Ivermectin Inhibits Bovine Herpesvirus 1 DNA Polymerase Nuclear Import and Interferes With Viral Replication


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
- UL30HA(F): 5′GCAAGCTTGCCACCATGTACCCATACGATGTTCCAGATTACGCTGACAGAGACTGCGAACAGGGC’3;
- UL30HA(R): 5′CGGAATTCTCAACTTTGATGGGGAGCTGCTGCTAGAATACCGAAGGCTGT’3; UL42FLAG(F): 5′GCAAGCTTGCCACCATGGATTACAAGGATGACGACGATAAGCTGCAGCCCCCCTCGCAT’3;
- UL42FLAG(R):5′CGGGATCCTTAAGGAGTTTCGCCCCCCTCCCCG’3.
2.2. Transfection of Cells with UL30-HA and UL42-FLAG Expressing Plasmids
2.3. Co-Immunoprecipitation Assay and Western Blot Analysis
2.4. Immunofluorescence and Confocal Laser Scanning Microscopy (CLSM)
2.5. Identification of cNLSs with Bioinformatics Analysis
2.6. Cells and Viruses
2.7. Compounds
2.8. MTT Cell Proliferation and Cytotoxicity Assay
2.9. Antiviral Activity of IVM in Cell Culture
2.10. Plaque Reduction Assays (PRAs)
2.11. Effect of IVM on BoHV-1 Binding
2.12. Effect of IVM on Virus Penetration
2.13. Statistical Analysis
3. Results
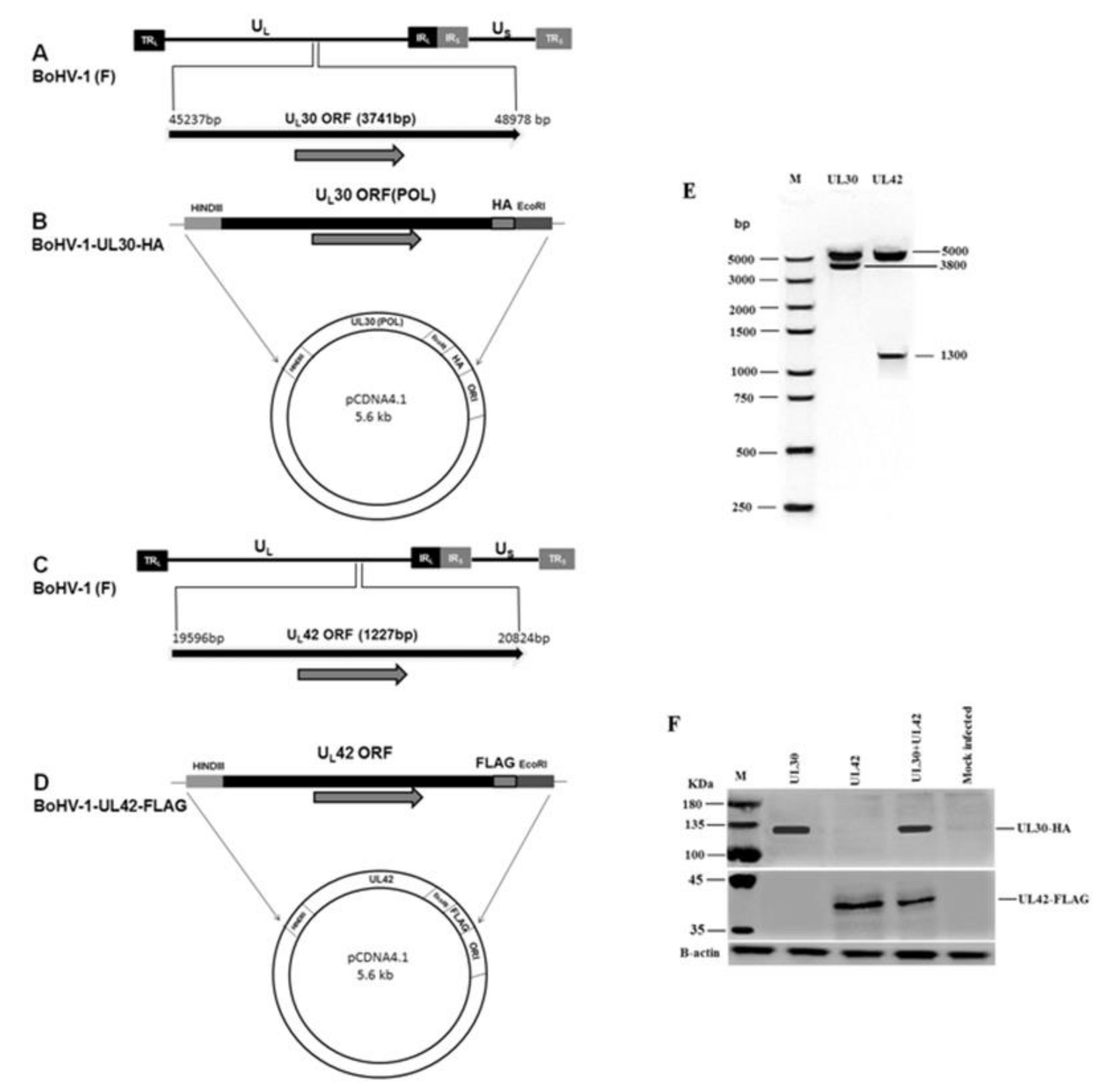
3.1. Construction and Characterization of BoHV-1 UL30-HA and UL42-FLAG Expression Plasmids
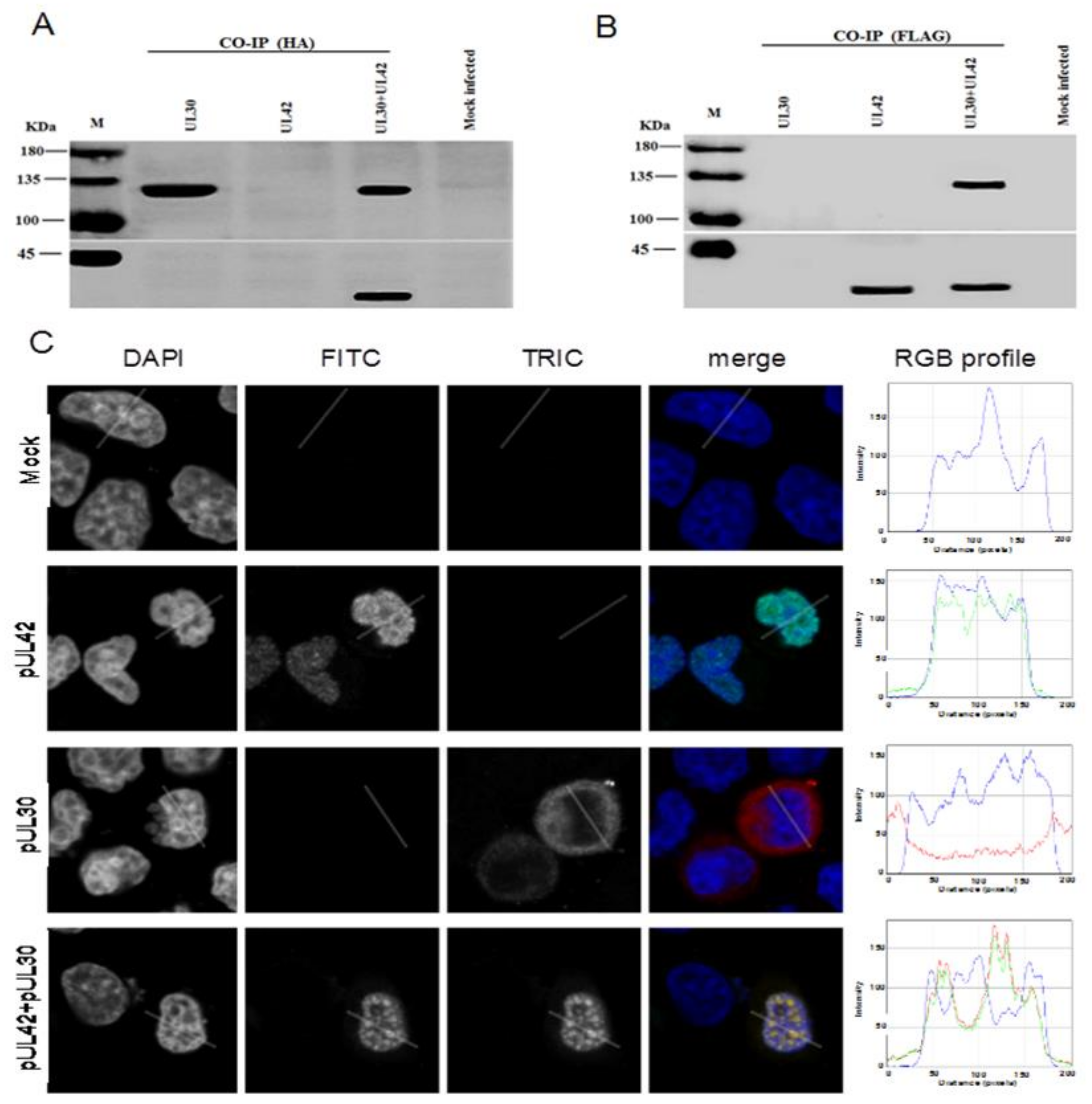
3.2. BoHV-1 UL30 and UL42 form a Complex in the Absence of other Viral Proteins
3.3. BoHV-1 pUL42 is Necessary and Sufficient for Nuclear Localization of pUL30
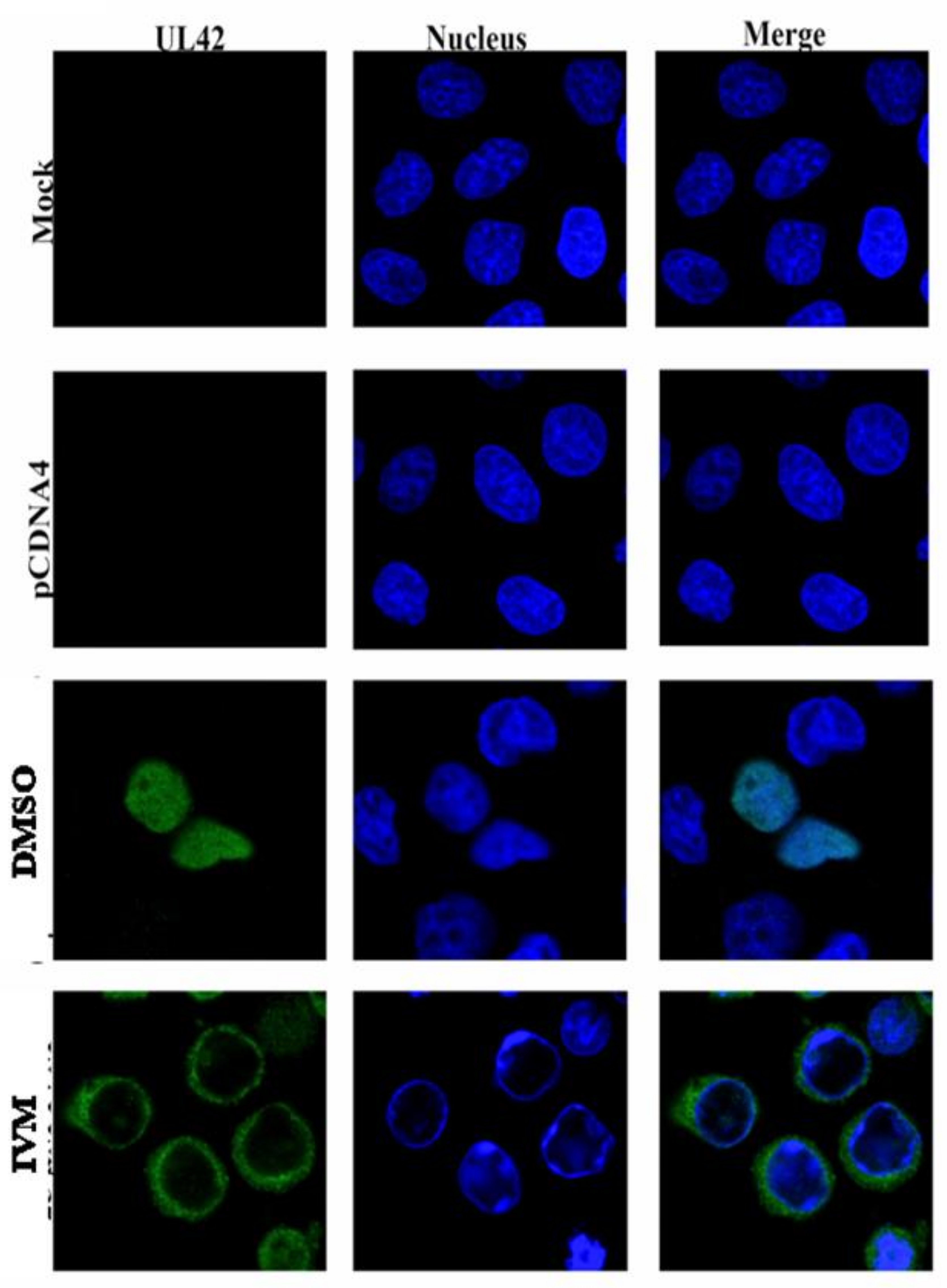
3.4. IVM Inhibits Nuclear Accumulation of BoHV-1 pUL42 Nuclear Accumulation
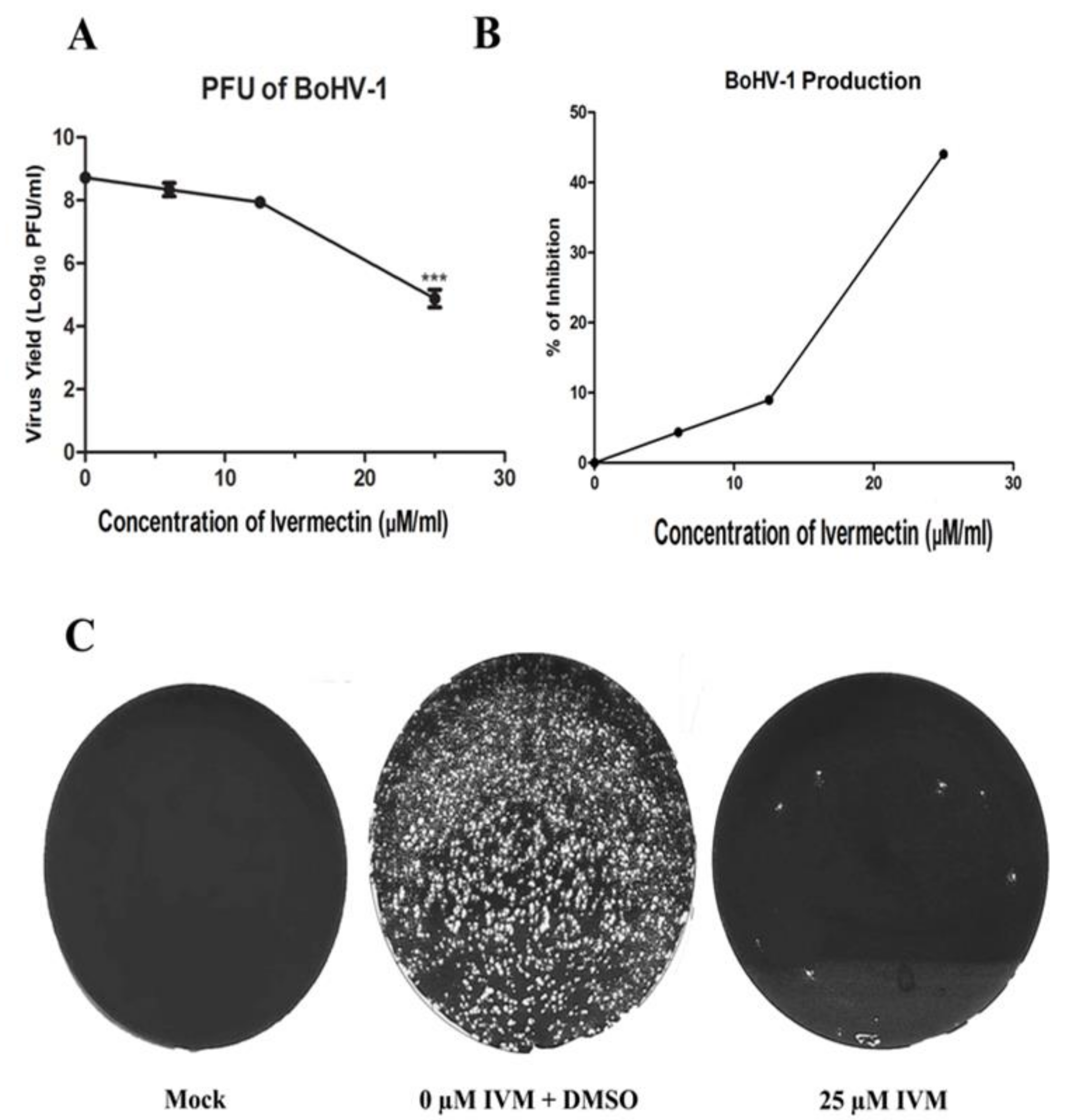
3.5. IVM Impairs BoHV-1 Life Cycle
3.6. IVM does not Markedly Affect Cell Viability.
