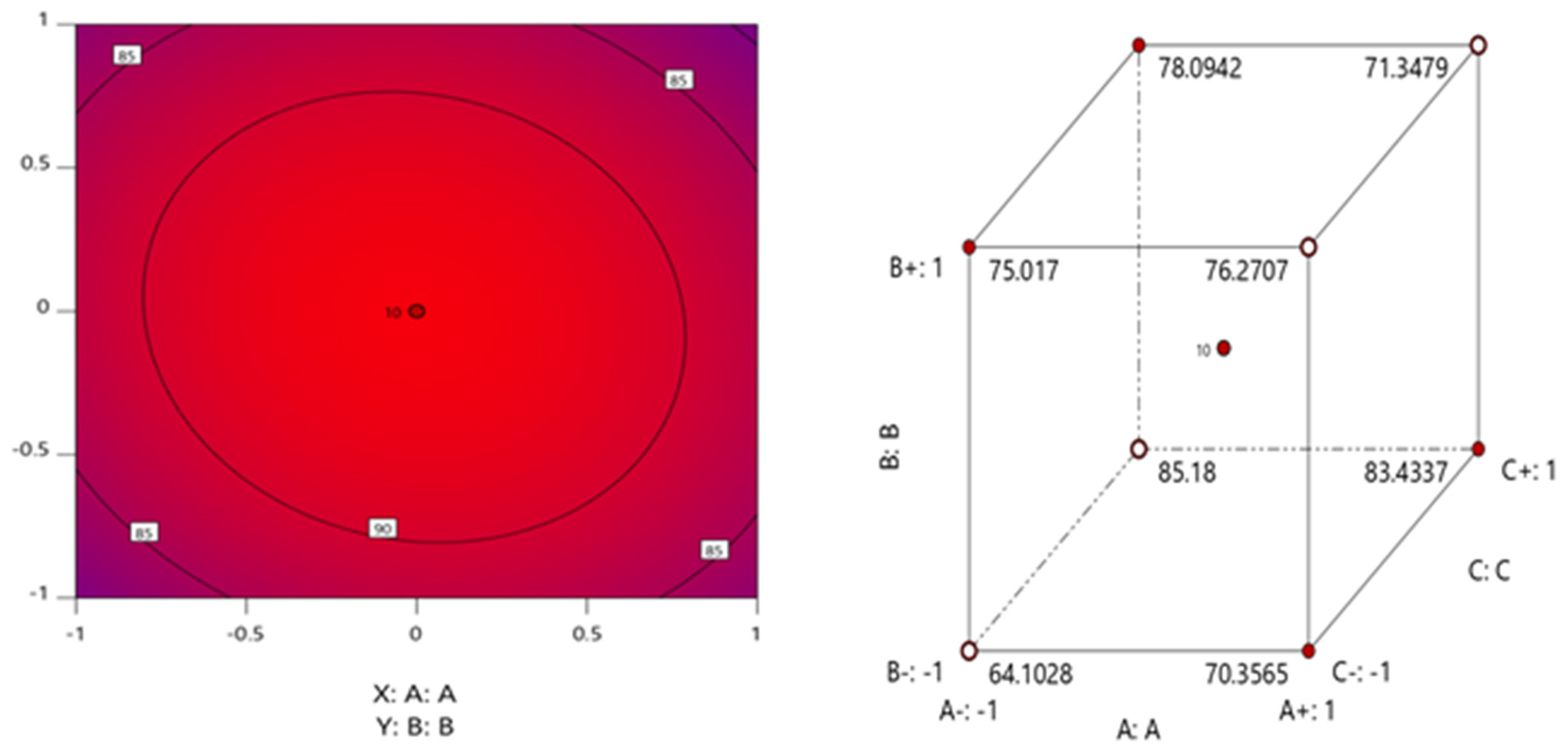
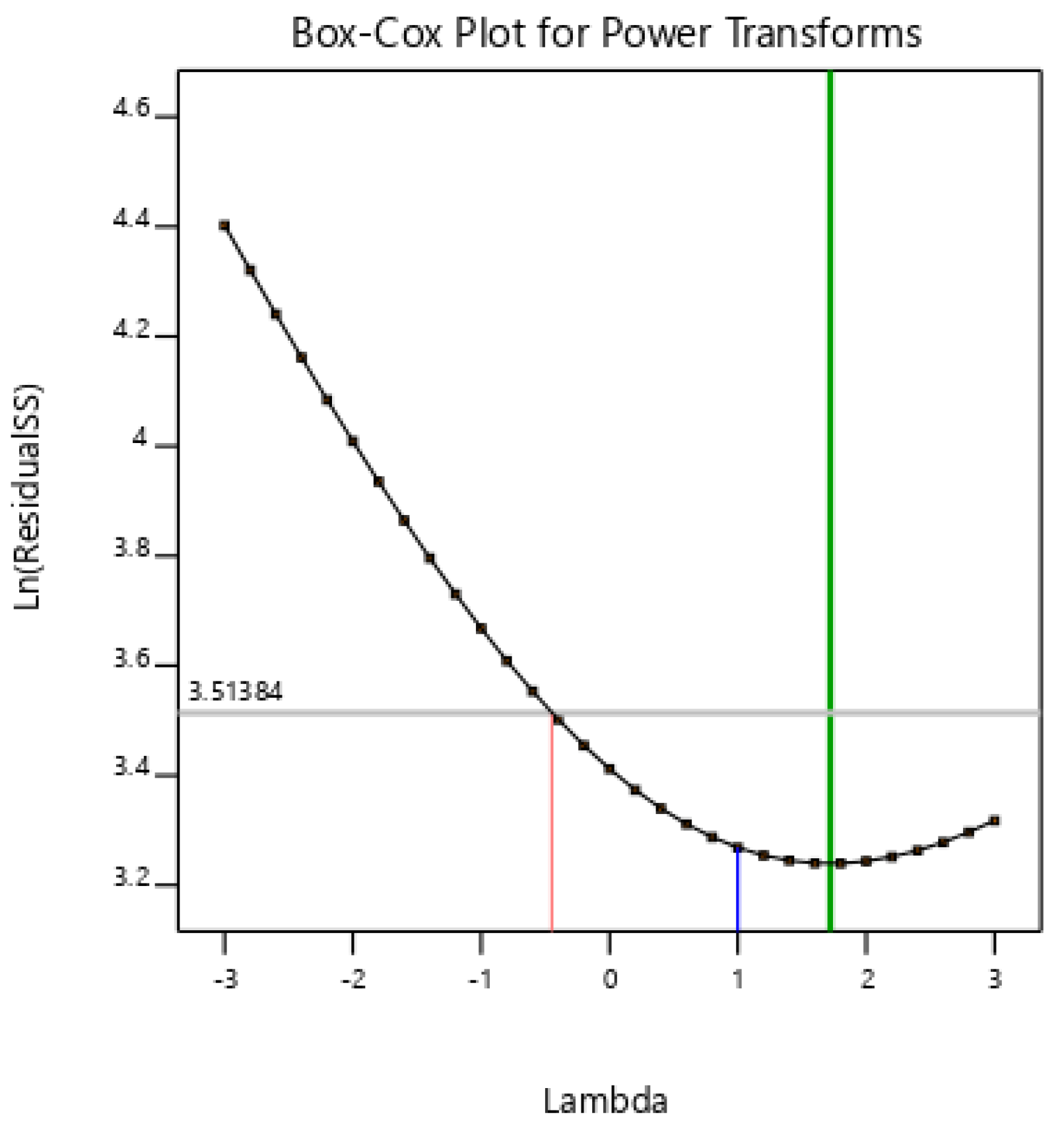
3.2. Optimization of Allethrin Degradation Conditions
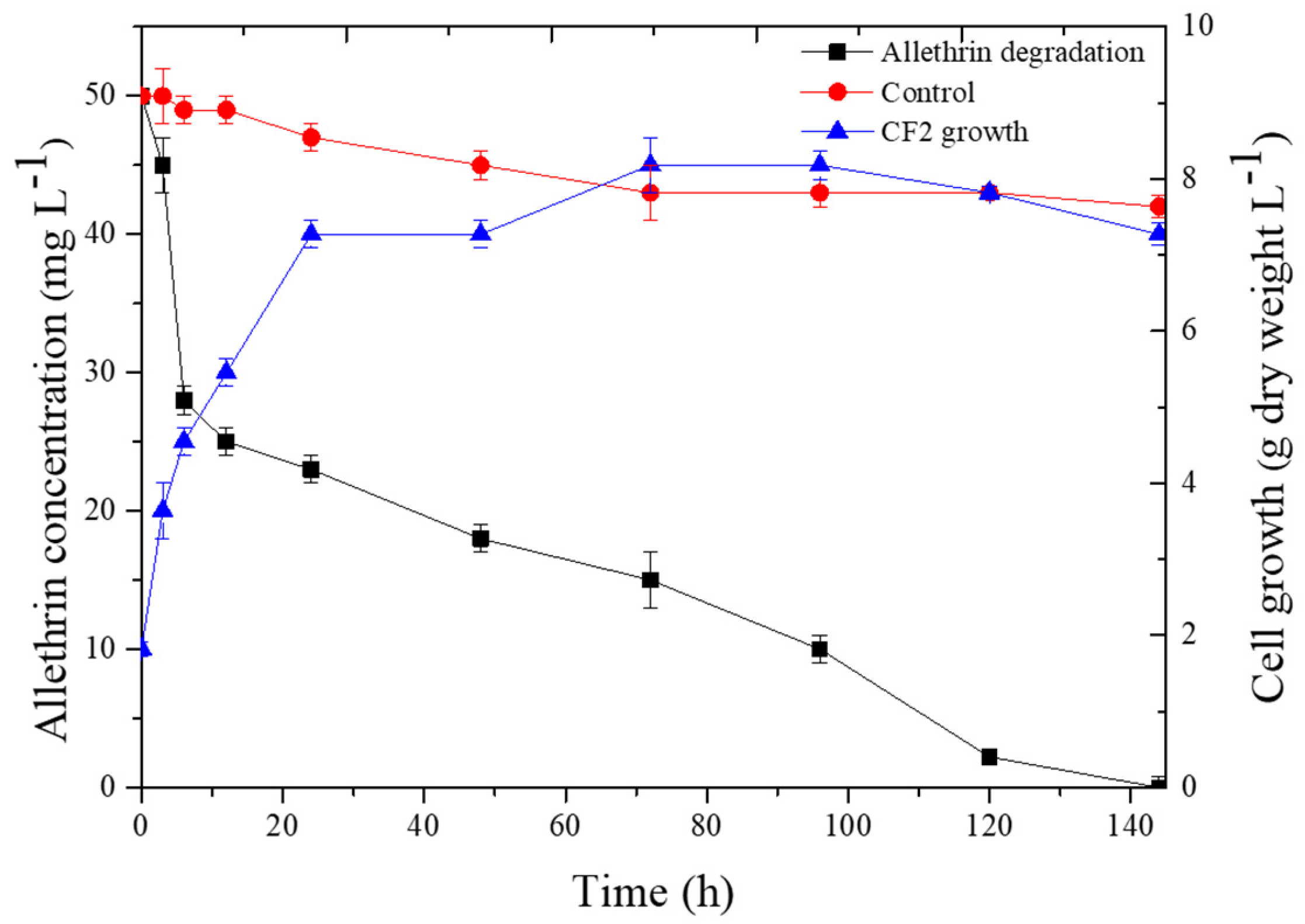
3.3. Degradation Kinetics of Allethrin with Strain CF2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wolansky, M.J.; Harrill, J.A. Neurobehavioral toxicology of pyrethroid insecticides in adult animals: A critical review. Neurotoxicol. Teratol. 2008, 30, 55–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paingankar, M.; Jain, M.; Deobagkar, D. Biodegradation of allethrin, a pyrethroid insecticide, by an Acidomonas sp. Biotechnol. Lett. 2005, 27, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- Na, H.Y.; Kim, Y.D.; Choi, Y.S.; Choi, Y.S.; Bae, C.H.; Song, Y.S. Allethrin and prallethrin stimulates MUC5AC expression through oxidative stress in human airway epithelial cells. Biochem. Biophysi. Res. Commun. 2018, 503, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Madhubabu, G.; Yenugu, S. Allethrin toxicity causes reproductive dysfunction in male rats. Environ. Toxicol. 2017, 32, 1701–1710. [Google Scholar] [CrossRef]
- Zhan, H.; Huang, Y.; Lin, Z.; Bhatt, P.; Chen, S. New insights into the microbial degradation and catalytic mechanism of synthetic pyrethroids. Environ. Res. 2020, 182, 109138. [Google Scholar] [CrossRef]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-degrading microorganisms and their potential for the bioremediation of contaminated soils: A review. Front. Microbiol. 2016, 7, 1463. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Alemany, Ò.; Eljarrat, E. Introduction to Pyrethroid Insecticides: Chemical Structures, Properties, Mode of Action and Use. In The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to pyrethroids. Toxicol. Rev. 2005, 24, 93–106. [Google Scholar] [CrossRef]
- Possetto, D.; Natera, J.; Sancho, M.I.; García, N.A.; Massad, W.A. Bioallethrin degradation by photo-fenton process in acetonitrile/water and aqueous β-cyclodextrin solutions. J. Photochem. Photobiol. Chem. 2018, 365, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Arif, A.; Salam, S.; Mahmood, R. Bioallethrin-induced generation of reactive species and oxidative damage in isolated human erythrocytes. Toxicol. In Vitro 2020, 65, 104810. [Google Scholar] [CrossRef]
