Resistome, Mobilome and Virulome Analysis of Shewanella algae and Vibrio spp. Strains Isolated in Italian Aquaculture Centers




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Strain Identification, Phenotypical and Biochemical Analysis
2.2. Antimicrobial Susceptibility Testing
2.3. Genomic DNA Extraction and Genome Sequencing
2.4. Genome Annotation
3. Results
3.1. Bacterial Strain Characterization, Phenotypical and Biochemical Results
3.2. Antimicrobial Susceptibility Testing
3.3. Genome Features
3.4. Resistome Analysis
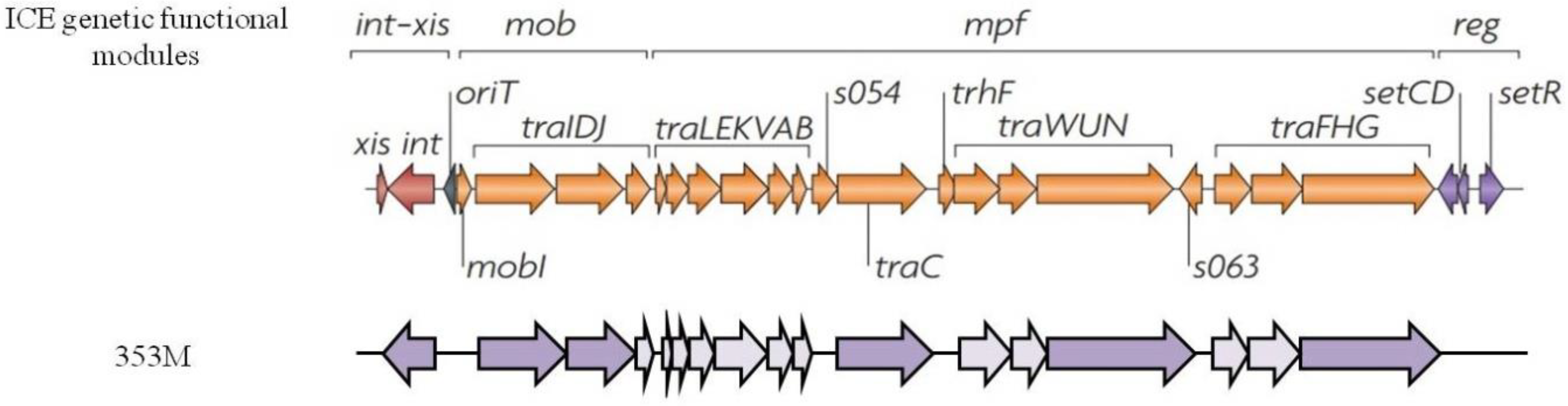
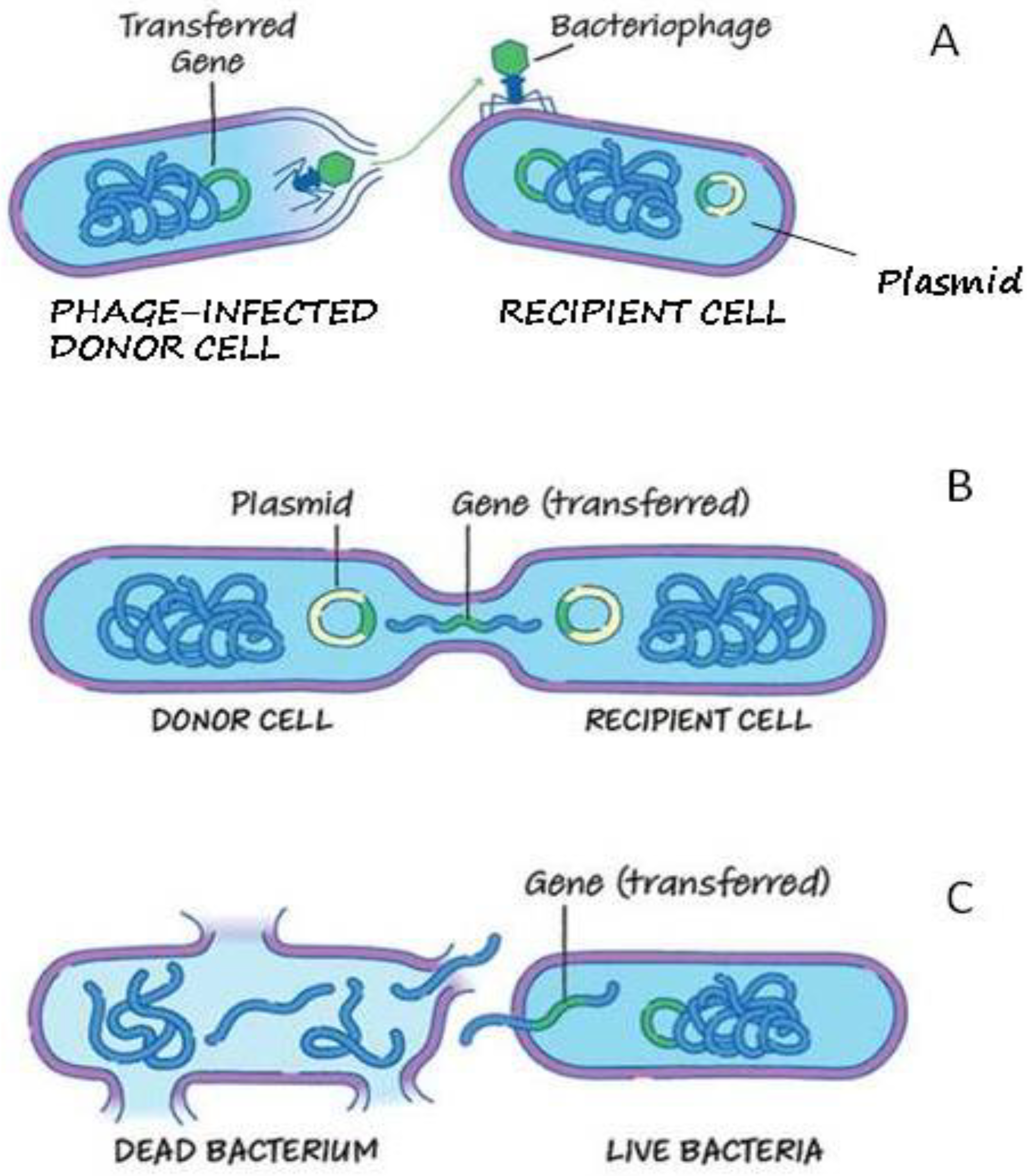
3.5. Mobilome Analysis
3.6. Virulome Analysis
3.7. Bacteriocins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jacob-Kokura, S.; Chan, C.Y.; Kaplan, L. Bacteremia and empyema caused by Shewanella algae in a trauma patient. Ann. Pharmacother. 2014, 48, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The genus Shewanella: From the briny depths below to human pathogen. Crit. Rev. Microbiol. 2014, 40, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Gallego, I.; Chaves, F.; Orellana, M.A. Epidemiological and clinical characteristics of Shewanella spp. infections in a tertiary hospital in Madrid. Infect. Dis. (Lond.) 2016, 48, 760–762. [Google Scholar] [CrossRef]
- Jousset, A.B.; Dabos, L.; Bonnin, R.A.; Girlich, D.; Potron, A.; Cabanel, N.; Dortet, L.; Glaser, P.; Naas, T. CTX-M-15-producing Shewanella species clinical isolate expressing OXA-535, a chromosome-encoded OXA-48 variant, putative progenitor of the plasmid-encoded OXA-436. Antimicrob. Agents Chemother. 2017, 62, e018791-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousfi, K.; Bekal, S.; Usongo, V.; Touati, A. Current trends of human infections and antibiotic resistance of the genus Shewanella. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Satomi, M. The Family Shewanellaceae. In The Prokaryotes, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Poirel, L.; Héritier, C.; Nordmann, P. Chromosome-encoded Ambler class D-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob. Agents Chemother. 2004, 48, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Rodriguez-Martinez, J.M.; Mammeri, H.; Liard, A.; Nordmann, P. Origin of plasmid-mediated quinolone resistance determinant QnrA. Antimicrob. Agents Chemother. 2005, 49, 3523–3525. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.M.; Kang, C.I.; Lee, C.S.; Kim, H.B.; Kim, E.C.; Kim, N.J.; Oh, M.D.; Choe, K.W. Treatment failure due to emergence of resistance to carbapenem during therapy for Shewanella algae bacteremia. J. Clin. Microbiol. 2006, 44, 1172–1174. [Google Scholar] [CrossRef] [Green Version]
- Lascols, C.; Podglajen, I.; Verdet, C.; Gauthier, V.; Gutmann, L.; Soussy, C.J.; Collatz, E.; Cambau, E. A plasmid-borne Shewanella algae gene, qnrA3, and its possible transfer In Vivo between Kluyvera ascorbata and Klebsiella pneumonia. J. Bacteriol. 2008, 190, 5217–5223. [Google Scholar] [CrossRef] [Green Version]
