Unraveling the Fungal Community Associated with Leaf Spot on Crataegus sp.

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Leaves
2.2. Fungal Isolation and Purification
2.3. Phenotypic Identification
2.4. Inoculum Preparation
2.5. Plant Infection Assay
2.6. Molecular Identification and Phylogenetic Analysis
3. Results
3.1. Disease Symptoms
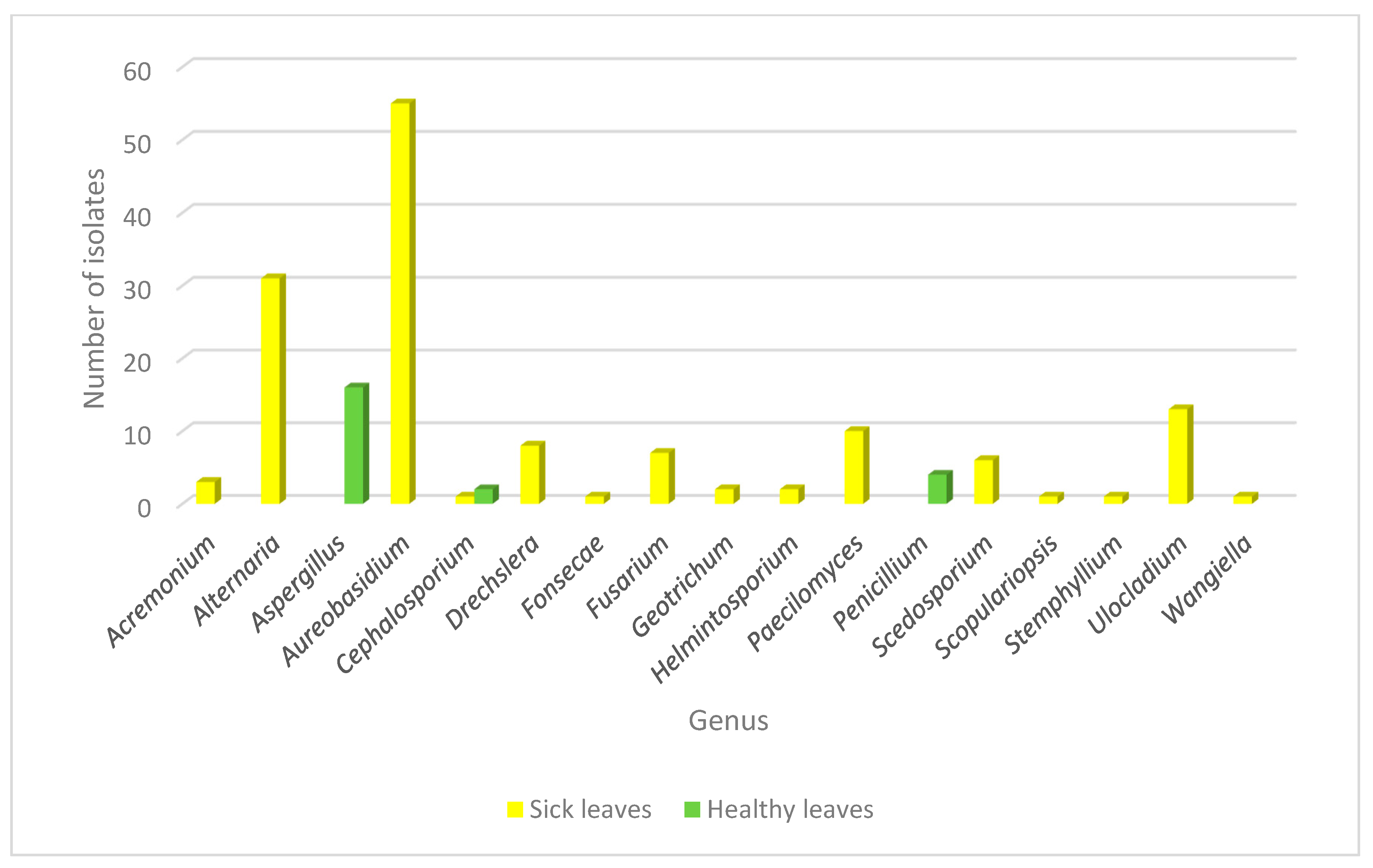
3.2. Fungal Community Associated to the Leaf Spot in Crataegus spp Plants
3.3. Pathogenicity Assay
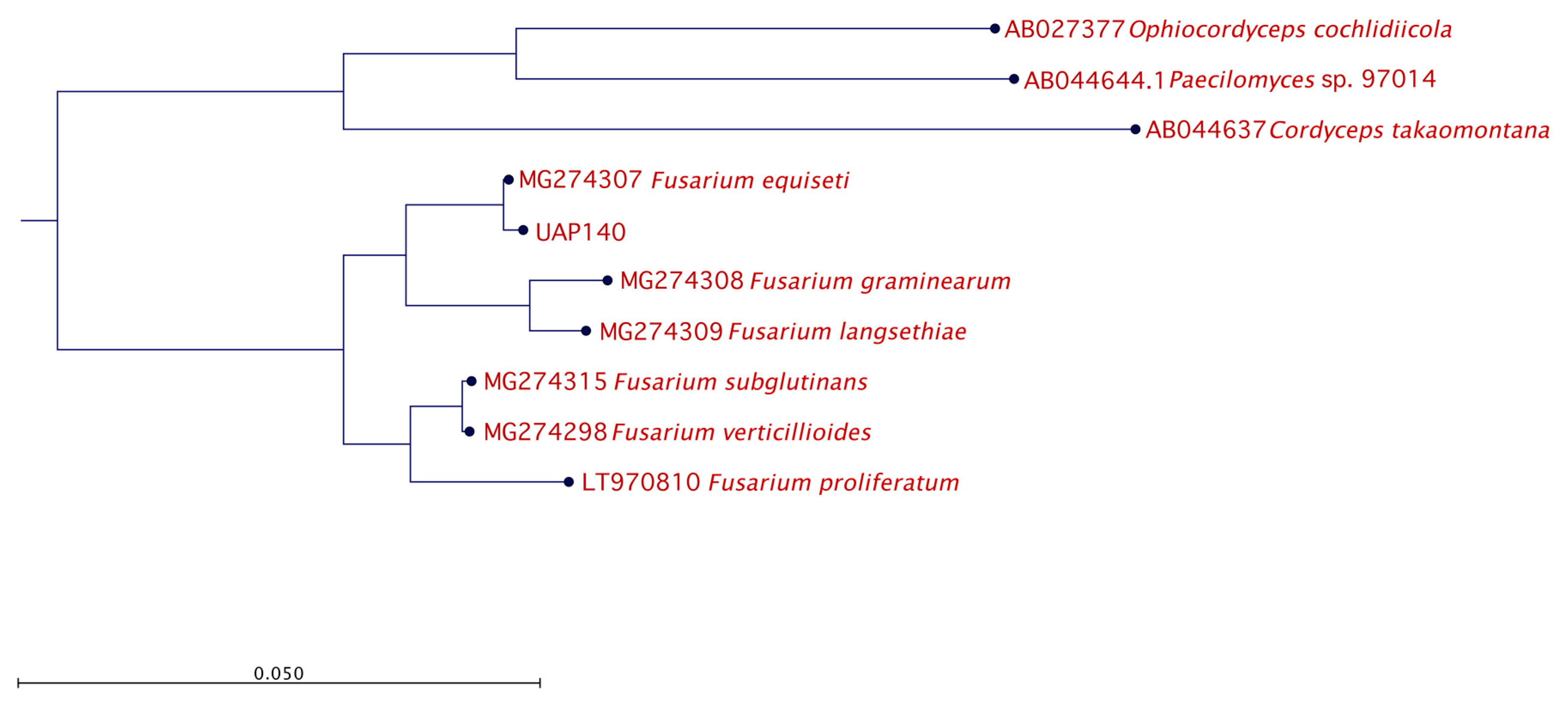
3.4. Molecular Identification
4. Discussion
4.1. Pathogens for Crataegus spp.
4.2. The Fungal Community Associated to Diseased Crataegus spp. Plants
4.3. The Context of the Disease
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chang, Q.; Zuo, Z.; Harrison, F.; Chow, M.S. Hawthorn. J. Clin. Pharmacol. 2002, 2, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Phipps, J.B.; O’Kennon, R.; Lance, R.W. Hawthorns and Medlars; Timber Press: Portland, OR, USA, 2003. [Google Scholar]
- Núñez-Colín, C.; Nieto-Angel, R.; Barrientos-Priego, A.F.; Sahagún-Castellanos, J.; Segura, S.; González-Andrés, F. Variability of three regional sources of germplasm of tejocote (Crataegus spp.) from central and southern Mexico. Genet. Resour. Crop. Evol. 2008, 55, 1159–1165. [Google Scholar] [CrossRef]
- Nieto-Angel, R.; Borys, M.W. Enfoques tecnológicos en la fruticultura. In Un tributo a Raúl Mosqueda, 1st ed.; Universidad Autónoma de Chapingo: Chapingo, México, 2008; Germoplasma y usos del tejocote en México. [Google Scholar]
- Khalil, R.; Abuharfeil, N.; Shabsoug, B. The effect of Crataegus aronica aqueous extract in rabbits fed with high cholesterol diet. Eur. J. Sci. Res. 2008, 22, 352–360. [Google Scholar]
- Pittler, M.H.; Schmidt, K.; Ernst, E. Hawthorn extract for treating chronic heart failure: Meta-analysis of randomized trials. Am. J. Med. 2003, 114, 665–674. [Google Scholar] [CrossRef]