- Hossain, M.M.; Suzuki, T.; Sato, I.; Takewaki, T.; Suzuki, K.; Kobayashi, H. Neuromechanical effects of pyrethroids, allethrin, cyhalothrin and deltamethrin on the cholinergic processes in rat brain. Life Sci. 2005, 77, 795–807. [Google Scholar] [CrossRef]
- Ramos-Chavez, L.A.; Sordo, M.; Calderon-Aranda, E.; Castañeda-Saucedo, E.; Ostrosky-Wegman, P.; Moreno-Godinez, M.E. A permethrin/allethrin mixture induces genotoxicity and cytotoxicity in human peripheral blood lymphocytes. J. Toxicol. Environ. Heal. Part A Curr. Issues 2015, 78, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Srivastava, P.K.; Al-Khedhairy, A.A.; Musarrat, J.; Shukla, Y. Allethrin-induced genotoxicity and oxidative stress in swiss albino mice. Mutat. Res. 2012, 747, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Chaitanya, R.K.; Golla, M.; Karnati, R. Allethrin toxicity on human corneal epithelial cells involves mitochondrial pathway mediated apoptosis. Toxicol. In Vitro 2013, 27, 2242–2248. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lin, Z.; Zhang, W.; Pang, S.; Bhatt, P.; Rene, E.R.; Kumar, A.J.; Chen, S. New insights into the microbial degradation of D-cyphenothrin in contaminated water/soil environments. Microorganims 2020, 8, 473. [Google Scholar] [CrossRef] [Green Version]
- Gangola, S.; Khati, P.; Sharma, A. Mycoremediation of imidaclopridin the presence of different soil amendments using Trichoderma longibrachiatum and Aspergillus oryzae isolated from pesticide contaminated agricultural fields of Uttarakhand. J. Biorem. Biodegredation 2015, 6, 2–5. [Google Scholar]
- Narendra, M.; Kavitha, G.; Helah Kiranmai, A.; Raghava Rao, N.; Varadacharyulu, N.C. Chronic exposure to pyrethroid-based allethrin and prallethrin mosquito repellents alters plasma biochemical profile. Chemosphere 2008, 73, 360–364. [Google Scholar] [CrossRef]
- Yang, J.; Feng, Y.; Zhan, H.; Liu, J.; Yang, F.; Zhang, K.; Zhang, L.; Chen, S. Characterization of a pyrethroid-degrading Pseudomonas fulva strain P31 and biochemical degradation pathway of D-phenothrin. Front. Microbiol. 2018, 9, 1003. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhan, H.; Chen, S. Insight into microbial applications for the biodegradation of pyrethroid insecticides. Front. Microbiol. 2019, 10, 1778. [Google Scholar] [CrossRef]