- Potron, A.; Poirel, L.; Nordmann, P. Origin of OXA-181, an emerging carbapenem-hydrolyzing oxacillinase, as a chromosomal gene in Shewanella xiamenensis. Antimicrob. Agents Chemother. 2011, 55, 4405–4407. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, T.; Olaitan, A.O.; Rolain, J.M. Whole genome sequence to decipher the resistome of Shewanella algae, a multidrug-resistant bacterium responsible for pneumonia, Marseille, France. Expert Rev. Anti Infect. Ther. 2016, 14, 269–275. [Google Scholar] [CrossRef]
- Ceccarelli, D.; van Essen-Zandbergen, A.; Veldman, K.T.; Tafro, N.; Haenen, O.; Mevius, D.J. Chromosome-based blaOXA-48-like variants in Shewanella species isolates from food-producing animals, fish, and the aquatic environment. Antimicrob. Agents Chemother. 2017, 61, e010131-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.H.; Tang, Y.Y.; Cheng, J.F.; Wu, Z.Y.; Mao, Y.C.; Liu, P.Y. Genome analysis of multidrug-resistant Shewanella algae isolated from human soft tissue sample. Front. Pharmacol. 2018, 9, 419. [Google Scholar] [CrossRef]
- Holt, H.M.; Søgaard, P.; Gahrn-Hansen, B. Ear infections with Shewanella alga: A bacteriologic, clinical and epidemiologic study of 67 cases. Clin. Microbiol. Infect. 1997, 3, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telke, A.A.; Rolain, J.M. Functional genomics to discover antibiotic resistance genes: The paradigm of resistance to colistin mediated by ethanolamine phosphotransferase in Shewanella algae MARS 14. Int. J. Antimicrob. Agents 2015, 46, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Jacobs-Slifka, K.M.; Newton, A.E.; Mahon, B.E. Vibrio alginolyticus infections in the USA, 1988–2012. Epidemiol. Infect. 2017, 145, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, H.A.; El Bayomi, R.M.; Hussein, M.A.; Khedr, M.H.E.; Abo Remela, E.M.; El-Ashram, A.M.M. Molecular characterization, antibiotic resistance pattern and biofilm formation of Vibrio parahaemolyticus and V. cholerae isolated from crustaceans and humans. Int. J. Food Microbiol. 2018, 274, 31–37. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-cholera vibrios: The microbial barometer of climate change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, D.; Salvia, A.M.; Sami, J.; Cappuccinelli, P.; Colombo, M.M. New cluster of plasmid-located class 1 integrons in Vibrio cholerae O1 and a dfrA15 cassette-containing integron in Vibrio parahaemolyticus isolated in Angola. Antimicrob. Agents Chemother. 2006, 50, 2493–2499. [Google Scholar] [CrossRef] [Green Version]
- Holm, K.O.; Bækkedal, C.; Söderberg, J.J.; Haugen, P. Complete genome sequences of seven Vibrio anguillarum strains as derived from PacBio sequencing. Genome Biol. Evol. 2018, 10, 1127–1131. [Google Scholar] [CrossRef]