- Güven, K.; Yücel, E.; Çetintaş, F. Antimicrobial activities of fruits of Crataegus and Pyrus species. Pharm. Biol. 2006, 44, 79–83. [Google Scholar] [CrossRef]
- Bahri-Sahloul, R.; Ben Fredj, R.; Boughalleb, N.; Shriaa, J.; Saguem, S.; Hilbert, J.L.; Harzallah-Skhiri, F. Phenolic composition and antioxidant and antimicrobial activities of extracts obtained from Crataegus azarolus L. var. aronia (Willd.) Batt. ovaries calli. J. Bot. 2014, 2014, 623651. [Google Scholar]
- Verma, S.K.; Jain, V.; Verma, D.; Khamesra, R. Crataegus Oxyacantha—A Cardioprotective herb. J. Herbal Med. Toxicol 2007, 1, 65–71. [Google Scholar]
- Bahri-Sahloul, R.; Ammar, S.; Fredj, R.B.; Saguem, S.; Grec, S.; Trotin, F. Polyphenol contents and antioxidant activities of extracts from flowers of two Crataegus azarolus L. varieties. Pakistan J. Biol. Sci. 2009, 12, 660–668. [Google Scholar]
- Nieto-Angel, R. Frutales Nativos. In Un recurso fitogenético de México, 1st ed.; Universidad Autónoma de Chapingo: Chapingo, México, 2007; Colección, conservación y caracterización del tejocote (Crataegus spp.); pp. 26–118. [Google Scholar]
- Nieto-Angel, R.; Borys, M.W. Relaciones fisiológicas y morfológicas de injertos de frutales sobre tejocote (Crataegus spp.) como portainjerto. Revista Chapingo Serie. Hortic. 1999, 5, 137–150. [Google Scholar] [CrossRef]
- Alvarado-Rosales, D.; Nieto-López, E.H.; Téliz-Ortiz, D.; Ayala-Escobar, V.; Silva-Rojas, H.V.; Nieto-Angel, R.; Leyva-Mir, S.G.; Jimenez-Nieto, A.; Méndez-Inocencio, C. First report of Gymnosporangium clavipes Cooke Peck affecting Crataegus mexicana var. Chapeado and C. gracilior in Mexico. Res. Plant Dis. 2015, 21, 250–253. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, W.A.; Lyon, H.H. Diseases of Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 2005. [Google Scholar]
- Prats, G. Microbiología clínica. Ed. Médica Panamericana 2006, 366. [Google Scholar]