- Gangola, S.; Joshi, S.; Kumar, S.; Pandey, S.C. Comparative analysis of fungal and bacterial enzymes in biodegradation of xenobiotic compounds. Smart Bioremediation Technol. Microb. Enzym. 2019, 10, 169–189. [Google Scholar]
- Bhatt, P.; Huang, Y.; Zhang, W.; Sharma, A.; Chen, S. Enhanced cypermethrin degradation kinetics and metabolic pathway in Bacillus thuringiensis strain SG4. Microorganims 2020, 8, 223. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Hu, Q.; Hu, M.; Luo, J.; Weng, Q.; Lai, K. Isolation and characterization of a fungus able to degrade pyrethroids and 3-phenoxybenzaldehyde. Bioresour. Technol. 2011, 102, 8110–8116. [Google Scholar] [CrossRef] [PubMed]
- Birolli, W.G.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Biodegradation of the pyrethroid pesticide esfenvalerate by marine-derived fungi. Mar. Biotechnol. 2016, 18, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Birolli, W.G.; Vacondio, B.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Enantioselective biodegradation of the pyrethroid (±)-lambda-cyhalothrin by marine-derived fungi. Chemosphere 2018, 197, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Brown, W.; de Melo Souza, P.L.; Murphy, C.D. Cyhalothrin biodegradation in Cunninghamella elegans. Environ. Sci. Pollut. Res. 2018, 26, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.C.; Liu, Y.H.; Wang, Z.Y.; Zhang, X.Y.; Li, H.; Liang, W.Q.; Luo, N.; Hu, J.M.; Lu, J.Q.; Luan, T.G.; et al. Molecular cloning, purification, and biochemical characterization of a novel pyrethroid-hydrolyzing esterase from Klebsiella sp. strain ZD112. J. Agric. Food Chem. 2006, 54, 836–842. [Google Scholar] [CrossRef]
- Chen, S.; Liu, C.; Peng, C.; Liu, H.; Hu, M.; Zhong, G. Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-trichloro-2-pyridinol by a new fungal strain Cladosporium cladosporioides Hu-01. PLoS ONE 2012, 7, e47205. [Google Scholar] [CrossRef] [Green Version]
- Birolli, W.G.; Arai, M.S.; Nitschke, M.; Porto, A.L.M. The pyrethroid (±)-lambda-cyhalothrin enantioselective biodegradation by a bacterial consortium. Pestic. Biochem. Physiol. 2019, 156, 129–137. [Google Scholar] [CrossRef]