- Bier, N.; Schwartz, K.; Guerra, B.; Strauch, E. Survey on antimicrobial resistance patterns in Vibrio vulnificus and Vibrio cholerae non-O1/non-O139 in Germany reveals carbapenemase-producing Vibrio cholerae in coastal waters. Front. Microbiol. 2015, 6, 1179. [Google Scholar] [CrossRef] [PubMed]
- Aberkane, S.; Compain, F.; Barraud, O.; Ouédraogo, A.S.; Bouzinbi, N.; Vittecoq, M.; Jean-Pierre, H.; Decré, D. Non-O1/non-O139 Vibrio cholerae avian isolate from France cocarrying the blaVIM-1 and blaVIM-4 genes. Antimicrob. Agents Chemother. 2015, 59, 6594–6596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, G.; Pazhani, G.P.; Sarkar, A.; Rajendran, K.; Mukhopadhyay, A.K.; Bhattacharya, M.K.; Ghosh, A.; Ramamurthy, T. Carbapenem resistance in clonally distinct clinical strains of Vibrio fluvialis isolated from diarrheal samples. Emerg. Infect. Dis. 2016, 22, 1754–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangat, C.S.; Boyd, D.; Janecko, N.; Martz, S.L.; Desruisseau, A.; Carpenter, M.; Reid-Smith, R.J.; Mulvey, M.R. Characterization of VCC-1, a novel Ambler class A carbapenemase from Vibrio cholerae isolated from imported retail shrimp sold in Canada. Antimicrob. Agents Chemother. 2016, 60, 1819–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, L.H.; Ab Mutalib, N.S.; Law, J.W.; Wong, S.H.; Letchumanan, V. Discovery on antibiotic resistance patterns of Vibrio parahaemolyticus in Selangor reveals carbapenemase producing Vibrio parahaemolyticus in marine and freshwater fish. Front. Microbiol. 2018, 9, 2513. [Google Scholar] [CrossRef] [PubMed]
- Di, D.Y.W.; Shin, H.; Han, D.; Unno, T.; Hur, H.G. High genetic diversity of Vibrio parahaemolyticus isolated from tidal water and mud of southern coast of South Korea. FEMS Microbiol. Ecol. 2019, 95, fiz022. [Google Scholar] [CrossRef]
- Rahman, T.; Yarnall, B.; Doyle, D.A. Efflux drug transporters at the forefront of antimicrobial resistance. Eur. Biophys. J. 2017, 46, 647–653. [Google Scholar] [CrossRef]
- Lubelski, J.; Konings, W.N.; Driessen, A.J. Distribution and physiology of ABC-type transporters contributing to multidrug resistance in bacteria. Microbiol. Mol. Biol. Rev. 2007, 71, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, T.; Tsuchiya, T. Multidrug efflux transporters in the MATE family. Biochimica Biophysica Acta 2009, 1794, 763–768. [Google Scholar] [CrossRef]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.J.; Saier, M.H., Jr. SMR-type multidrug resistance pumps. Curr. Opin. Drug Discov. Dev. 2001, 4, 237–245. [Google Scholar]
- Nikaido, H. Structure and mechanism of RND-type multidrug efflux pumps. Adv. Enzymol. Relat. Areas Mol. Biol. 2011, 77, 1–60. [Google Scholar]