- Salazar-Cerezo, S.; Martínez-Montiel, N.; Cruz-López, M.C.; Martínez-Contreras, R. Fungal diversity and community composition of culturable fungi in Stanhopea tigrina cast gibberellin producers. Front. Microbiol. 2018, 9, 612. [Google Scholar] [CrossRef] [PubMed]
- Koneman, E. Diagnóstico Microbiólogico: Texto y Atlas a color; Editorial Médica Panamericana: Buenos Aires, Argentina, 2001; p. 1432. [Google Scholar]
- Arenas, R. Micología Médica Ilustrada, 3rd ed.; Mc Graw-Hill: México, 2008. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bac. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munguía-Pérez, R.; Díaz-Cabrera, E.; Martínez-Montiel, N.; Muñoz-Rojas, J.; Martínez-Contreras, R. Fungal diversity in soil samples from a Mexican region with endemic dermatomycoses. Micol. Apl. Int. 2011, 23, 1. [Google Scholar]
- Rull, J.; Aluja, M.; Feder, J.; Berlocher, S. Distribution and host range of hawthorn-infesting Rhagoletis (Diptera: Tephritidae) in Mexico. Ann. Entomol. Soc. Am. 2006, 99, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Yun, H.Y.; Lee, K.J.; Kim, Y.H.; Lee, S.K. First Report of Gymnosporangium globosum Causing American Hawthorn Rust in Korea. Plant Pathol. J. 2008, 24, 84–86. [Google Scholar] [CrossRef] [Green Version]
- Udayashankar, A.C.; Chandra Nayaka, S.; Archana, B.; Anjana, G.; Niranjana, S.R.; Mortensen, S.N.; Lund, O.S.; Prakash, H.S. Specific PCR-based detection of Alternaria helianthi: The cause of blight and leaf spot in sunflower. Arch. Microbiol 2012, 194, 923–932. [Google Scholar] [CrossRef]
- Logrieco, A.; Bottalico, A.; Mul, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. Eur J. Plant. Pathol 2003, 109, 645–667. [Google Scholar] [CrossRef]
- Lucas, G.B.; Campbell, C.L.; Lucas, L.T. Introduction to Plant Diseases: Identification and Management, 2nd ed.; Van Nostrand Reinhold Company: New York, NY, USA, 1992. [Google Scholar]