- Oliveira, B.R.; Penetra, A.; Cardoso, V.V.; Benoliel, M.J.; Crespo, M.T.V.; Samson, R.A.; Pereira, V.J. Biodegradation of pesticides using fungi species found in the aquatic environment. Environ. Sci. Poll. Res. 2015, 22, 11781–11791. [Google Scholar] [CrossRef]
- Barh, A.; Kumari, B.; Sharma, S.; Annepu, S.K.; Kumar, A.; Kamal, S.; Sharma, V.P. Mushroom mycoremediation: Kinetics and mechanism. Smart Bioremediation Technol. Microb. Enzym. 2019, 1, 1–22. [Google Scholar]
- Zhao, J.; Jia, D.; Chi, Y.; Yao, K. Co-metabolic enzymes and pathways of 3-phenoxybenzoic acid degradation by Aspergillus oryzae M-4. Ecotoxicol. Environ. Saf. 2020, 189, 109953. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhatt, K.; Huang, Y.; Ziqiu, L.; Chen, S. Esterase is a powerful tool for the biodegradation of pyrethroid insecticides. Chemosphere 2020, 244, 125507. [Google Scholar] [CrossRef] [PubMed]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Bioaugmentation as a strategy for the remediation of pesticide-polluted soil: A review. Chemosphere 2017, 172, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Huang, Y.; Rene, E.R.; Kumar, A.J.; Chen, S. Mechanism of allethrin biodegradation by a newly isolated Sphingomonas trueperi strain CW3 from wastewater sludge. Bioresour. Technol. 2020, 305, 123074. [Google Scholar] [CrossRef]
- Chen, S.; Luo, J.; Hu, M.; Geng, P.; Zhang, Y. Microbial detoxification of bifenthrin by a novel yeast and its potential for contaminated soils treatment. PLoS ONE 2012, 7, e30862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, H.; Wang, H.; Liao, L.; Feng, Y.; Fan, X.; Zhang, L.; Chen, S. Kinetics and novel degradation pathway of permethrin in Acinetobacter baumannii ZH-14. Front. Microbiol. 2018, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Gangola, S.; Sharma, A.; Bhatt, P.; Khati, P.; Chaudhary, P. Presence of esterase and laccase in Bacillus subtilis facilitates biodegradation and detoxification of cypermethrin. Sci. Rep. 2018, 8, 12755. [Google Scholar] [CrossRef]
- Chen, S.; Yang, L.; Hu, M.; Liu, J. Biodegradation of fenvalerate and 3-phenoxybenzoic acid by a novel Stenotrophomonas sp. strain ZS-S-01 and its use in bioremediation of contaminated soils. Appl. Microbiol. Biotechnol. 2011, 90, 755–767. [Google Scholar] [CrossRef]
