Genome-Scale Metabolic Model Reconstruction and in Silico Investigations of Methane Metabolism in Methylosinus trichosporium OB3b



Abstract
:1. Introduction
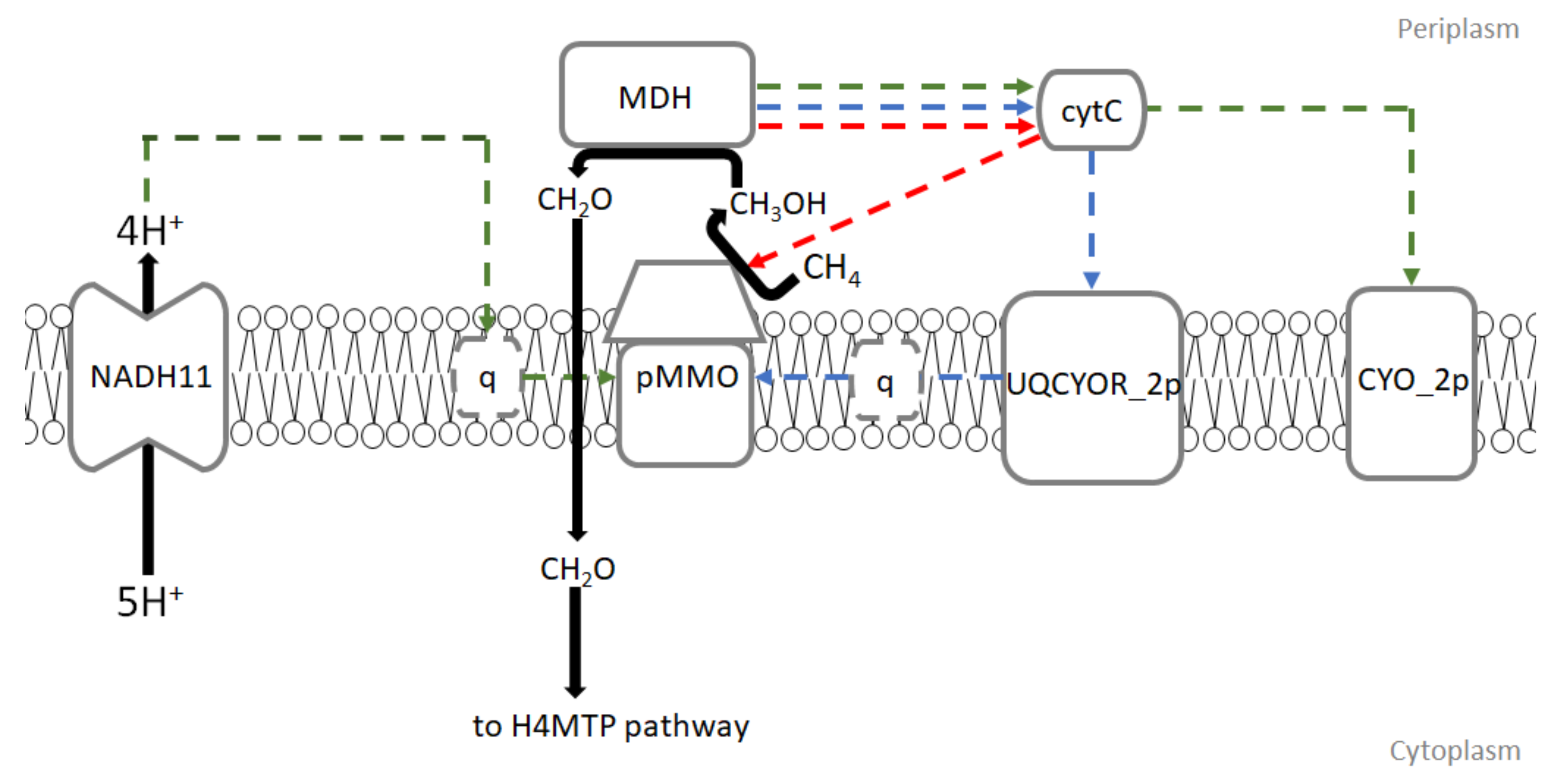
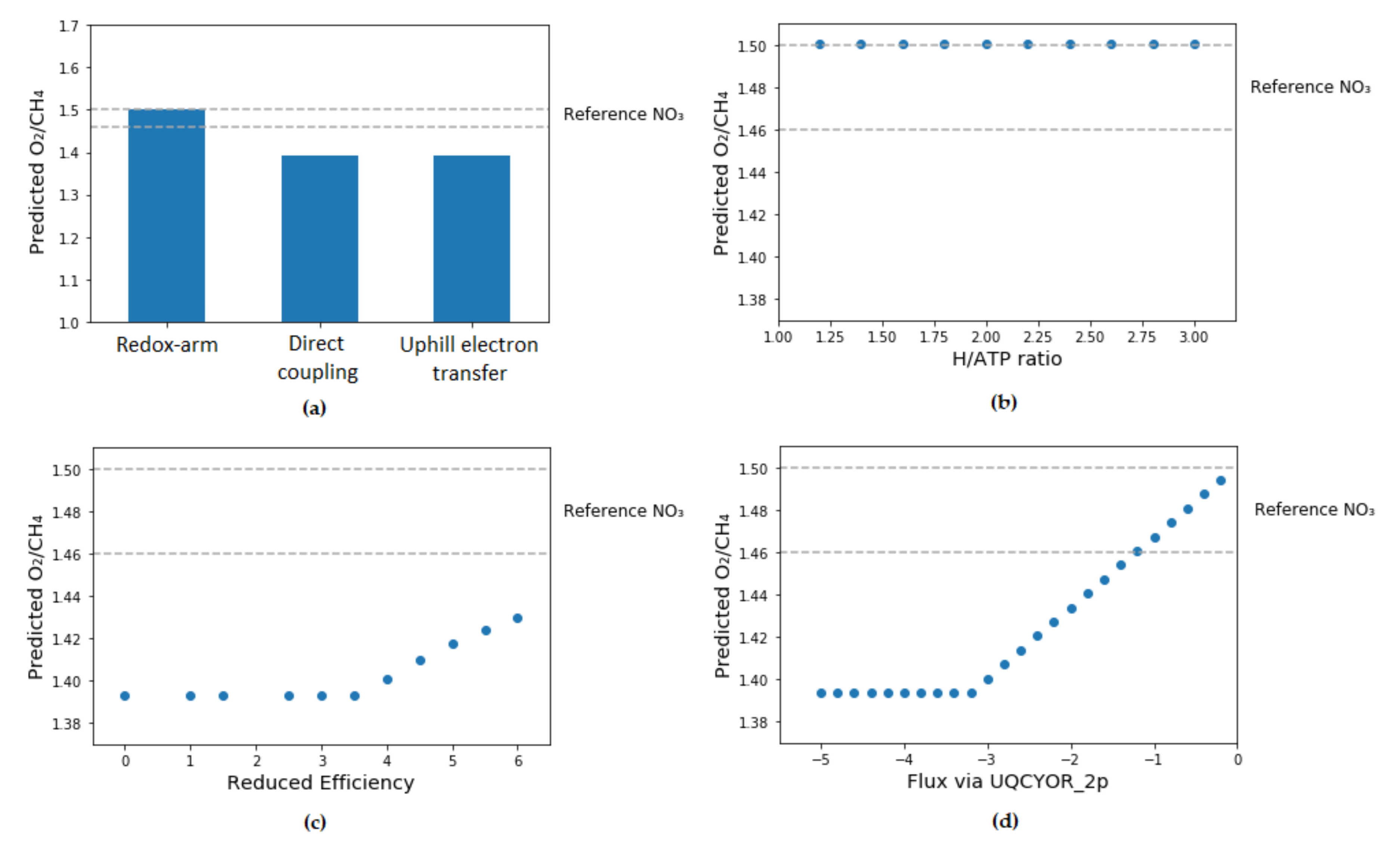
- Redox-arm mode: In this mode, ubiquinol is the electron donor for methane oxidation while methanol dehydrogenase (MDH) gives electrons via cytochrome c directly to complex IV. This leads up to proton motive force and ATP production. Recently, this mode of electron transfer has been suggested to be the main mode in Methylocystis, another type II methanotroph genus [17,18].