- Groh, J.L.; Luo, Q.; Ballard, J.D.; Krumholz, L.R. Genes that inhance the ecological fitness of Shewanella oneidensis MR-1 in sediments reveal the value of antibiotic resistance. Appl. Environ. Microbiol. 2007, 73, 492–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pembroke, J.T.; Piterina, A.V.C. A novel ICE in the genome of Shewanella putrefaciens W3-18-1: Comparison with the SXT/R391 ICE-like elements. FEMS Microbiol. Lett. 2006, 264, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Carraro, N.; Burrus, V. Biology of Three ICE Families: SXT/R391, ICEBs1, and ICESt1/ICESt3. Microbiol. Spectr. 2014, 2, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, Y.; Li, Z.; Liu, Z.; Li, X.; Diao, B.; Kan, B.; Wang, D. Distribution and genetic characteristics of SXT/R391 Integrative Conjugative Elements in Shewanella spp. from China. Front. Microbiol. 2018, 9, 920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamruzzaman, M.; Nischibuchi, M. Detection and characterization of a functional insertion sequence, ISVpa2, in Vibrio parahaemolyticus. Gene 2008, 409, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Yao, L.; Li, F.; Tan, Z.; Zhai, Y.; Wang, L. Chacterization of antimicrobial resistance of Vibrio parahaemolyticus from cultured sea cucumbers (Apostichopus japonicas). Lett. Appl. Microbiol. 2014, 59, 147–154. [Google Scholar] [CrossRef]
- Almuzara, M.; Montaña, S.; Lazzaro, T.; Uong, S.; Parmeciano Di Noto, G.; Traglia, G.; Bakai, R.; Centrón, D.; Iriarte, A.; Quiroga, C.; et al. Genetic analysis of a PER-2-producing Shewanella sp. strain harbouring a variety of mobile genetic elements and antibiotic resistance determinants. J. Glob. Antimicrob. Resist. 2017, 11, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Rossen, J.W.A.; Friedrich, A.W.; Moran-Gilad, J.; ESCMID Study Group for Genomic and Molecular Diagnostics (ESGMD). Practical issues in implementing whole-genome-sequencing in routine diagnostic microbiology. Clin. Microbiol. Infect. 2018, 24, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Köser, C.U.; Ellington, M.J.; Peacock, S.J. Whole-genome sequencing to control antimicrobial resistance. Trends Genet. 2014, 30, 401–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labella, A.; Gennari, M.; Ghidini, V.; Trento, I.; Manfrin, A.; Borrego, J.J.; Lleo, M.M. High incidence of antibiotic multi-resistant bacteria in coastal areas dedicated to fish farming. Mar. Pollut. Bull. 2013, 70, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Pires, J.; Novais, A.; Peixe, L. Blue-carba, an easy biochemical test for detection of diverse carbapenemase producers directly from bacterial cultures. J. Clin. Microbiol. 2013, 51, 4281–4283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder web-services for easy identification of acquired antibiotic resistance and E. coli virulence genes in bacteriophage and prophage nucleotide sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Moura, A.; Soares, M.; Pereira, C.; Leitão, N.; Henriques, I.; Correia, A. INTEGRALL: A database and search engine for integrons, integrases and gene cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D323–D326. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J. ICEberg 2.0: An updated database of bacterial integrative and conjugative elements. Nucleic Acids Res. 2019, 47, D660–D665. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozov, R.; Brown Kav, A.; Bogumil, D.; Shterzer, N.; Halperin, E.; Mizrahi, I.; Shamir, R. Recycler: An algorithm for detecting plasmids from de novo assembly graphs. Bioinformatics 2017, 33, 475–482. [Google Scholar]
- Héritier, C.; Poirel, L.; Nordmann, P. Genetic and biochemical characterization of a chromosome-encoded carbapenem-hydrolyzing Ambler class D beta-lactamase from Shewanella algae. Antimicrob. Agents Chemother. 2004, 48, 1670–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Anes, J.; McCusker, M.P.; Fanning, S.; Martins, M. The ins and outs of RND efflux pumps in Escherichia coli. Front. Microbiol. 2015, 6, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, A.F.; Waldor, M.K. Integrative and conjugative elements: Mosaic mobile genetic elements inabling dynamic lateral gene flow. Nat. Rev. Microbiol. 2010, 8, 552–563. [Google Scholar] [CrossRef]
- Teuber, M. Veterinary use and antibiotic resistance. Curr. Opin. Microbiol. 2001, 4, 493–499. [Google Scholar] [CrossRef]
- Hexing, W.; Lingshuang, R.; Xin, Y.; Jing, H.; Yue, C.; Gengsheng, H.; Qingwu, J. Antibiotic residues in meat, milk and aquatic products in Shanghai and human exposure assessment. Food Control 2017, 80, 217–225. [Google Scholar]
- Sorum, H. Antimicrobial Drug Resistance in Fish Pathogens. In Antimicrobial Resistance in Bacteria of Animal Origin; Aarestrup, F.M., Ed.; ASM Press: Washington, DC, USA, 2006; pp. 213–238. [Google Scholar]
- OIE. Report of the Meeting of the OIE ad hoc Group on Antimicrobial Resistance; Annex 9; OIE: Paris, France, 2018. [Google Scholar]
- Regulation, H. The Council of the European Communities. Council Regulation (ECC) No 2377/90 Laying down a Community Procedure for the Establishment of Maximum Residue Limits of Veterinary Medicine Products in Foodstuffs of Animal Origins; Annex III. Off. J. Eur. Commun. 1990, 224, 1–8. [Google Scholar]
- Scarano, C.; Spanu, C.; Ziino, G.; Pedonese, F.; Dalmasso, A.; Spanu, V.; Virdis, S.; De Santis, E.P. Antibiotic resistance of Vibrio species isolated from Sparus aurata reared in Italian mariculture. New Microbiol. 2014, 37, 329–337. [Google Scholar] [PubMed]
- Ottaviani, D.; Bacchiocchi, I.; Masini, L.; Leoni, F.; Carraturo, A.; Giammarioli, M.; Sbaraglia, G. Antimicrobial susceptibility of potentially pathogenic halophilic vibrios isolated from seafood. Int. J. Antimicrob. Agents 2001, 18, 135–140. [Google Scholar] [CrossRef]