- Chen, S.; Lai, K.; Li, Y.; Hu, M.; Zhang, Y.; Zeng, Y. Biodegradation of deltamethrin and its hydrolysis product 3-phenoxybenzaldehyde by a newly isolated Streptomyces aureus strain HP-S-01. Appl. Microbiol. Biotechnol. 2011, 90, 1471–1483. [Google Scholar] [CrossRef]
- Chen, S.; Geng, P.; Xiao, Y.; Hu, M. Bioremediation of β-cypermethrin and 3-phenoxybenzaldehyde contaminated soils using Streptomyces aureus HP-S-01. Appl. Microbiol. Biotechnol. 2012, 94, 505–515. [Google Scholar] [CrossRef]
- Chen, S.; Hu, M.; Liu, J.; Zhong, G.; Yang, L.; Rizwan-ul-Haq, M.; Han, H. Biodegradation of beta-cypermethrin and 3-phenoxybenzoic acid by a novel Ochrobactrum lupini DG-S-01. J. Hazard. Mater. 2011, 187, 433–440. [Google Scholar] [CrossRef]
- Chen, S.; Chang, C.; Deng, Y.; An, S.; Dong, Y.H.; Zhou, J.; Hu, M.; Zhong, G.; Zhang, L.H. Fenpropathrin biodegradation pathway in Bacillus sp. DG-02 and its potential for bioremediation of pyrethroid-contaminated soils. J. Agric. Food Chem. 2014, 62, 2147–2157. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Deng, Y.; Chang, C.; Lee, J.; Cheng, Y.; Cui, Z.; Zhou, J.; He, F.; Hu, M.; Zhang, L.H. Pathway and kinetics of cyhalothrin biodegradation by Bacillus thuringiensis strain ZS-19. Sci. Rep. 2015, 5, 8784. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T. Simultaneous determination of 18 pyrethroids in indoor air by gas chromatography/mass spectrometry. J. Chromatogr. A 2009, 1216, 5069–5076. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Xie, W.; Chen, S.; Zhang, N.; Zou, Y.; Dong, X.; Rashid, M.; Xiao, Y.; Hu, M.; Zhong, G. Separation of cis- and trans-cypermethrin by reversed-phase high-performance liquid chromatography. J. Chromatogr. Sci. 2015, 53, 612–618. [Google Scholar] [CrossRef] [Green Version]
- Pankaj; Sharma, A.; Gangola, S.; Khati, P.; Kumar, G.; Srivastava, A. Novel pathway of cypermethrin biodegradation in a Bacillus sp. strain SG2 isolated from cypermethrin-contaminated agriculture field. 3 Biotech 2016, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Dong, Y.H.; Chang, C.; Deng, Y.; Zhang, X.F.; Zhong, G.; Song, H.; Hu, M.; Zhang, L.H. Characterization of a novel cyfluthrin-degrading bacterial strain Brevibacterium aureum and its biochemical degradation pathway. Bioresour. Technol. 2013, 132, 16–23. [Google Scholar] [CrossRef]
