RHΔgra17Δnpt1 Strain of Toxoplasma gondii Elicits Protective Immunity Against Acute, Chronic and Congenital Toxoplasmosis in Mice

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Parasites
2.3. Engineering of RHΔgra17Δnpt1 Mutant Strains
2.4. Optimization of the Vaccination Dose
2.5. Immune Responses Induced by Immunization with RHΔgra17Δnpt1
2.6. Protection of Mice Against Acute and Chronic Infection
2.7. Protection Against Congenital Transmission
2.8. Statistical Analysis
3. Results
3.1. Construction of RHΔgra17Δnpt1 Strain
3.2. Characterization and Evaluation of RHΔgra17Δnpt1 Attenuation
3.3. Immune Responses Induced by Vaccination
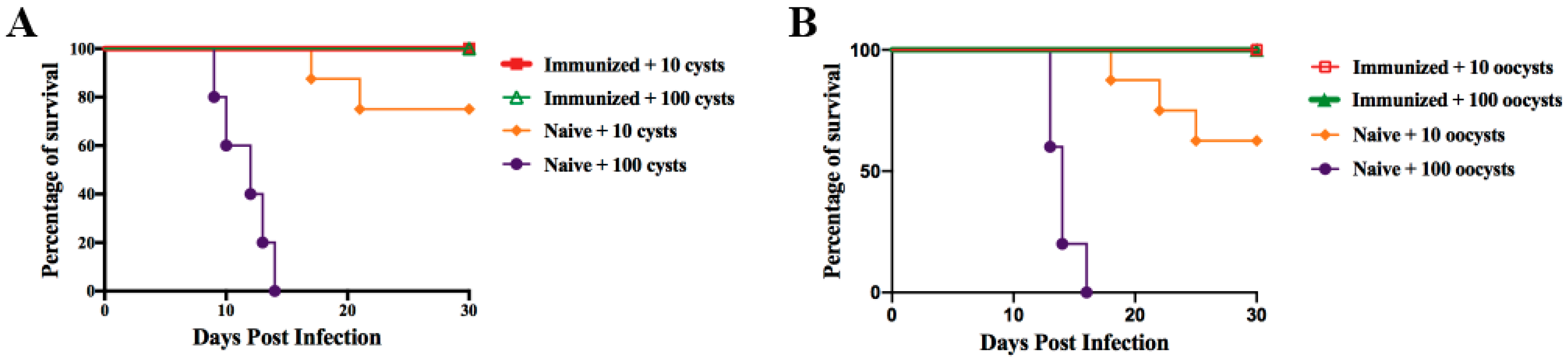
3.4. Protection Against Acute Infection
3.5. Protection against Chronic Infection
3.6. Protection Against Congenital Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, E.; Pollak, A.; Reiterowona, I. Recent trends in research on congenital toxoplasmosis. Int. J. Parasitol. 2001, 31, 115–144. [Google Scholar] [CrossRef]
- Robert-Gangneux, F. It is not only the cat that did it: How to prevent and treat congenital toxoplasmosis. J. Infect. 2014, 68, S125–S133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.D.; Wang, S.C.; Liu, H.H.; Ma, H.Y.; Li, Z.Y.; Wei, F.; Zhu, X.Q.; Liu, Q. Prevalence and burden of Toxoplasma gondii infection in HIV-infected people: A systematic review and meta-analysis. Lancet HIV 2017, 4, e177–e188. [Google Scholar] [CrossRef]
- Sensini, A. Toxoplasma gondii infection in pregnancy: Opportunities and pitfalls of serological diagnosis. Clin. Microbiol. Infect. 2010, 12, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Holland, A.; Joseph, D. Drugs in development for toxoplasmosis: Advances, challenges, and current status. Drug Des. Dev. Ther. 2017, 11, 273–293. [Google Scholar]
- Wang, J.L.; Zhang, N.Z.; Li, T.T.; He, J.J.; Elsheikha, H.M.; Zhu, X.Q. Advances in the development of Anti-Toxoplasma gondii vaccines: Challenges, opportunities, and perspectives. Trends Parasitol. 2019, 35, 239–253. [Google Scholar] [CrossRef]
- Buxton, D.; Tomson, K.; Maley, S.; Wright, S.; Bos, H.J. Vaccination of sheep with a live incomplete strain (S48) of Toxoplasma gondii and their immunity to challenge when pregnant. Vet. Rec. 1991, 129, 89–93. [Google Scholar] [CrossRef]
- Hiszczyńska-Sawicka, E.; Gatkowska, J.M.; Grzybowski, M.M.; Długońska, H. Veterinary vaccines against toxoplasmosis. Parasitology 2014, 141, 1365–1378. [Google Scholar] [CrossRef]
- Zhang, N.Z.; Wang, M.; Xu, Y.; Petersen, E.; Zhu, X.Q. Recent advances in developing vaccines against Toxoplasma gondii: An update. Expert Rev. Vaccines 2015, 14, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Buxton, D.; Innes, E.A. A commercial vaccine for ovine toxoplasmosis. Parasitology 1995, 110 (Suppl. 1), S11–S16. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Elsheikha, H.M.; Zhu, W.N.; Chen, K.; Li, T.T.; Yue, D.M.; Zhang, X.X.; Huang, S.Y.; Zhu, X.Q. Immunization with Toxoplasma gondii GRA17 deletion mutant induces partial protection and survival in challenged mice. Front. Immunol. 2017, 8, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Chen, K.; Cong, W.; Yang, W.B.; Bai, M.J.; Huang, S.Y.; Zhu, X.Q. Live attenuated Pru: Δcdpk2 strain of Toxoplasma gondii protects against acute, chronic, and congenital toxoplasmosis. J. Infect. Dis. 2018, 218, 768–777. [Google Scholar] [CrossRef] [Green Version]