- Laganà, P.; Caruso, G.; Minutoli, E.; Zaccone, R.; Santi, D. Susceptibility to antibiotics of Vibrio spp. and Photobacterium damsela ssp. piscicida strains isolated from Italian aquaculture farms. New Microbiol. 2011, 34, 53–63. [Google Scholar] [PubMed]
- Ottaviani, D.; Leoni, F.; Talevi, G.; Masini, L.; Santarelli, S.; Rocchegiani, E.; Susini, F.; Montagna, C.; Monno, R.; D’Annibale, L.; et al. Extensive investigation of antimicrobial resistance in Vibrio parahaemolyticus from shellfish and clinical sources, Italy. Int. J. Antimicrob. Agents 2013, 42, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Scarano, C.; Piras, F.; Virdis, S.; Ziino, G.; Nuvoloni, R.; Dalmasso, A.; De Santis, E.P.L.; Spanu, C. Antibiotic resistance of Aeromonas spp. strains isolated from Sparus aurata reared in Italian mariculture farms. Int. J. Food Microbiol. 2018, 284, 91–97. [Google Scholar] [CrossRef]
- Poirel, L.; Cattoir, V.; Nordmann, P. Plasmid-mediated quinolone resistance; interactions between human, animal, and environmental ecologies. Front. Microbiol. 2012, 2, 3–24. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Poeta, P.; Hébraud, M.; Capelo, J.L.; Igrejas, G. Mechanisms of quinolone action and resistance: Where do we stand? J. Med. Microb. 2017, 66, 551–559. [Google Scholar] [CrossRef]
- Bado, I.; Papa-Ezdra, R.; Cordeiro, N.; Outeda, M.; Caiata, L.; García-Fulgueiras, V.; Seija, V.; Vignoli, R. Detection of qnrVC6, within a new genetic context, in an NDM-1-producing Citrobacter freundii clinical isolate from Uruguay. J. Glob. Antimicrob. Resist. 2018, 14, 95–98. [Google Scholar] [CrossRef]
- Liu, J.; Yang, L.; Chen, D.; Peters, B.M.; Li, L.; Li, B.; Xu, Z.; Shirtliff, M.E. Complete sequence of pBM413, a novel multidrug resistance megaplasmid carrying qnrVC6 and blaIMP-45 from Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2018, 51, 145–150. [Google Scholar] [CrossRef]
- Torri, A.; Bertini, S.; Schiavone, P.; Congestrì, F.; Matteucci, M.; Sparacino, M.; Testa, G.; Pedna, M.F.; Sambri, V. Shewanella algae infection in Italy: Report of 3 years’ evaluation along the coast of the northern Adriatic Sea. New Microb. New Infect. 2018, 23, 39–43. [Google Scholar] [CrossRef]
- Miranda, C.D.; Kehrenberg, C.; Ulep, C.; Schwarz, S.; Roberts, M.C. Diversity of tetracycline resistance genes in bacteria from Chilean salmon farms. Antimicrob. Agents Chemother. 2003, 47, 883–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossman, T.H. Tetracycline antibiotics and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Biology Notes. Available online: https://thebiologynotes.com/carbapenem-resistant-gram-negative-bacteria/ (accessed on 15 January 2020).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Sampling Date | Source | 16S rRNA Gene Identification | Antimicrobial Resistance Profile |
|---|---|---|---|---|