- Cycoń, M.; Zmijowska, A.; Piotrowska-Seget, Z. Enhancement of deltamethrin degradation by soil bioaugmentation with two different strains of Serratia marcescens. Int. Environ. Sci. Technol. 2014, 11, 1305–1316. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.; Feng, Y.; Fan, X.; Chen, S. Recent advances in glyphosate biodegradation. Appl. Microbiol. Biotechnol. 2018, 10, 5033–5043. [Google Scholar] [CrossRef]
- Huang, Y.; Zhan, H.; Bhatt, P.; Chen, S. Paraquat degradation from contaminated environments: Current achievements and perspectives. Front. Microbiol. 2019, 10, 1754. [Google Scholar] [CrossRef] [Green Version]
- Tang, A.X.; Liu, H.; Liu, Y.Y.; Li, Q.Y.; Qing, Y.M. Purification and characterization of a novel β-cypermethrin-degrading aminopeptidase from Pseudomonas aeruginosa GF31. J. Agric. Food Chem. 2017, 65, 9412–9418. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, Y.; Zhan, H.; Bhatt, P.; Chen, S. An overview of strobilurin fungicide degradation: Current status and future perspective. Front. Microbiol. 2020, 11, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Lin, Z.; Pang, S.; Bhatt, P.; Chen, S. Insights into the biodegradation of lindane (γ-hexachlorocyclohexane) using a microbial system. Front. Microbiol. 2020, 11, 522. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Zhang, W.; Pang, S.; Huang, Y.; Mishra, S.; Bhatt, P.; Chen, S. Current approaches to and future perspectives on methomyl degradation in contaminated soil/water environments. Molecules 2020, 25, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, K.; Deng, W.; Zhu, Y.; Yao, K.; Li, J.; Liu, A.; Ao, X.; Zou, L.; Zhou, K.; He, L.; et al. Simultaneous degradation of β-cypermethrin and 3-phenoxybenzoic acid by Eurotium cristatum ET1, a novel“golden flower fungus” strain isolated from Fu Brick Tea. Microbiol. Open 2018, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Deng, W.Q.; Lin, D.R.; Yao, K.; Yuan, H.Y.; Wang, Z.L.; Li, J.L.; Zou, L.K.; Han, X.F.; Zhou, K.; He, L.; et al. Characterization of a novel beta-cypermethrin-degrading Aspergillus niger YAT strain and the biochemical degradation pathway of beta-cypermethrin. Appl. Microbiol. Biotechnol. 2015, 99, 8187–8198. [Google Scholar] [CrossRef]