3.7. IVM Does Not Interfere With BoHV-1 Attachment and Entry.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Raza, S.; Deng, M.; Shahin, F.; Yang, K.; Hu, C.; Chen, Y.; Chen, H.; Guo, A. A bovine herpesvirus 1 pUL51 deletion mutant shows impaired viral growth in vitro and reduced virulence in rabbits. Oncotarget 2016, 7, 12235–12253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.C.; Shen, H.Y.; Cheng, L.T.; Liu, S.S.; Chu, C.Y. Effectiveness of a BHV-1/BEFV bivalent vaccine against bovine herpesvirus type 1 infection in cattle. Res. Vet. Sci. 2016, 109, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.; Zhu, L.; Keel, B.N.; Smith, T.P.L.; Jones, C. The Wnt Signaling Pathway Is Differentially Expressed during the Bovine Herpesvirus 1 Latency-Reactivation Cycle: Evidence That Two Protein Kinases Associated with Neuronal Survival, Akt3 and BMPR2, Are Expressed at Higher Levels during Latency. J. Virol. 2018, 92, e01937-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Mayet, F.S.; Sawant, L.; Thunuguntla, P.; Jones, C. Combinatorial Effects of the Glucocorticoid Receptor and Kruppel-Like Transcription Factor 15 on Bovine Herpesvirus 1 Transcription and Productive Infection. J. Virol. 2017, 91, e00904-17. [Google Scholar] [CrossRef] [Green Version]
- Sawant, L.; Kook, I.; Vogel, J.L.; Kristie, T.M.; Jones, C. The Cellular Coactivator HCF-1 Is Required for Glucocorticoid Receptor-Mediated Transcription of Bovine Herpesvirus 1 Immediate Early Genes. J. Virol. 2018, 92, e00987-18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Workman, A.; Jones, C. Potential Role for a beta-Catenin Coactivator (High-Mobility Group AT-Hook 1 Protein) during the Latency-Reactivation Cycle of Bovine Herpesvirus 1. J. Virol. 2017, 91, e02132-16. [Google Scholar] [CrossRef] [Green Version]
- Muylkens, B.; Thiry, J.; Kirten, P.; Schynts, F.; Thiry, E. Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet. Res. 2007, 38, 181–209. [Google Scholar] [CrossRef] [Green Version]