| 353M | 10/08/2010 | Water Open sea | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 178CP | 13/06/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 146bCP | 13/06/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 144bCP | 13/06/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 219VB | 30/09/2010 | Water Veneto I station | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 82CP | 15/04/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, CS, SUL, FOS |
| 38LV | 22/07/2011 | Water Varano lake | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS, TET, TGC |
| 57CP | 15/04/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, CS, SUL, FOS, TET, TGC |
| 60CP | 15/04/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 83CP | 15/04/2011 | Water Veneto II station | Shewanella algae | CEF, FOX, IPM, AMX, CS, SUL, FOS |
| 28AD | 01/02/2007 | European seabass | Vibrio anguillarum | CEF, FOX, AMX, TIC, AMC, TCC, TEM, CS, SUL, AKN |
| VPE116 | 26/06/2007 | Water Caleri lagoon | Vibrio parahaemolyticus | AMX, TIC, TMN, KMN, AKN, GMI, CS, SUL |
| Antimicrobial | Pentapeptide Protein | Enzyme | Efflux System | Pump Family | Strain |
|---|---|---|---|---|---|
| Beta-lactams | OXA-55-like | All S. algae | |||
| AmpC | All S. algae, V. anguillarum 28AD | ||||
| MexAB-OprM | RND | All S. algae, V. parahaemolyticus VPE116 | |||
| Fluoroquinolones | QnrA3 | 353M, 219VB, 38LV, 57CP, 60CP | |||
| QnrA7 | 144bCP, 178CP, 146bCP, 82CP, 83CP | ||||
| QnrVC6 | V. anguillarum 28AD | ||||
| MdtK | MATE | All S. algae, V. anguillarum 28AD | |||
| EmrAB-TolC | MFS | All S. algae, V. parahaemolyticus VPE116 | |||
| MepAB | All S. algae, V. anguillarum 28AD | ||||
| NorM | MATE | V. anguillarum 28AD, V. parahaemolyticus VPE116 | |||
| Bmr3 | MFS | V. anguillarum 28AD | |||
| MdtH | MFS | V. anguillarum 28AD | |||
| MfpA | V. anguillarum 28AD | ||||
| Multiple substrates | AcrAB-TolC | RND | All S. algae, V. parahaemolyticus VPE116 | ||
| AcrEF-TolC | RND | All S. algae, V. parahaemolyticus VPE116 | |||
| MdtABC-TolC | RND | All strains | |||
| EmrD | MFS | All strains | |||
| EmrYK-TolC | MFS | All strains excepting for 353M, 219VB, 82CP, 38LV | |||
| MdtN | RND | All strains | |||
| DrrA | ABC | All strains excepting for V. anguillarum 28AD | |||
| Stp | MFS | All S. algae | |||
| Bcr | MFS | All S. algae | |||
| MdtG | MFS | V. parahaemolyticus VPE116 | |||
| MdlB | ABC | V. parahaemolyticus VPE116 | |||
| Tetracyclines | TetR | MFS | V. parahaemolyticus VPE116 | ||
| Tet34 | MFS | V. anguillarum 28AD, V. parahaemolyticus VPE116 | |||
| Tet35 | MFS | V. parahaemolyticus VPE116 | |||
| Macrolides | MacAB-TolC | ABC | All strains | ||
| MdtE-TolC | RND | V. anguillarum 28AD, V. parahaemolyticus VPE116 | |||
| Polymyxins | EptA | All strains | |||
| Chloramphenicol | MdtL | MFS | All strains | ||
| Cat | All strains excepting for V. anguillarum 28AD |