- Direct coupling mode: In this mode, methanol dehydrogenase passes electrons directly to pMMO with cytochrome c as an electron donor for methane oxidation.
- In uphill electron transfer, a reverse electron flow from cytochrome c to ubiquinone via ubiquinol-cytochrome-c reductase occurs. This mode is expected to significantly reduce efficiency of electron transport chain in methanotrophs.
2. Materials and Methods
2.1. Model Reconstruction and Curation
- To keep reactions balanced and to avoid metabolite production out of nowhere, reactions in the draft model were checked for mass and charge balance;
- To avoid unfeasible ATP production in the model, only reactions related to electron transport chain and lower glycolysis (phosphoglycerate kinase and pyruvate kinase) were allowed to produce ATP;
- To avoid proton flux outside the cell, only reactions of electron transport chain or reactions linked to some energy cost were allowed to have flux towards exterior of the cell;
- To make the draft model user-friendly and comparable with other models, all reaction and metabolite IDs were changed to BiGG Models IDs [40].
2.2. Applied Constraints
2.3. Used Software
3. Results and Discussion
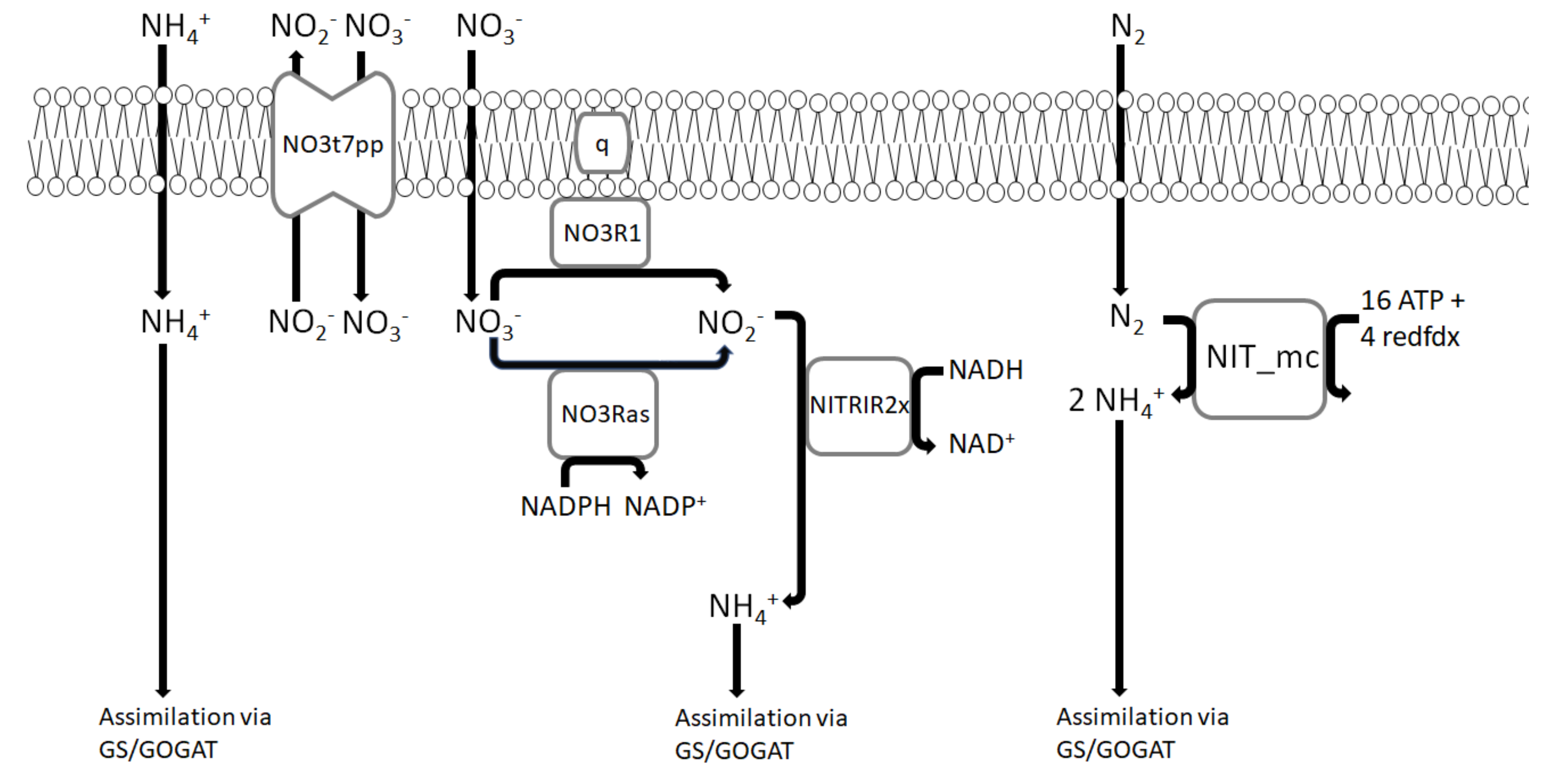
3.1. Nitrogen Metabolism
3.2. Determination of Mode of Electron Transfer to pMMO
3.3. Model Validation and Predictions under Different Environmental Conditions
- In the first pathway, hydroxylamine is oxidized to nitrite via hydroxylamine oxidoreductase which is then reduced to nitric oxide and nitrous oxide. Despite the fact that no gene for hydroxylamine oxidoreductase was found in M. trichosporium OB3b genome, this reaction (model reaction ID: HAORipp) was added due to multiple reports of nitric oxide and nitrous oxide production by M. trichosporium OB3b [42,64].
- In the second pathway, the formed hydroxylamine is reduced back to ammonium. The genome of M. trichosporium OB3b possesses a gene for hydroxylamine reductase (CQW49_14985) and the corresponding reaction (model reaction ID: HAMR) was added to the model.