- Andrei, G.; De Clercq, E.; Snoeck, R. Novel inhibitors of human CMV. Curr. Opin. Investig. Drugs 2008, 9, 132–145. [Google Scholar]
- De Clercq, E. A cutting-edge view on the current state of antiviral drug development. Med. Res. Rev. 2013, 33, 1249–1277. [Google Scholar] [CrossRef]
- Alvisi, G.; Jans, D.A.; Camozzi, D.; Avanzi, S.; Loregian, A.; Ripalti, A.; Palu, G. Regulated transport into the nucleus of herpesviridae DNA replication core proteins. Viruses 2013, 5, 2210–2234. [Google Scholar] [CrossRef] [Green Version]
- Pari, G.S.; Anders, D.G. Eleven loci encoding trans-acting factors are required for transient complementation of human cytomegalovirus oriLyt-dependent DNA replication. J. Virol. 1993, 67, 6979–6988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlich, D.; Kostka, S.; Kraft, R.; Dingwall, C.; Laskey, R.A.; Hartmann, E.; Prehn, S. Two different subunits of importin cooperate to recognize nuclear localization signals and bind them to the nuclear envelope. Curr. Biol. 1995, 5, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Tran, E.J.; Bolger, T.A.; Wente, S.R. SnapShot: Nuclear transport. Cell 2007, 131, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalderon, D.; Richardson, W.D.; Markham, A.F.; Smith, A.E. Sequence requirements for nuclear location of simian virus 40 large-T antigen. Nature 1984, 311, 33–38. [Google Scholar] [CrossRef]
- Cansizoglu, A.E.; Lee, B.J.; Zhang, Z.C.; Fontoura, B.M.; Chook, Y.M. Structure-based design of a pathway-specific nuclear import inhibitor. Nat. Struct. Mol. Biol. 2007, 14, 452–454. [Google Scholar] [CrossRef]
- Alvisi, G.; Jans, D.; Guo, J.; Pinna, L.; Ripalti, A. A protein kinase CK2 site flanking the nuclear targeting signal enhances nuclear transport of human cytomegalovirus ppUL44. Traffic 2005, 6, 1002–1013. [Google Scholar] [CrossRef] [Green Version]