| Strain | Insertion Sequence (IS) Family | IS Family Subgroup | Integron/Integrase | Bacteriophage |
|---|---|---|---|---|
| 353M | IS630, ISL3, IS3 IS630 IS3 IS4 | ISSpu8 ISSlo2 ISVvu3 | IntI Class I integron | Prophage-1 incomplete |
| 219VB | IS200/IS605, IS630, ISL3 IS481 IS110, IS3 IS630 IS4 | ISSpu18 ISSpu8 ISVvu3 | Class I integron | Aeromonas phage phiO18P (NC_009542) |
| 144BCP | ISL3 IS481 IS630/ISSpu8 IS4 IS3, IS630, IS110 IS200/IS605 IS110, IS4 | ISSpu18 ISVvu3 ISSod ISSba | Class I integron | Escherichia phage D108 (NC_013594) Shewanella sp. phage 1/44 (NC_025463) Prophage-1 and Prophage-2 incomplete |
| 178CP | ISL3 IS481 IS630/ISSpu8 IS4 IS3, IS630, IS110 IS200/IS605 IS110, IS4 | ISSpu18 ISVvu3 ISSod ISSba | Class I integron | Escherichia phage D108 (NC_013594) Shewanella sp. phage 1/44 (NC_025463) Prophage-1 and Prophage-2 incomplete |
| 82CP | IS630, ISL3, IS481/ISSpu18, IS110, IS3 IS630 | ISSod16 | IntI Class I integron | Prophage-1 Aeromonas phage phiO18P (NC_009542) |
| 146BCP | ISL3 IS481 IS630/ISSpu8 IS4 IS3, IS630, IS110 IS200/IS605 IS110, IS4 | ISSpu18 ISVvu3 ISSod ISSba | Class I integron | Escherichia phage D108 (NC_013594) Shewanella sp. phage 1/44 (NC_025463) Prophage-1 and Prophage-2 incomplete |
| 83CP | IS200/IS605, ISL3, IS110 IS630 IS630 IS4 IS3 | ISSpu8 ISVvu3 ISSba6 ISSlo2 | IntI Class I integron | Prophage-1 Aeromonas phage phiO18P (NC_009542) |
| 38LV | ISL3 IS481 IS110 IS630 | ISSpu18 ISSod16 | IntI Class I integron | - |
| 57CP | IS200/IS605, ISL3, IS110 IS3 IS630 IS630 IS4 | ISSpu8 ISSod16 ISVvu3 ISSba6 | IntI Class I integron | Enterobacteria phage phi92 (NC_023693) incomplete |
| 60CP | IS200/IS605, ISL3, IS110 IS3 IS630 | ISSlo2 ISSod16 | Class I integron | 2 Unknown Prophages incomplete |
| 28AD | IS200/IS605, IS5/IS1182, ISAs1 IS30 | ISSpu13 ISVa6 | IntI Class I integron | 2 Unknown Prophages incomplete |
| VPE116 | IS5/IS1182 IS5 IS5 IS3 | ISSpu14 ISVpa3 ISVisp3 | IntI | Enterobacterial phage mEp213 (NC_019720) Vibrio phage VP882 (NC_009016) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zago, V.; Veschetti, L.; Patuzzo, C.; Malerba, G.; Lleo, M.M. Resistome, Mobilome and Virulome Analysis of Shewanella algae and Vibrio spp. Strains Isolated in Italian Aquaculture Centers. Microorganisms 2020, 8, 572. https://doi.org/10.3390/microorganisms8040572
Zago V, Veschetti L, Patuzzo C, Malerba G, Lleo MM. Resistome, Mobilome and Virulome Analysis of Shewanella algae and Vibrio spp. Strains Isolated in Italian Aquaculture Centers. Microorganisms. 2020; 8(4):572. https://doi.org/10.3390/microorganisms8040572
Chicago/Turabian StyleZago, Vanessa, Laura Veschetti, Cristina Patuzzo, Giovanni Malerba, and Maria M. Lleo. 2020. "Resistome, Mobilome and Virulome Analysis of Shewanella algae and Vibrio spp. Strains Isolated in Italian Aquaculture Centers" Microorganisms 8, no. 4: 572. https://doi.org/10.3390/microorganisms8040572