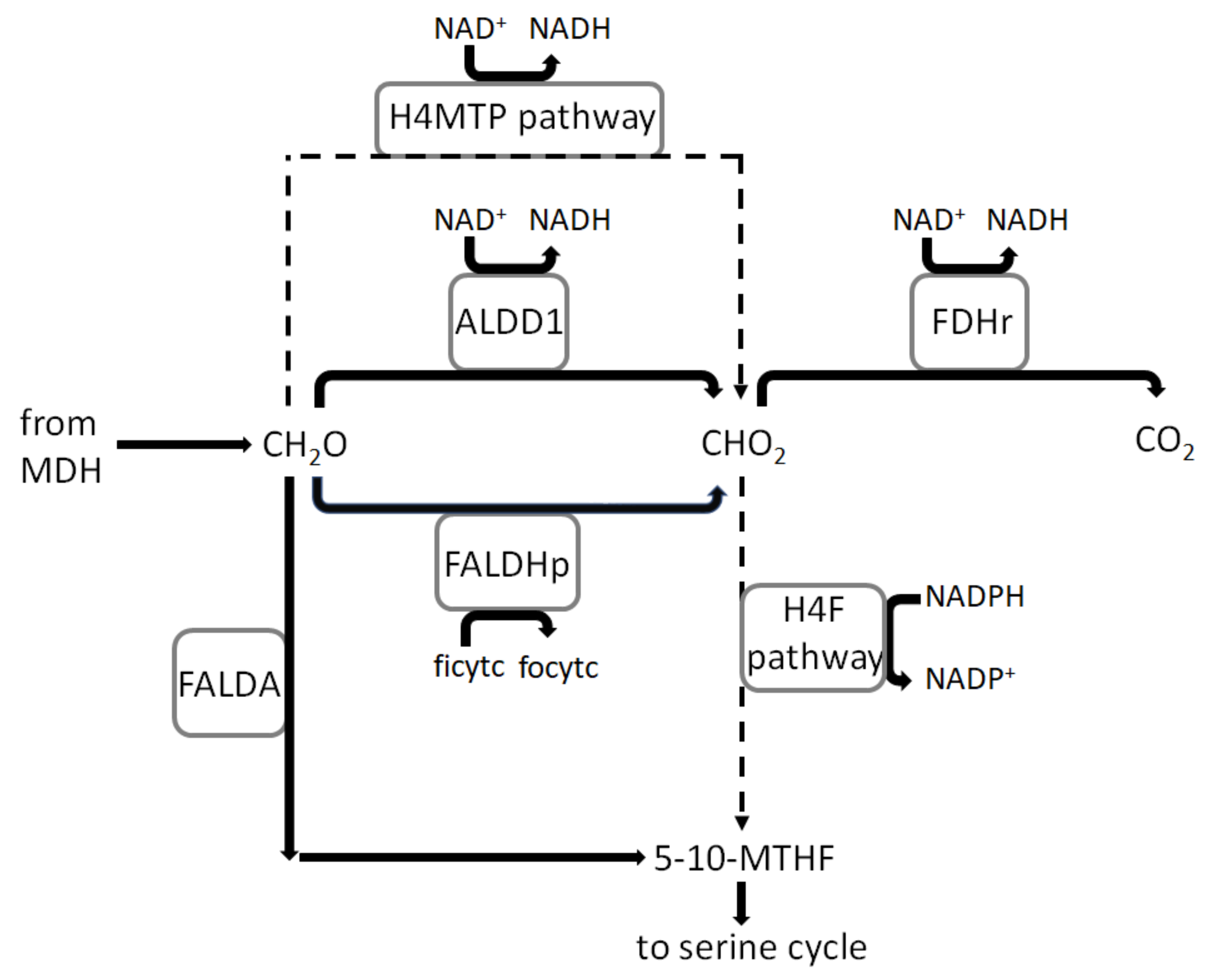
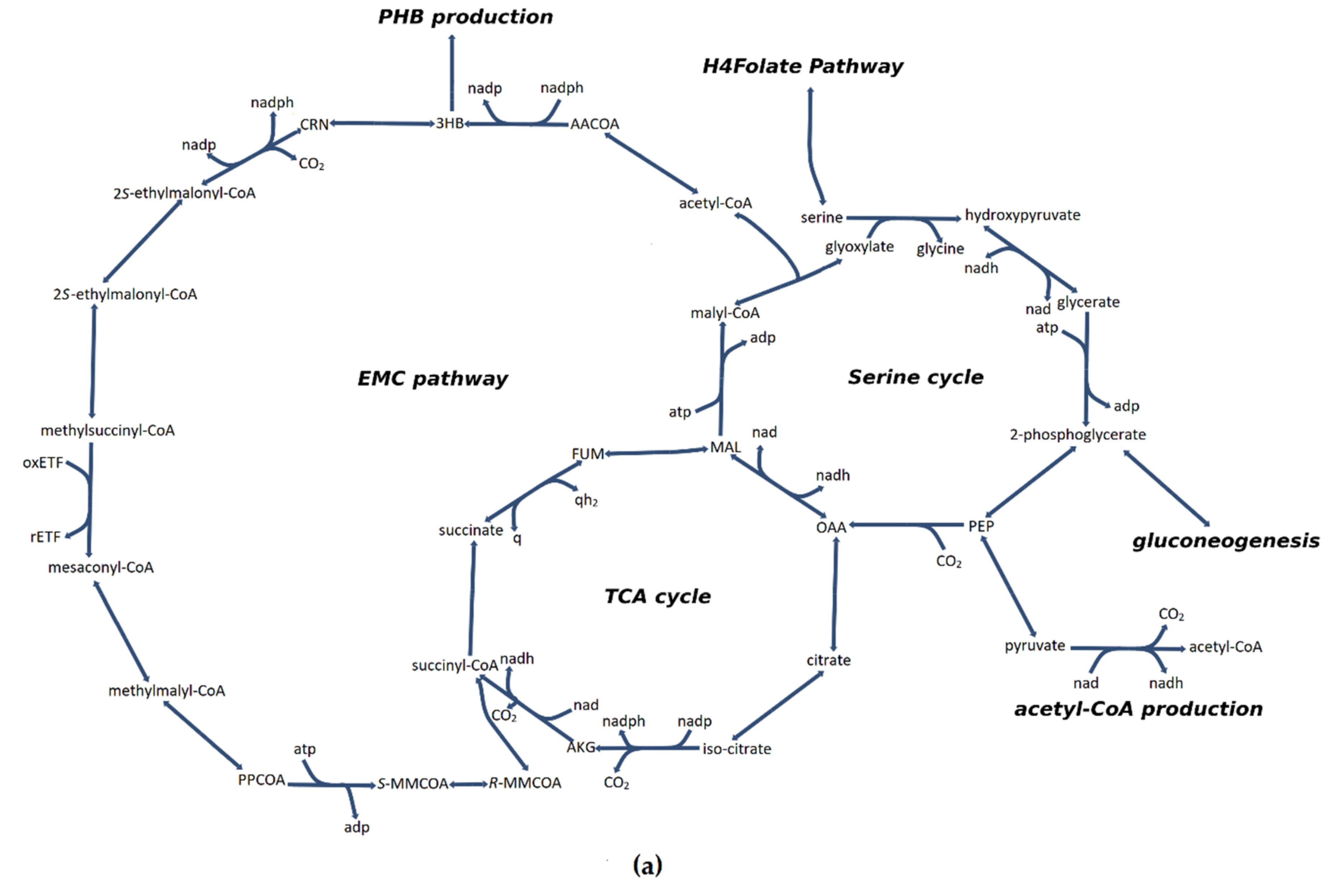
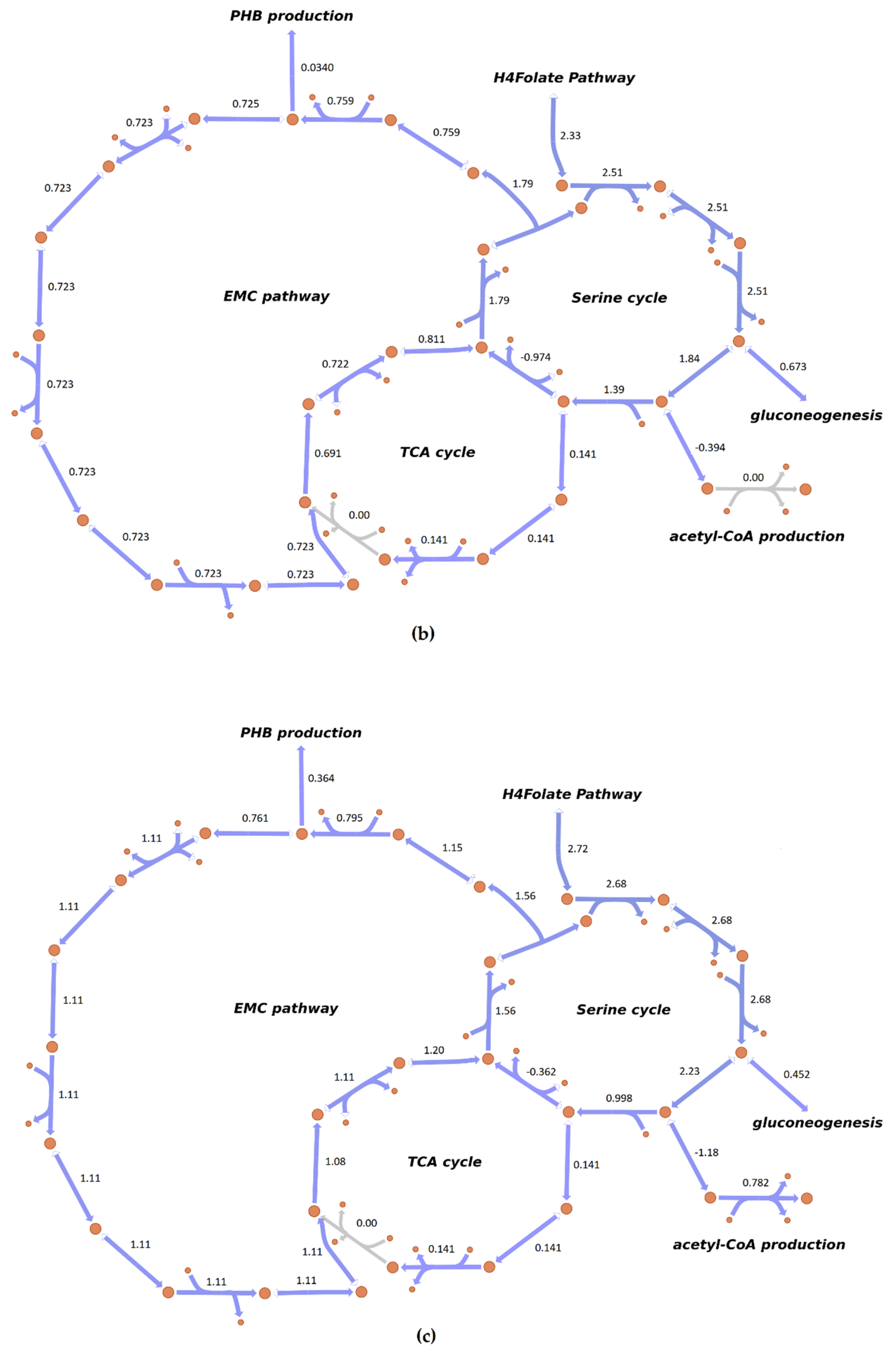
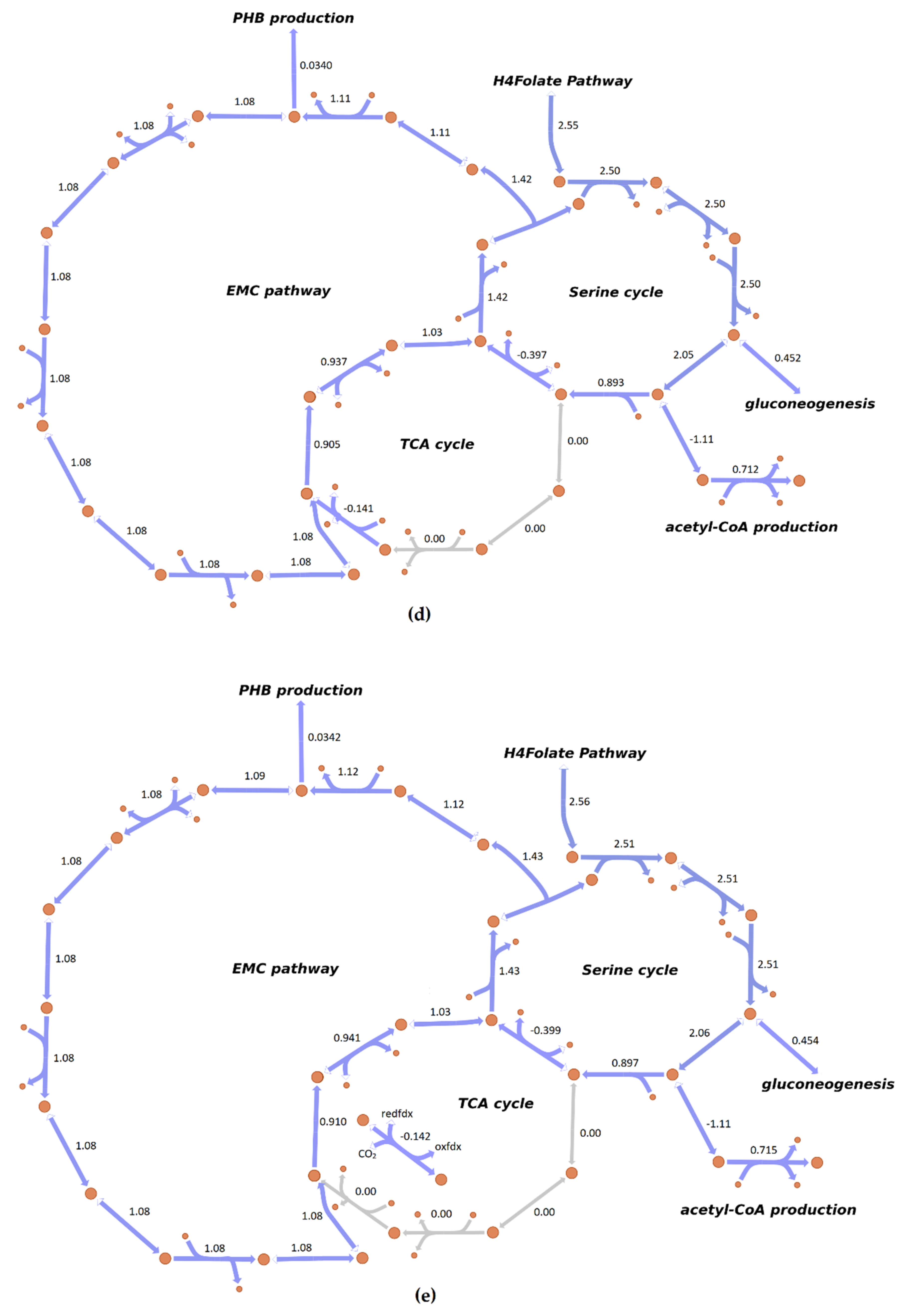
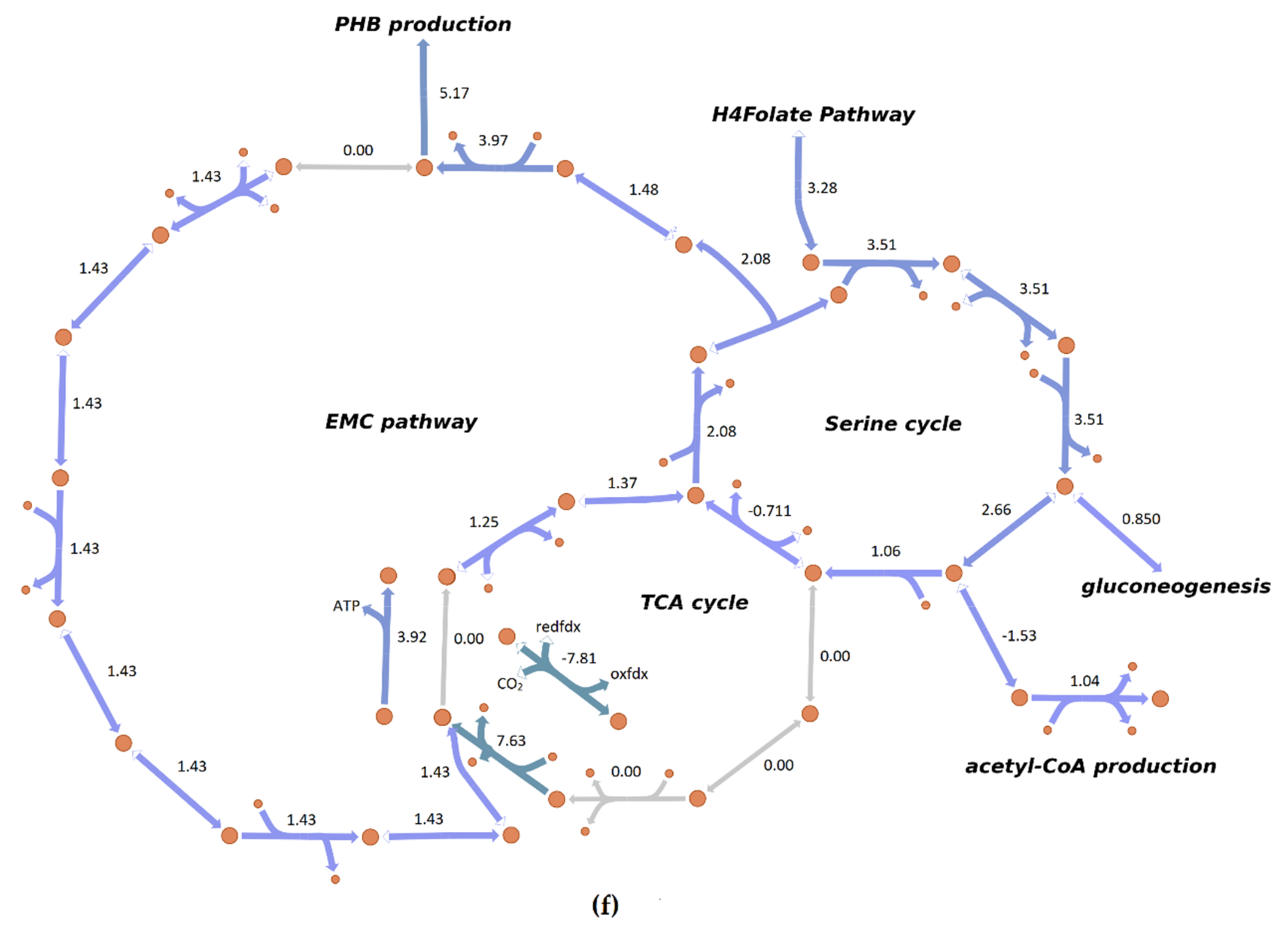
3.4. Flux Distribution in Central Carbon Metabolism
3.5. Final Model Statistics
3.6. Future Research Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stein, L.Y.; Yoon, S.; Semrau, J.D.; DiSpirito, A.A.; Crombie, A.; Murrell, J.C.; Vuilleumier, S.; Kalyuzhnaya, M.G.; Op den Camp, H.J.M.; Bringel, F.; et al. Genome sequence of the obligate methanotroph Methylosinus trichosporium strain OB3b. J. Bacteriol. 2010, 192, 6497–6498. [Google Scholar] [CrossRef] [Green Version]
- Whittenbury, R.; Phillips, K.H.; Wilkinson, J.F. Enrichment, isolation and some properties of methane-utilizing bacteria. J. Gen. Microbiol. 1970, 61, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Gu, W.; Park, D.; Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanobactin from Methylosinus trichosporium OB3b inhibits N2O reduction in denitrifiers. ISME J. 2018, 12, 2086–2089. [Google Scholar] [CrossRef] [Green Version]
- McCabe, J.W.; Vangala, R.; Angel, L.A. Binding selectivity of methanobactin from Methylosinus trichosporium OB3b for copper(I), silver(I), zinc(II), nickel(II), cobalt(II), manganese(II), lead(II), and iron(II). J. Am. Soc. Mass Spectrom. 2017, 28, 2588–2601. [Google Scholar] [CrossRef] [PubMed]
- Krentz, B.D.; Mulheron, H.J.; Semrau, J.D.; DiSpirito, A.A.; Bandow, N.L.; Haft, D.H.; Vuilleumier, S.; Murrell, J.C.; McEllistrem, M.T.; Hartsel, S.C.; et al. A comparison of methanobactins from Methylosinus trichosporium OB3b and Methylocystis strain SB2 predicts methanobactins are synthesized from diverse peptide precursors modified to create a common core for binding and reducing copper ions. Biochemistry 2010, 49, 10117–10130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.W.; Zea, C.J.; Do, Y.S.; Semrau, J.D.; Antholine, W.E.; Hargrove, M.S.; Pohl, N.L.; Boyd, E.S.; Geesey, G.G.; Hartsel, S.C.; et al. Spectral, kinetic, and thermodynamic properties of Cu(I) and Cu(II) binding by methanobactin from Methylosinus trichosporium OB3b. Biochemistry 2006, 45, 1442–1453. [Google Scholar] [CrossRef] [PubMed]
- Pesch, M.-L.; Christl, I.; Barmettler, K.; Kraemer, S.M.; Kretzschmar, R. Isolation and purification of Cu-free methanobactin from Methylosinus trichosporium OB3b. Geochem. Trans. 2011, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Hakemian, A.S.; Kondapalli, K.C.; Telser, J.; Hoffman, B.M.; Stemmler, T.L.; Rosenzweig, A.C. The metal centers of particulate methane monooxygenase from Methylosinus trichosporium OB3b. Biochemistry 2008, 47, 6793–6801. [Google Scholar] [CrossRef] [Green Version]