- Yu, Y.L.; Fang, H.; Wang, X.; Wu, X.M.; Shan, M. Characteristics of fungal a fungal strain capable of degrading chlorpyrifos and its use in detoxification of the insecticide on vegetables. Biodegradation 2011, 17, 487–494. [Google Scholar] [CrossRef]
- Chen, S.; Lin, Q.; Xiao, Y.; Deng, Y.; Chang, C.; Zhong, G.; Hu, M.; Zhang, L.H. Monooxygenase, a novel beta-cypermethrin degrading enzyme from Streptomyces sp. PLoS ONE 2013, 8, e75450. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hu, C.; Zhang, R.; Sun, A.; Li, D.; Shi, X. Mechanism study of cyfluthirn biodegradation by Photobacterium ganghwense with comparative metabolomics. Appl. Microbiol. Biotechnol. 2018, 103, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, S.H.; Gao, Y.Q.; Hu, W.; Hu, M.Y.; Zhong, G.H. Isolation of a novel beta-cypermethrin degrading strain Bacillus subtilis BSF01 and its biodegradation pathway. Appl. Microbiol. Biotechnol. 2015, 99, 2849–2859. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luo, J.J.; Hu, M.Y.; Lai, K.P.; Geng, P.; Huang, H.S. Enhancement of cypermethrin degradation by a coculture of Bacillus cereus ZH-3 and Streptomyces aureus HP-S-01. Bioresour. Technol. 2012, 110, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, S.H.; Yan, Y.C. Isomerization and biodegradation of beta-cypermethrin by Pseudomonas aeruginosa CH7 with biosurfactant production. Bioresour. Technol. 2011, 102, 7139–7146. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Lin, Z.; Zhang, W.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the microbial degradation and biochemical mechanisms of neonicotinoids. Front. Microbiol. 2020, 11, 868. [Google Scholar]
- Guo, P.; Wang, B.; Hang, B.J.; Li, L.; Ali, S.W.; He, J.; Li, S. Pyrethroid-degrading Sphingobium sp. JZ-2 and the purification and characterization of a novel pyrethroid hydrolase. Int. Biodeterior. Biodegradation 2009, 63, 1107–1112. [Google Scholar] [CrossRef]
- Wang, B.; Guo, P.; Hang, B.; Li, L.; He, J.; Li, S.P. Cloning of a novel pyrethroid-hydrolyzing carboxylesterase gene from Sphingobium sp. strain JZ-1. Appl. Environ. Microbiol. 2009, 75, 5496–5500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Chen, S.; Hu, M.; Rizwan-ul-Haq, M.; Yang, L.; Li, H. Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int. J. Environ. Sci. Technol. 