- Alvisi, G.; Ripalti, A.; Ngankeu, A.; Giannandrea, M.; Caraffi, S.G.; Dias, M.M.; Jans, D.A. Human cytomegalovirus DNA polymerase catalytic subunit pUL54 possesses independently acting nuclear localization and ppUL44 binding motifs. Traffic 2006, 7, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Alvisi, G.; Musiani, D.; Jans, D.A.; Ripalti, A. An importin alpha/beta-recognized bipartite nuclear localization signal mediates targeting of the human herpes simplex virus type 1 DNA polymerase catalytic subunit pUL30 to the nucleus. Biochemistry 2007, 46, 9155–9163. [Google Scholar] [CrossRef]
- Alvisi, G.; Avanzi, S.; Musiani, D.; Camozzi, D.; Leoni, V.; Ly-Huynh, J.D.; Ripalti, A. Nuclear import of HSV-1 DNA polymerase processivity factor UL42 is mediated by a C-terminally located bipartite nuclear localization signal. Biochemistry 2008, 47, 13764–13777. [Google Scholar] [CrossRef]
- Chen, Y.; Ciustea, M.; Ricciardi, R.P. Processivity factor of KSHV contains a nuclear localization signal and binding domains for transporting viral DNA polymerase into the nucleus. Virology 2005, 340, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, D.; Kanda, T.; Murata, T.; Saito, S.; Sugimoto, A.; Narita, Y.; Tsurumi, T. Nuclear Transport of Epstein-Barr Virus DNA Polymerase is dependent on the BMRF1 Polymerase Processivity Factor and Molecular Chaperone Hsp90. J. Virol. 2013, 87, 6482–6491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, C.; Liu, W.; Wang, B.; Dang, R.; Qiu, L.; Ren, J.; Yan, C.; Yang, Z.; Wang, X. Ivermectin inhibits DNA polymerase UL42 of pseudorabies virus entrance into the nucleus and proliferation of the virus in vitro and vivo. Antivir. Res. 2018, 159, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.N.Y.; Atkinson, S.C.; Wang, C.; Lee, A.; Bogoyevitch, M.A.; Borg, N.A.; Jans, D.A. The broad spectrum antiviral ivermectin targets the host nuclear transport importin alpha/beta1 heterodimer. Antivir. Res. 2020, 104760. [Google Scholar] [CrossRef] [PubMed]