- Daughtrey, M.L. Plagas y enfermedades de las plantas en macetas con flores; American Phytopathological Society, Ed.; Mundi-Prensa: Madrid, España, 2001. [Google Scholar]
- Luo, H.; Tao, Y.Q.; Fan, X.Y.; Oh, S.K.; Lu, H.X.; Denga, J.X. Identification and Characterization of Alternaria iridiaustralis Causing Leaf Spot on Iris ensata in China. Mycobiology 2018, 46, 168–171. [Google Scholar] [CrossRef] [Green Version]
- Wee, J.I.; Park, J.H.; Back, C.G.; You, Y.H.; Chang, T. First Report of Leaf Spot Caused by Alternaria tenuissima on Black Chokeberry (Aronia melanocarpa) in Korea. Mycobiology 2016, 44, 187–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.P.; Paul, N.C.; Lee, H.B.; Yu, S.H. First Record of Alternaria simsimi Causing Leaf Spot on Sesame (Sesamum indicum L.) in Korea. Mycobiology 2014, 42, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garganese, F.; Schena, L.; Siciliano, I.; Prigigallo, M.I.; Spadaro, D.; De Grassi, A. Characterization of Citrus-Associated Alternaria Species in Mediterranean Areas. PLoS ONE 2016, 11, e0163255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garganese, F.; Ippolito, A.; di Rienzo, V.; Lotti, C.; Montemurro, C.; Sanzani, S.M. A new high-resolution melting assay for genotyping Alternaria species causing citrus brown spot. J. Sci. Food Agric. 2018, 98, 4578–4583. [Google Scholar] [CrossRef] [PubMed]
- Rivera, V.; Rengifo, J.; Khan, M.; Geiser, D.M.; Mansfield, M.; Secor, G. First Report of a novel Fusarium species causing yellowing decline of sugar beet in Minnesota. Plant. Dis. 2008, 92, 1589. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.T.; Kirkpatrick, S.C.; Gordon, T.R. Fusarium wilt of strawberry caused by Fusarium oxysporum in California. Plant. Dis 2009, 93, 1077. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Rodríguez, R.; Guerrero, B.; González, M.; Pons, J.; Jiménez, J.; Ramírez, J.; Andrio, E.; Mendoza, M. Caracterización de especies de Fusarium asociadas a la pudrición de raíz de maíz en Guanajuato, México. Iberoamericana de Fitopatología 2010, 28, 124–134. [Google Scholar]
