The Control of Intestinal Inflammation: A Major Objective in the Research of Probiotic Strains as Alternatives to Antibiotic Growth Promoters in Poultry



Abstract
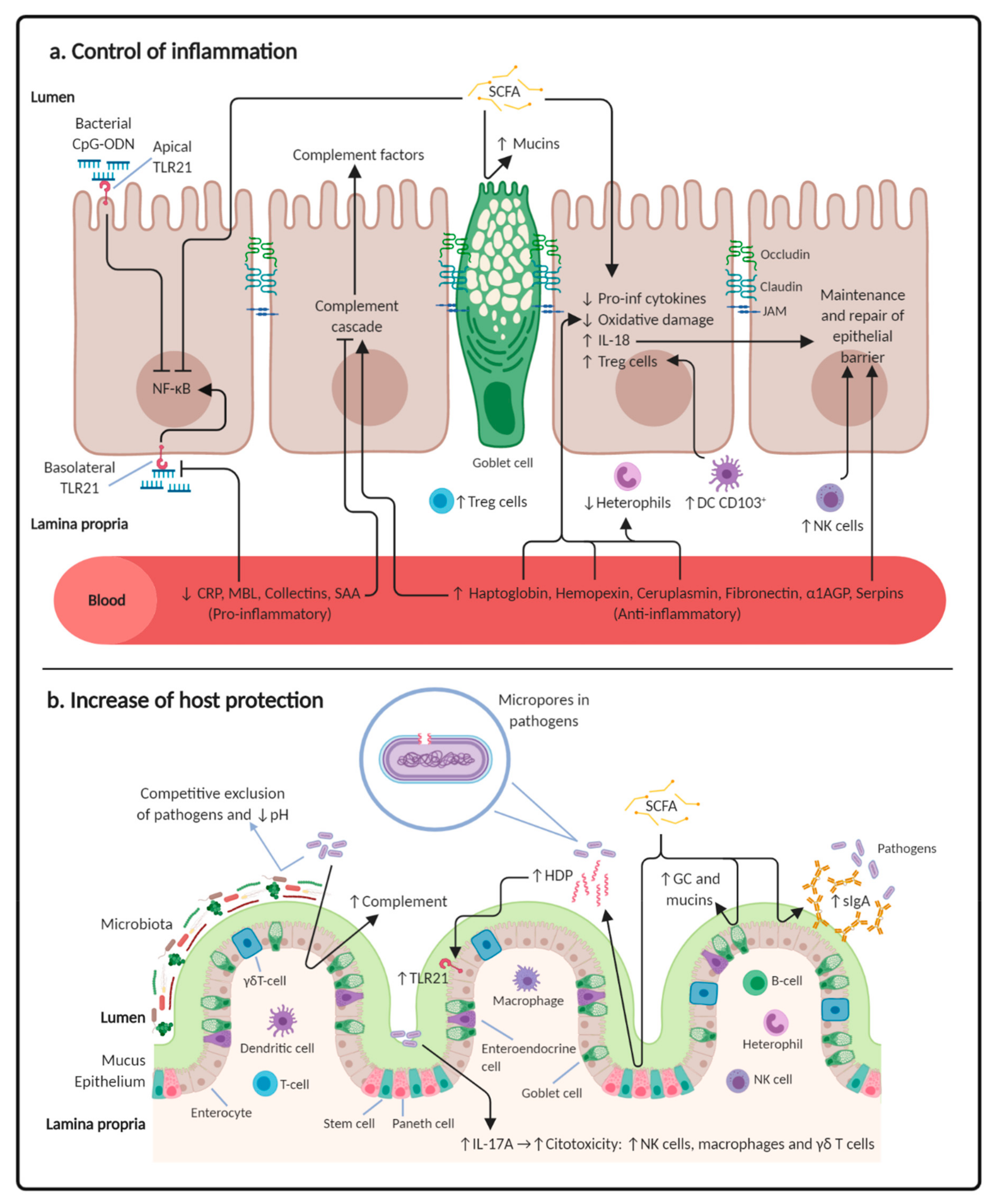
:1. Introduction
1.1. The Threats
1.2. The Role of Inflammation
1.3. The Role of Probiotics on the Control of Inflammation
2. Effects of Probiotics on the Immune System
2.1. Effects of Probiotics on Acute-Phase Response
2.2. Effects of Probiotics on Complement
2.3. Effects of Probiotics on TLR Repertoire
2.4. Effects of Probiotics on Cells of the Innate Immune System
2.5. Effects of Probiotics on Antimicrobial Peptides (AMPs) Production
3. Conclusions
Funding
Conflicts of Interest
References
- FAO. Food Outlook: Biannual Report on Global Food Markets. In Biannual report on global food markets 2017; FAO: Rome, Italy, 2017; p. 152. [Google Scholar]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. In The Review on Antimicrobial Resistance 2016; HM Government and Wellcome Trust: London, UK, 2016. [Google Scholar]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Regulation, O. No 1831/2003 of the European Parliament and Council of 22 September 2003 on additives for use in animal nutrition. Off. J. Eur. Commun. 2003, 268, 29–43. [Google Scholar]
- Wierup, M. The control of microbial diseases in animals: Alternatives to the use of antibiotics. Int. J. Antimicrob. Agents 2000, 14, 315–319. [Google Scholar] [CrossRef]
- Dahiya, J.; Wilkie, D.; Van Kessel, A.; Drew, M. Potential strategies for controlling necrotic enteritis in broiler chickens in post-antibiotic era. Anim. Feed. Sci. Technol. 2006, 129, 60–88. [Google Scholar] [CrossRef]
- Choct, M. Alternatives to in-feed antibiotics in monogastric animal industry. ASA Tech. Bull. 2001, 30, 1–6. [Google Scholar]
- Hofacre, C.L.; Smith, J.A.; Mathis, G.F. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate. Poult. Sci. 2018, 97, 1929–1933. [Google Scholar] [CrossRef]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, U.; et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Heal. Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Huyghebaert, G.; Ducatelle, R.; Van Immerseel, F. An update on alternatives to antimicrobial growth promoters for broilers. Vet. J. 2011, 187, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Hunkapiller, A.A.; Layton, A.C.; Chang, Y.-J.; Robbins, K.R. Response of Intestinal Microbiota to Antibiotic Growth Promoters in Chickens. Foodborne Pathog. Dis. 2013, 10, 331–337. [Google Scholar] [CrossRef]