2011, 8, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Li, K.; Song, J.; Shi, Y.; Yan, Y. Molecular cloning, purification and biochemical characterization of a novel pyrethroid-hydrolyzing carboxylesterase gene from Ochrobactrum anthropi YZ-1. J. Hazard. Mater. 2012, 221–222, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Jilani, S.; Khan, M.A. Biodegradation of cypermethrin by Pseudomonas in a batch activated sludge process. Int. J. Environ. Sci. Technol. 2006, 3, 371–380. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | X1 | X2 | X3 | Allethrin Degradation (%) |
|---|---|---|---|---|
| 1 | +1 | +1 | −1 | 76.0 ± 0.88 d |
| 2 | 0 | 0 | 0 | 93.0 ± 0.57 g |
| 3 | 0 | 0 | −1.68 | 75.0 ± 1.10 d |
| 4 | 0 | 0 | 0 | 93.0 ± 1.70 g |
| 5 | +1 | −1 | −1 | 69.0 ± 0.57 b |
| 6 | −1 | −1 | −1 | 64.0 ± 0.57 a |
| 7 | 0 | 0 | 0 | 94.5 ± 0.28 ghi |
| 8 | 0 | 0 | 0 | 95.5 ± 0.28 hi |
| 9 | −1 | +1 | −1 | 73.0 ± 0.00 c |
| 10 | 0 | 0 | 0 | 95.6 ± 0.57 i |
| 11 | −1.68 | 0 | 0 | 76.0 ± 0.57 d |
| 12 | 0 | 0 | 0 | 93.5 ± 0.57 gh |
| 13 | 0 | 0 | 0 | 93.0 ± 0.57 g |
| 14 | 0 | +1.68 | 0 | 75.0 ± 0.57 d |
| 15 | −1 | +1 | +1 | 79.0 ± 0.57 e |
| 16 | 0 | 0 | 0 | 94.3 ± 0.03 ghi |
| 17 | −1 | −1 | +1 | 85.0 ± 0.00 f |
| 18 | 0 | 0 | +1.68 | 85.2 ± 0.11 f |
| 19 | +1 | +1 | +1 | 71.0 ± 0.60 c |
| 20 | 0 | −1.68 | 0 | 75.0 ± 0.00 d |
| 21 | +1 | −1 | +1 | 85.0 ± 0.00 f |
| 22 | 0 | 0 | 0 | 95.3 ± 0.57 hi |
| 23 | +1.68 | 0 | 0 | 75.0 ± 0.57 d |
| 24 | 0 | 0 | 0 | 94.5 ± 0.28 ghi |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatt, P.; Zhang, W.; Lin, Z.; Pang, S.; Huang, Y.; Chen, S. Biodegradation of Allethrin by a Novel Fungus Fusarium proliferatum Strain CF2, Isolated from Contaminated Soils. Microorganisms 2020, 8, 593. https://doi.org/10.3390/microorganisms8040593
Bhatt P, Zhang W, Lin Z, Pang S, Huang Y, Chen S. Biodegradation of Allethrin by a Novel Fungus Fusarium proliferatum Strain CF2, Isolated from Contaminated Soils. Microorganisms. 2020; 8(4):593. https://doi.org/10.3390/microorganisms8040593
Chicago/Turabian StyleBhatt, Pankaj, Wenping Zhang, Ziqiu Lin, Shimei Pang, Yaohua Huang, and Shaohua Chen. 2020. "Biodegradation of Allethrin by a Novel Fungus Fusarium proliferatum Strain CF2, Isolated from Contaminated Soils" Microorganisms 8, no. 4: 593. https://doi.org/10.3390/microorganisms8040593