- Miyaji, A.; Rosenzweig, A.C.; Ragsdale, S.W. Chapter fourteen - particulate methane monooxygenase from Methylosinus trichosporium OB3b. In Methods in Methane Metabolism, Part B: Methanotrophy; Rosenzweig, A.C., Ragsdale, S.W., Eds.; Academic Press: San Diego, CA, USA, 2011; Volume 495, pp. 211–225. [Google Scholar]
- Takeguchi, M.; Ohashi, M.; Okura, I. Role of iron in particulate methane monooxygenase from Methylosinus trichosporium OB3b. BioMetals 1999, 12, 123–129. [Google Scholar] [CrossRef]
- Fox, B.G.; Froland, W.A.; Dege, J.E.; Lipscomb, J.D. Methane monooxygenase from Methylosinus trichosporium OB3b. Purification and properties of a three-component system with high specific activity from a type II methanotroph. J. Biol. Chem. 1989, 264, 10023–10033. [Google Scholar]
- Phelps, P.A.; Agarwal, S.K.; Speitel, G.E.; Georgiou, G. Methylosinus trichosporium OB3b mutants having constitutive expression of soluble methane monooxygenase in the presence of high levels of copper. Appl. Environ. Microbiol. 1992, 58, 3701–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elango, N.; Radhakrishnan, R.; Froland, W.A.; Wallar, B.J.; Earhart, C.A.; Lipscomb, J.D.; Ohlendorf, D.H. Crystal structure of the hydroxylase component of methane monooxygenase from Methylosinus trichosporium OB3b. Protein Sci. 1997, 6, 556–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiol. Rev. 2010, 34, 496–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakemian, A.S.; Rosenzweig, A.C. The biochemistry of methane oxidation. Annu Rev. Biochem. 2007, 76, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P.; Sayler, G.S. Optimization and maintenance of soluble methane monooxygenase activity in Methylosinus trichosporium OB3b. Biodegradation 1994, 5, 1–11. [Google Scholar]
- Bordel, S.; Rojas, A.; Muñoz, R. Reconstruction of a genome scale metabolic model of the polyhydroxybutyrate producing methanotroph Methylocystis parvus OBBP. Microb. Cell Fact. 2019, 18, 104. [Google Scholar] [CrossRef]
- Bordel, S.; Rodríguez, Y.; Hakobyan, A.; Rodríguez, E.; Lebrero, R.; Muñoz, R. Genome scale metabolic modeling reveals the metabolic potential of three Type II methanotrophs of the genus Methylocystis. Metab. Eng. 2019, 54, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Doronina, N.V.; Ezhov, V.A.; Trotsenko, Y.A. Growth of Methylosinus trichosporium OB3b on methane and poly-β-hydroxybutyrate biosynthesis. Appl. Biochem. Microbiol. 2008, 44, 182–185. [Google Scholar] [CrossRef]
- Williams, A.M. The Biochemistry and Physiology of Poly-Beta-Hydroxybutyrate Metabolism in Methylosinus trichosporium OB3b. PhD Thesis, Cranfield Institute of Technology, Cranfield, UK, 1988. Available online: https://dspace.lib.cranfield.ac.uk/handle/1826/4449 (accessed on 3 January 2020).
- Asenjo, J.A.; Suk, J.S. Microbial conversion of methane into poly-β-hydroxybutyrate (PHB): Growth and intracellular product accumulation in a type II methanotroph. J. Ferment. 1986, 64, 271–278. [Google Scholar] [CrossRef]