- Gold, D.A.; Kaplan, A.D.; Lis, A.; Bett, G.C.; Rosowski, E.E.; Cirelli, K.M.; Bougdour, A.; Sidik, S.M.; Beck, J.R.; Lourido, S.; et al. The Toxoplasma dense granule proteins GRA17 and GRA23 mediate the movement of small molecules between the host and the parasitophorous vacuole. Cell Host Microbe 2015, 17, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Mercier, C.; Cesbron-Delauw, M.F. Toxoplasma secretory granules: One population or more? Trends Parasitol. 2015, 31, 60–71. [Google Scholar] [CrossRef]
- Rajendran, E.; Hapuarachchi, S.V.; Miller, C.M.; Fairweather, S.J.; Cai, Y.; Smith, N.C.; Cockburn, I.A.; Bröer, S.; Kirk, K.; van Dooren, G.G.; et al. Cationic amino acid transporters play key roles in the survival and transmission of apicomplexan parasites. Nat. Commun. 2017, 8, 14455. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Liu, K.; Yang, L.; Xia, G.L.; Tan, Q.Y.; Wang, G. Measurement and analysis of the reproductive performance in six commonly used SPF mice and rats. Chin. J. Comp. Med. 2016, 10, 9–13. [Google Scholar]
- Roos, D.S.; Donald, R.G.; Morrissette, N.S.; Moulton, A.L. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. Methods Cell Biol. 1994, 45, 27–63. [Google Scholar]
- Hu, R.S.; He, J.J.; Elsheikha, H.M.; Zhang, F.K.; Zou, Y.; Zhao, G.H.; Cong, W.; Zhu, X.Q. Differential brain MicroRNA expression profiles after acute and chronic infection of mice with Toxoplasma gondii oocysts. Front. Microbiol. 2018, 9, 2316. [Google Scholar] [CrossRef] [Green Version]
- Staggs, S.E.; See, M.J.; Dubey, J.P.; Villegas, E.N. Obtaining highly purified Toxoplasma gondii oocysts by a discontinuous cesium chloride gradient. J. Vis. Exp. 2009, 33, e1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.L.; Bai, M.J.; Elsheikha, H.M.; Liang, Q.L.; Li, T.T.; Cao, X.Z.; Zhu, X.Q. Novel roles of dense granule protein 12 (GRA12) in Toxoplasma gondii infection. FASEB J. 2020, 34, 3165–3178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shwab, E.K.; Saraf, P.; Zhu, X.; Zhou, D.H.; Mcferrin, B.M.; Ajzenberg, D.; Schares, G.; Hammond-Aryee, K.; van Helden, P.; Higgins, S.A.; et al. Human impact on the diversity and virulence of the ubiquitous zoonotic parasite Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 2018, 115, E6956–E6963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.B.; Wang, J.L.; Gui, Q.; Zou, Y.; Chen, K.; Liu, Q.; Liang, Q.L.; Zhu, X.Q.; Zhou, D.H. Immunization with a Live-attenuated RH:ΔNPT1 strain of Toxoplasma gondii induces strong protective immunity against Toxoplasmosis in mice. Front. Microbiol. 2019, 10, 1875. [Google Scholar] [CrossRef] [Green Version]