- Davis, M.R.; Subbarao, K.V.; Raid, N.R.; Kurtz, E.A. Plagas y enfermedades de la lechuga; American Phytopathological Society, Ed.; Mundi-Prensa: Madrid, España, 2002. [Google Scholar]
- Han, K.S.; Park, J.H.; Back, C.G.; Park, M.J. First report of Fusarium subglutinans causing leaf spot disease on Cymbidium orchids in Korea. Mycobiology 2015, 43, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Parkunan, V.; Ji, P. First Report of Leaf Spot on Onion Caused by Fusarium acuminatum in Georgia. Plant. Dis. 2013, 97, 1384. [Google Scholar] [CrossRef]
- Garibaldi, A.; Gilardi, G.; Ortu, G.; Gullino, M.L. First Report of Leaf Spot of Wild Rocket (Diplotaxis tenuifolia) Caused by Fusarium equiseti in Italy. Plant Dis. 2015, 99, 1183. [Google Scholar] [CrossRef]
- Kishore, R.; Tripathi, R.D.; Johri, J.K.; Shukla, D.S. Some new fungal diseases of opium poppy (Papaver somniferum). J. Plant Pathol. 1987, 3, 213–217. [Google Scholar]
- Hafeez, R.; Akhtar, N.; Shoaib, A.; Bashir, U.; Haider, M.S.; Awan, Z.A. First report of Geotrichum candidum from Pakistan causing postharvest sour rot in loquat (Eriobotrya Japonica). J. Anim. Plant Sci. 2015, 25, 1737–1740. [Google Scholar]
- Hussain, M.; Hamid, M.I.; Ghazanfar, M.U.; Akhtar, N.; Raza, M. First report of fruit rot of strawberry caused by Geotrichum candidum in Pakistan. Plant. Dis. 2016, 100, 1948. [Google Scholar] [CrossRef]
- Michailides, T.J.; Morgan, D.P.; Day, K.R. First report of sour rot of California peaches and nectarines caused by yeasts. Plant Dis. 2004, 88, 222. [Google Scholar] [CrossRef]