- Alvisi, G.; Roth, D.M.; Camozzi, D.; Pari, G.S.; Loregian, A.; Ripalti, A.; Jans, D.A. The flexible loop of the human cytomegalovirus DNA polymerase processivity factor ppUL44 is required for efficient DNA binding and replication in cells. J. Virol. 2009, 83, 9567–9576. [Google Scholar] [CrossRef] [Green Version]
- Alvisi, G.; Marin, O.; Pari, G.; Mancini, M.; Avanzi, S.; Loregian, A.; Jans, D.A.; Ripalti, A. Multiple phosphorylation sites at the C-terminus regulate nuclear import of HCMV DNA polymerase processivity factor ppUL44. Virology 2011, 417, 259–267. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six classes of nuclear localization signals specific to different binding grooves of importin alpha. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Alvisi, G.; Paolini, L.; Contarini, A.; Zambarda, C.; Di Antonio, V.; Colosini, A.; Mercandelli, N.; Timmoneri, M.; Palu, G.; Caimi, L.; et al. Intersectin goes nuclear: Secret life of an endocytic protein. Biochem. J. 2018, 475, 1455–1472. [Google Scholar] [CrossRef] [Green Version]
- Shahin, F.; Raza, S.; Yang, K.; Hu, C.; Chen, Y.; Chen, H.; Guo, A. Bovine herpesvirus 1 tegument protein UL21 plays critical roles in viral secondary envelopment and cell-to-cell spreading. Oncotarget 2017, 8, 94462–94480. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, A.; Prince, D.; Lo, C.Y.; Lee, L.H.; Chu, T.C. Antiviral activity of theaflavin digallate against herpes simplex virus type 1. Antivir. Res. 2015, 118, 56–67. [Google Scholar] [CrossRef]