- Shah, N.N.; Hanna, M.L.; Taylor, R.T. Batch cultivation of Methylosinus trichosporium OB3b: V. Characterization of poly-beta-hydroxybutyrate production under methane-dependent growth conditions. Biotechnol. Bioeng. 1996, 49, 161–171. [Google Scholar] [CrossRef]
- Scott, D.; Brannan, J.; Higgins, I.J. The effect of growth conditions on intracytoplasmic membranes and methane mono-oxygenase activities in Methylosinus trichosporium OB3b. Microbiology 1981, 125, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Lontoh, S.; Semrau, J.D. Methane and trichloroethylene degradation by Methylosinus trichosporium OB3b expressing particulate methane monooxygenase. Appl Environ Microbiol 1998, 64, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Tsien, H.C.; Brusseau, G.A.; Hanson, R.S.; Waclett, L.P. Biodegradation of trichloroethylene by Methylosinus trichosporium OB3b. Appl Environ Microbiol 1989, 55, 3155–3161. [Google Scholar] [CrossRef] [Green Version]
- Fitch, M.W.; Speitel, G.E.; Georgiou, G. Degradation of trichloroethylene by methanol-grown cultures of Methylosinus trichosporium OB3b PP358. Appl. Environ. Microbiol. 1996, 62, 1124–1128. [Google Scholar] [CrossRef] [Green Version]
- Strong, P.J.; Laycock, B.; Mahamud, S.N.S.; Jensen, P.D.; Lant, P.A.; Tyson, G.; Pratt, S. The opportunity for high-performance biomaterials from methane. Microorganisms 2016, 4, 11. [Google Scholar] [CrossRef]
- Ritala, A.; Häkkinen, S.T.; Toivari, M.; Wiebe, M.G. Single Cell Protein—State-of-the-art, industrial landscape and patents 2001–2016. Front. Microbiol. 2017, 8, 2009. [Google Scholar] [CrossRef]
- Ro, S.Y.; Rosenzweig, A.C. Recent advances in the genetic manipulation of Methylosinus trichosporium OB3b. Methods Enzymol. 2018, 605, 335–349. [Google Scholar]
- Thiele, I.; Palsson, B.Ø. A protocol for generating a high-quality genome-scale metabolic reconstruction. Nat. Protoc. 2010, 5, 93–121. [Google Scholar] [CrossRef] [Green Version]
- Peyraud, R.; Schneider, K.; Kiefer, P.; Massou, S.; Vorholt, J.A.; Portais, J. Genome-scale reconstruction and system level investigation of the metabolic network of Methylobacterium extorquens AM1. BMC Syst. Biol. 2011, 5, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Heil, J.R.; Lynch, M.D.J.; Cheng, J.; Matysiakiewicz, O.; D’Alessio, M.; Charles, T.C. The completed PacBio single-molecule real-time sequence of Methylosinus trichosporium Strain OB3b reveals the presence of a third large plasmid. Genome Announc. 2017, 5, e01349–17. [Google Scholar] [CrossRef] [Green Version]
- Sonnhammer, E.L.L.; Östlund, G. InParanoid 8: Orthology analysis between 273 proteomes, mostly eukaryotic. Nucleic Acids Res. 2015, 43, D234–D239. [Google Scholar] [CrossRef]
- Norsigian, C.J.; Fang, X.; Seif, Y.; Monk, J.M.; Palsson, B.O. A workflow for generating multi-strain genome-scale metabolic models of prokaryotes. Nat. Protoc. 2020, 15, 1–14. [Google Scholar] [CrossRef]