- Gigley, J.P.; Fox, B.A.; Bzik, D.J. Cell-mediated immunity to Toxoplasma gondii develops primarily by local th1 host immune responses in the absence of parasite replication. J. Immunol. 2009, 182, 1069–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayles, P.C.; Gibson, G.W.; Johnson, L.L. B cells are essential for vaccination-induced resistance to virulent Toxoplasma gondii. Infect. Immun. 2000, 68, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.A.; Sibley, L.D. Modulation of innate immunity by Toxoplasma gondii virulence effectors. Nat. Rev. Microbiol. 2012, 10, 766–778. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, D.L.; Fatima, R.; Gigley, J.P. Comparative analysis of conventional natural killer cell responses to acute infection with Toxoplasma gondii strains of different virulence. Front. Immunol. 2016, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Gazzinelli, R.T.; Wysocka, M.; Hieny, S.; Scharton-Kersten, T.; Cheever, A.; Kuhn, R.; Müller, W.; Trinchieri, G.; Sher, A. In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4+ T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha. J. Immunol. 1996, 157, 798–805. [Google Scholar]
- Group, S.S.; Thiébaut, R.; Leproust, S.; Chêne, G.; Gilbert, R. Effectiveness of prenatal treatment for congenital toxoplasmosis: A meta-analysis of individual patients’ data. Lancet 2007, 369, 115–122. [Google Scholar]
- Zhang, Y.H.; He, M.; Wang, Y.; Liao, A.H. Modulators of the balance between M1 and M2 macrophages during pregnancy. Front. Immunol. 2017, 8, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lao, K.; Zhao, M.; Li, Z.; Liu, X.; Zhang, H.; Jiang, Y.; Wang, Y.; Hu, X. IL-10 regulate decidual Tregs apoptosis contributing to the abnormal pregnancy with Toxoplasma gondii infection. Microb. Pathog. 2015, 89, 210–216. [Google Scholar] [CrossRef] [PubMed]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Q.-L.; Sun, L.-X.; Elsheikha, H.M.; Cao, X.-Z.; Nie, L.-B.; Li, T.-T.; Li, T.-S.; Zhu, X.-Q.; Wang, J.-L. RHΔgra17Δnpt1 Strain of Toxoplasma gondii Elicits Protective Immunity Against Acute, Chronic and Congenital Toxoplasmosis in Mice. Microorganisms 2020, 8, 352. https://doi.org/10.3390/microorganisms8030352
Liang Q-L, Sun L-X, Elsheikha HM, Cao X-Z, Nie L-B, Li T-T, Li T-S, Zhu X-Q, Wang J-L. RHΔgra17Δnpt1 Strain of Toxoplasma gondii Elicits Protective Immunity Against Acute, Chronic and Congenital Toxoplasmosis in Mice. Microorganisms. 2020; 8(3):352. https://doi.org/10.3390/microorganisms8030352
Chicago/Turabian StyleLiang, Qin-Li, Li-Xiu Sun, Hany M. Elsheikha, Xue-Zhen Cao, Lan-Bi Nie, Ting-Ting Li, Tao-Shan Li, Xing-Quan Zhu, and Jin-Lei Wang. 2020. "RHΔgra17Δnpt1 Strain of Toxoplasma gondii Elicits Protective Immunity Against Acute, Chronic and Congenital Toxoplasmosis in Mice" Microorganisms 8, no. 3: 352. https://doi.org/10.3390/microorganisms8030352