- Cantatore, A.; Randall, S.D.; Traum, D.; Adams, S.D. Effect of black tea extract on herpes simplex virus-1 infection of cultured cells. BMC Complement.Altern. Med. 2013, 13, 139. [Google Scholar] [CrossRef] [Green Version]
- Levings, R.L.; Roth, J.A. Immunity to bovine herpesvirus 1: I. Viral lifecycle and innate immunity. Anim. Health Res. Rev. 2013, 14, 88–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.L.; Han, Y.; Liu, W.; Pang, X.Y.; Zheng, B.; Zhang, Y.; Zhou, X.N. Antivirus effectiveness of ivermectin on dengue virus type 2 in Aedes albopictus. PLoS Negl. Trop. Dis. 2018, 12, e0006934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frizzoda Silva, L.; Kook, I.; Doster, A.; Jones, C. Bovine herpesvirus 1 regulatory proteins bICP0 and VP16 are readily detected in trigeminal ganglionic neurons expressing the glucocorticoid receptor during the early stages of reactivation from latency. J. Virol. 2013, 87, 11214–11222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottlieb, J.; Marcy, A.I.; Coen, D.M.; Challberg, M.D. The Herpes-Simplex Virus Type-1 Ul42 Gene-Product—A Subunit of DNA-Polymerase That Functions to Increase Processivity. J. Virol. 1990, 64, 5976–5987. [Google Scholar] [CrossRef] [Green Version]
- Van Voorhis, W.C.; Hooft van Huijsduijnen, R.; Wells, T.N. Profile of William C. Campbell, Satoshi Omura, and Youyou Tu, 2015 Nobel Laureates in Physiology or Medicine. Proc. Natl. Acad. Sci. USA 2015, 112, 15773–15776. [Google Scholar] [CrossRef] [Green Version]
- Trailovic, S.M.; Ivanovic, S.R.; Varagic, V.M. Ivermectin effects on motor coordination and contractions of isolated rat diaphragm. Res. Vet. Sci. 2011, 91, 426–433. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin alpha/beta-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef] [Green Version]