- Lieven, C.; Petersen, L.A.H.; Jørgensen, S.B.; Gernaey, K.V.; Herrgard, M.J.; Sonnenschein, N. A genome-scale metabolic model for Methylococcus capsulatus (Bath) suggests reduced efficiency electron transfer to the particulate methane monooxygenase. Front. Microbiol. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Matsen, J.B.; Yang, S.; Stein, L.Y.; Beck, D.A.C.; Kalyuzhanaya, M.G. Global molecular analyses of methane metabolism in methanotrophic alphaproteobacterium, Methylosinus trichosporium OB3b. Part I: Transcriptomic study. Front. Microbiol. 2013, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Kalyuzhanaya, M.G.; Yang, S.; Matsen, J.B.; Konopka, M.; Green-Saxena, A.; Clubb, J.; Sadilek, M.; Orphan, V.J.; Beck, D. Global molecular analyses of methane metabolism in methanotrophic alphaproteobacterium, Methylosinus trichosporium OB3b. Part II. Metabolomics and 13C-labeling study. Front. Microbiol. 2013, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Methanotroph Commons. Available online: http://www.methanotroph.org/wiki/metabolic-pathways/ (accessed on 3 January 2020).
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [Green Version]
- King, Z.A.; Lu, J.; Dräger, A.; Miller, P.; Federowicz, S.; Lerman, J.A.; Ebrahim, A.; Palsson, B.O.; Lewis, N.E. BiGG Models: A platform for integrating, standardizing and sharing genome-scale models. Nucleic Acids Res. 2016, 44, D515–D522. [Google Scholar] [CrossRef]
- Graham, D.W.; Chaudhary, J.A.; Hanson, R.S.; Arnold, R.G. Factors affecting competition between Type I and Type II methanotrophs in two-organism, continuous-flow reactors. Microb. Ecol. 1993, 25, 1–17. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, J.; Wang, X.; Zhang, Y. Coupled effects of methane monooxygenase and nitrogen source on growth and poly-β-hydroxybutyrate (PHB) production of Methylosinus trichosporium OB3b. J. Environ. Sci. 2017, 52, 49–57. [Google Scholar] [CrossRef]
- Park, S.; Hanna, L.; Taylor, R.T.; Droege, M.W. Batch cultivation of Methylosinus trichosporium OB3b. I: Production of soluble methane monooxygenase. Biotechnol. Bioeng. 1991, 38, 423–433. [Google Scholar] [CrossRef]
- Lee, S.-W.; Keeney, D.R.; Lim, D.-H.; Dispirito, A.A.; Semrau, J.D. Mixed pollutant degradation by Methylosinus trichosporium OB3b expressing either soluble or particulate methane monooxygenase: Can the tortoise beat the hare? Appl. Environ. Microbiol. 2006, 72, 7503–7509. [Google Scholar] [CrossRef] [Green Version]
- Sun A., K.; Wood T., K. Trichloroethylene degradation and mineralization by pseudomonads and Methylosinus trichosporium OB3b. Appl. Microbiol. Biotechnol. 1996, 45, 248–256. [Google Scholar] [CrossRef]