- Alam, M.W.; Rehman, A.; Malik, A.U.; Iqbal, Z.; Amin, M.; Ali, S.; Hameed, A.; Sarfraz, S. First Report of Geotrichum candidum Causing Postharvest Sour Rot of Peach in Punjab, Pakistan. Plant Dis. 2017, 101, 1543. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
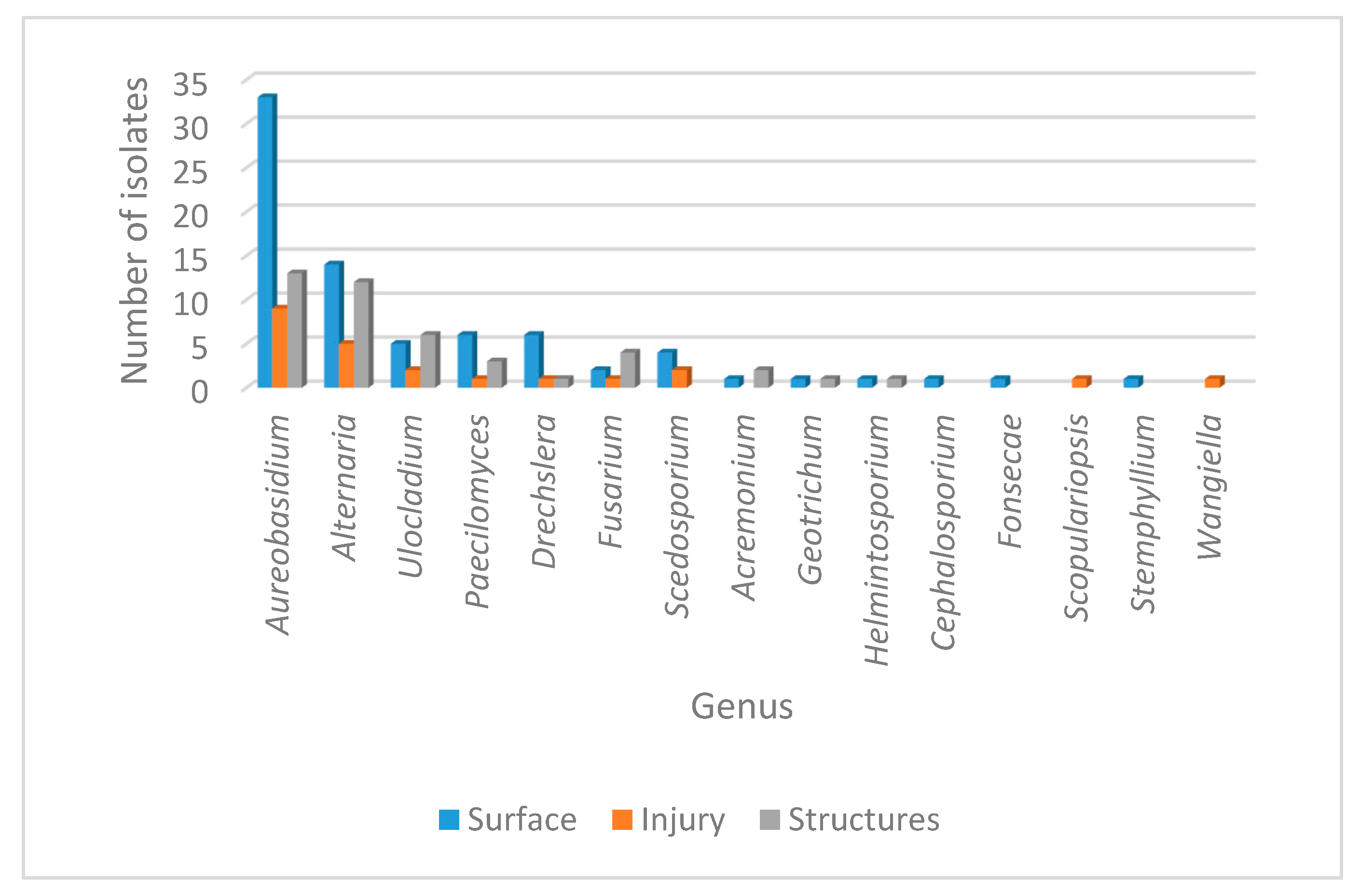
| Genus | Surface | Injury | Structures | Total |
|---|---|---|---|---|
| Aureobasidium sp. | 32 | 9 | 13 | 54 |
| Alternaria sp. | 14 | 5 | 12 | 31 |
| Ulocladium sp. | 5 | 2 | 6 | 13 |
| Paecilomyces sp. | 6 | 1 | 3 | 10 |
| Drechslera sp. | 6 | 1 | 1 | 8 |
| Fusarium sp. | 2 | 1 | 4 | 7 |
| Scedosporium sp. | 4 | 2 | 0 | 6 |
| Acremonium sp. | 1 | 0 | 2 | 3 |
| Geotrichum sp. | 1 | 0 | 1 | 2 |
| Helmintosporium sp. | 1 | 0 | 1 | 2 |
| Cephalosporium sp. | 1 | 0 | 0 | 1 |
| Fonsecae sp. | 1 | 0 | 0 | 1 |
| Phialophora sp. | 1 | 0 | 0 | 1 |
| Scopulariopsis sp. | 0 | 1 | 0 | 1 |
| Stemphylium sp. | 1 | 0 | 0 | 1 |
| Wangiella sp. | 0 | 1 | 0 | 1 |
| Candida sp. | 4 | 2 | 0 | 6 |
| Trichosporum sp. | 0 | 0 | 1 | 1 |
| Total | 80 | 25 | 44 | 149 |
| Genus | Isolate |
|---|---|
| Alternaria sp. | UAP035 UAP119 UAP127 |
| Aureobasidium sp. | UAP078 UAP087 UAP096 |
| Fusarium sp. | UAP118 UAP140 UAP168 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Cerezo, S.; Meneses-Sánchez, M.d.l.C.; Martínez-Contreras, R.D.; Martínez-Montiel, N. Unraveling the Fungal Community Associated with Leaf Spot on Crataegus sp. Microorganisms 2020, 8, 459. https://doi.org/10.3390/microorganisms8030459
Salazar-Cerezo S, Meneses-Sánchez MdlC, Martínez-Contreras RD, Martínez-Montiel N. Unraveling the Fungal Community Associated with Leaf Spot on Crataegus sp. Microorganisms. 2020; 8(3):459. https://doi.org/10.3390/microorganisms8030459
Chicago/Turabian StyleSalazar-Cerezo, Sonia, María de la Cruz Meneses-Sánchez, Rebeca D. Martínez-Contreras, and Nancy Martínez-Montiel. 2020. "Unraveling the Fungal Community Associated with Leaf Spot on Crataegus sp." Microorganisms 8, no. 3: 459. https://doi.org/10.3390/microorganisms8030459