- Barrows, N.J.; Campos, R.K.; Powell, S.T.; Prasanth, K.R.; Schott-Lerner, G.; Soto-Acosta, R.; Galarza-Munoz, G.; McGrath, E.L.; Urrabaz-Garza, R.; Gao, J.; et al. A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell Host Microb. 2016, 20, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Mastrangelo, E.; Pezzullo, M.; De Burghgraeve, T.; Kaptein, S.; Pastorino, B.; Dallmeier, K.; de Lamballerie, X.; Neyts, J.; Hanson, A.M.; Frick, D.N.; et al. Ivermectin is a potent inhibitor of flavivirus replication specifically targeting NS3 helicase activity: New prospects for an old drug. J. Antimicrob. Chemother. 2012, 67, 1884–1894. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Fu, X.; Huang, L.; Ma, Y.; Ding, X.; Zhu, L.; Zhu, G. The synergistic antiviral effects of GSH in combination with acyclovir against BoHV-1 infection in vitro. Acta Virol. 2016, 60, 328–332. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, S.; Shahin, F.; Zhai, W.; Li, H.; Alvisi, G.; Yang, K.; Chen, X.; Chen, Y.; Chen, J.; Hu, C.; et al. Ivermectin Inhibits Bovine Herpesvirus 1 DNA Polymerase Nuclear Import and Interferes With Viral Replication. Microorganisms 2020, 8, 409. https://doi.org/10.3390/microorganisms8030409
Raza S, Shahin F, Zhai W, Li H, Alvisi G, Yang K, Chen X, Chen Y, Chen J, Hu C, et al. Ivermectin Inhibits Bovine Herpesvirus 1 DNA Polymerase Nuclear Import and Interferes With Viral Replication. Microorganisms. 2020; 8(3):409. https://doi.org/10.3390/microorganisms8030409
Chicago/Turabian StyleRaza, Sohail, Farzana Shahin, Wenjun Zhai, Hanxiong Li, Gualtiero Alvisi, Kui Yang, Xi Chen, Yingyu Chen, Jianguo Chen, Changmin Hu, and et al. 2020. "Ivermectin Inhibits Bovine Herpesvirus 1 DNA Polymerase Nuclear Import and Interferes With Viral Replication" Microorganisms 8, no. 3: 409. https://doi.org/10.3390/microorganisms8030409