- Rodrigues, A.S.; Salgado, B.V. Analysis of methane biodegradation by Methylosinus trichosporium OB3b. Braz. J. Microbiol. 2009, 40, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.Y.; Hur, D.H.; Lee, J.H.; Park, C.-H.; Chang, I.S.; Lee, J.W.; Lee, E.Y. Batch Conversion of methane to methanol using Methylosinus trichosporium OB3b as biocatalyst. J. Microbiol. Biotechn. 2015, 25, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Tays, C.; Guarnieri, M.T.; Sauvageau, D.; Stein, L.Y. Combined effects of carbon and nitrogen source to optimize growth of proteobacterial methanotrophs. Front. Microbiol. 2018, 9, 2239. [Google Scholar] [CrossRef]
- Machado, D.; Andrejev, S.; Tramontano, M.; Patil, K.R. Fast automated reconstruction of genome-scale metabolic models for microbial species and communities. Nucleic Acids Res. 2018, 46, 7542–7553. [Google Scholar] [CrossRef]
- Lewis, N.E.; Hixson, K.K.; Conrad, T.M.; Lerman, J.A.; Charusanti, P.; Polpitiya, A.D.; Adkins, J.N.; Schramm, G.; Purvine, S.O.; Lopez-Ferrer, D.; et al. Omic data from evolved E. coli are consistent with computed optimal growth from genome-scale models. Mol. Syst. Biol. 2010, 6, 390. [Google Scholar] [CrossRef]
- Ebrahim, A.; Lerman, J.A.; Palsson, B.O.; Hyduke, D.R. COBRApy: Constraints-based reconstruction and analysis for Python. BMC Syst. Biol. 2013, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- King, Z.A.; Dräger, A.; Ebrahim, A.; Sonnenschein, N.; Lewis, N.E.; Palsson, B.O. Escher: A web application for building, sharing, and embedding data-rich visualizations of biological pathways. PLoS Comput. Biol. 2015, 11, e1004321. [Google Scholar] [CrossRef] [Green Version]
- Murrell, J.C.; Dalton, H. Ammonia assimilation in Methylococcus capsulatus (Bath) and other obligate methanotrophs. Microbiology 1983, 129, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Chu, K.-H.; Alvarez-Cohen, L. Effect of nitrogen source on growth and trichloroethylene degradation by methane-oxidizing bacteria. Appl. Environ. Microbiol. 1998, 64, 3451–3457. [Google Scholar] [CrossRef] [Green Version]
- De la Torre, A.; Metivier, A.; Chu, F.; Laurens, L.M.L.; Beck, D.A.C.; Pienkos, P.T.; Lidstrom, M.E.; Kalyuzhnaya, M.G. Genome-scale metabolic reconstructions and theoretical investigation of methane conversion in Methylomicrobium buryatense strain 5G(B1). Microb. Cell Fact. 2015, 14, 188. [Google Scholar] [CrossRef] [Green Version]
- Takeguchi, M.; Miyakawa, K.; Okura, I. Properties of the membranes containing the particulate methane monooxygenase from Methylosinus trichosporium OB3b. Biometals 1998, 11, 229–234. [Google Scholar] [CrossRef]
- Miyaji, A. Particulate Methane Monooxygenase from Methylosinus trichosporium OB3b. In Methods in Enzymology; Elsevier: Cambridge, MA, USA, 2011; Volume 495, pp. 211–225. [Google Scholar]
- Methylosinus Trichosporium UniProt Search Results. Available online: https://www.uniprot.org/uniprot/?query=methylosinus%20trichosporium&fil=reviewed%3Ayes&sort=score (accessed on 4 December 2019).
- Methylococcus Capsulatus UniProt Search Results. Available online: https://www.uniprot.org/uniprot/?query=methylococcus+capsulatus+AND+reviewed%3Ayes&sort=score (accessed on 4 December 2019).
- Herbert, D.; Elsworth, R.; Telling, R.C. The continuous culture of bacteria; a theoretical and experimental study. J. Gen. Microbiol. 1956, 14, 601–622. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-de-Cossio-Diaz, J.; Leon, K.; Mulet, R. Characterizing steady states of genome-scale metabolic networks in continuous cell cultures. PLoS Comput. Biol. 2017, 13, e1005835. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.A.; Nyerges, G.; Kozlowski, J.A.; Poret-Peterson, A.T.; Stein, L.Y.; Klotz, M.G. Model of the molecular basis for hydroxylamine oxidation and nitrous oxide production in methanotrophic bacteria. FEMS Microbiol. Lett. 2011, 322, 82–89. [Google Scholar] [CrossRef]
- Sutka, R.L.; Ostrom, N.E.; Ostrom, P.H.; Breznak, J.A.; Gandhi, H.; Pitt, A.J.; Li, F. Distinguishing nitrous oxide production from nitrification and denitrification on the basis of isotopomer abundances. Appl. Environ. Microbiol. 2006, 72, 638–644. [Google Scholar] [CrossRef] [Green Version]
- Yoshinari, T. Nitrite and nitrous oxide production by Methylosinus trichosporium. Can. J. Microbiol. 1985, 31, 139–144. [Google Scholar] [CrossRef]
- Akberdin, I.R.; Thompson, M.; Hamilton, R.; Desai, N.; Alexander, D.; Henard, C.A.; Guarnieri, M.T.; Kalyuzhnaya, M.G. Methane utilization in Methylomicrobium alcaliphilum 20ZR: A systems approach. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Crowther, G.J.; Kosály, G.; Lidstrom, M.E. Formate as the main branch point for methylotrophic metabolism in Methylobacterium extorquens AM1. J. Bacteriol. 2008, 190, 5057–5062. [Google Scholar] [CrossRef] [Green Version]
- Trotsenko, Y.A.; Murrell, J.C. Metabolic aspects of aerobic obligate methanotrophy. Adv. Appl. Microbiol. 2008, 63, 183–229. [Google Scholar]
- Park, S.; Shah, N.N.; Taylor, R.T.; Droege, M.W. Batch cultivation of Methylosinus trichosporium OB3b: II. Production of particulate methane monooxygenase. Biotechnol. Bioeng. 1992, 40, 151–157. [Google Scholar] [CrossRef]
- Lieven, C.; Beber, M.E.; Olivier, B.G.; Bergmann, F.T.; Babaei, P.; Bartell, J.A.; Blank, L.M.; Chauhan, S.; Correia, K.; Diener, C.; et al. Memote: A community driven effort towards a standardized genome-scale metabolic model test suite. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.D.; Hwang, I.Y.; Lee, O.K.; Kim, D.; Kalyuzhnaya, M.G.; Mariyana, R.; Hadiyati, S.; Kim, M.S.; Lee, E.Y. Systematic metabolic engineering of Methylomicrobium alcaliphilum 20Z for 2,3-butanediol production from methane. Metab. Eng. 2018, 47, 323–333. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Lee, E.Y. Biological conversion of methane to putrescine using genome-scale model-guided metabolic engineering of a methanotrophic bacterium Methylomicrobium alcaliphilum 20Z. Biotechnol. Biofuels 2019, 12, 147. [Google Scholar] [CrossRef]
- Ang, K.S.; Lakshmanan, M.; Lee, N.-R.; Lee, D.-Y. Metabolic modeling of microbial community interactions for health, environmental and biotechnological applications. Curr. Genomics 2018, 19, 712–722. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO3−, pMMO | NO3−, sMMO | NH4+, pMMO | NH4+, sMMO | N2, pMMO | N2, sMMO | |
|---|---|---|---|---|---|---|
| Methane URF | −14.9 | −14.9 | −14.9 | −14.9 | −0.88 | −1.17 |
| Oxygen URF | −22.36 | −23.16 | −22.49 | −24.43 | −1.75 | −2.33 |
| CO2 URF | 9.97 | 10.50 | 7.75 | 9.65 | 0.875 | 1.17 |
| NH4 URF | 0 | 0 | −1.74 | −1.28 | 0 | 0 |
| N2 URF | 0 | 0 | 0 | 0 | 0 | 0 |
| NO3 URF | −1.20 | −1.07 | 0 | 0 | 0 | 0 |
| Uphill Electron Transfer Flux | 0 | 0 | 0 | 0 | 0 | 0 |
| Growth Rate | 0.123 | 0.109 | 0.178 | 0.131 | 0 | 0 |
| Growth Yield 1 | 0.51 | 0.46 | 0.74 | 0.55 | 0 | 0 |
| O2/CH4 Ratio | 1.50 | 1.55 | 1.51 | 1.64 | 2 | 2 |
| NH4/CH4 Ratio | 0 | 0 | 0.12 | 0.086 | 0 | 0 |
| N2/CH4 Ratio | 0 | 0 | 0 | 0 | 0 | 0 |
| NO3/CH4 Ratio | 0.08 | 0.07 | 0 | 0 | 0 | 0 |
| ATP/CH4 Ratio | 1.43 | 1.24 | 1.87 | 1.44 | 4 | 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naizabekov, S.; Lee, E.Y. Genome-Scale Metabolic Model Reconstruction and in Silico Investigations of Methane Metabolism in Methylosinus trichosporium OB3b. Microorganisms 2020, 8, 437. https://doi.org/10.3390/microorganisms8030437
Naizabekov S, Lee EY. Genome-Scale Metabolic Model Reconstruction and in Silico Investigations of Methane Metabolism in Methylosinus trichosporium OB3b. Microorganisms. 2020; 8(3):437. https://doi.org/10.3390/microorganisms8030437
Chicago/Turabian StyleNaizabekov, Sanzhar, and Eun Yeol Lee. 2020. "Genome-Scale Metabolic Model Reconstruction and in Silico Investigations of Methane Metabolism in Methylosinus trichosporium OB3b" Microorganisms 8, no. 3: 437. https://doi.org/10.3390/microorganisms8030437