- Cho, I.; Yamanishi, S.; Cox, L.; Methe, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, U.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Seal, B.S.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal production. Anim. Heal. Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celi, P.; Verlhac, V.; Calvo, E.P.; Schmeisser, J.; Kluenter, A.-M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed. Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Ducatelle, R.; Goossens, E.; De Meyer, F.; Eeckhaut, V.; Antonissen, G.; Haesebrouck, F.; Van Immerseel, F. Biomarkers for monitoring intestinal health in poultry: Present status and future perspectives. Vet. Res. 2018, 49, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. BioMed Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tarradas, J.; Esteve-Garcia, E.; Brufau, J. Review of immune stimulator substances/agents that are susceptible of being used as feed additives: Mode of action and identification of end-points for efficacy assessment. EFSA Support. Publ. 2015, 12, 905E. [Google Scholar]
- Al-Khalaifah, H.S. Benefits of probiotics and/or prebiotics for antibiotic-reduced poultry. Poult. Sci. 2018, 97, 3807–3815. [Google Scholar] [CrossRef]
- Ma, T.; Suzuki, Y.; Guan, L.L. Dissect the mode of action of probiotics in affecting host-microbial interactions and immunity in food producing animals. Vet. Immunol. Immunopathol. 2018, 205, 35–48. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. The role of the gut microbiome in shaping the immune system of chickens. Vet. Immunol. Immunopathol. 2018, 204, 44–51. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. Gut immunity: Its development and reasons and opportunities for modulation in monogastric production animals. Anim. Heal. Res. Rev. 2018, 19, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend or foe for animal production? Poult. Sci. 2018, 97, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H.; Genovese, K.J.; Swaggerty, C.L.; He, H.; Broom, L. Inflammatory phenotypes in the intestine of poultry: Not all inflammation is created equal. Poult. Sci. 2018, 97, 2339–2346. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J.; Blumberg, R.S. The immunology of mucosal models of inflammation. Annu. Rev. Immunol. 2002, 20, 495–549. [Google Scholar] [CrossRef]
- Smith, A.L.; Powers, C.; Beal, R.K. Chapter 13—The Avian Enteric Immune System in Health and Disease. In Avian Immunology, 2nd ed.; Academic Press: Boston, MA, USA, 2014; pp. 227–250. [Google Scholar] [CrossRef]
- Berghman, L.R. Immune responses to improving welfare. Poult. Sci. 2016, 95, 2216–2218. [Google Scholar] [CrossRef]
- Iseri, V.J.; Klasing, K.C. Changes in the Amount of Lysine in Protective Proteins and Immune Cells after a Systemic Response to Dead Escherichia coli: Implications for the Nutritional Costs of Immunity. Integr. Comp. Boil. 2014, 54, 922–930. [Google Scholar] [CrossRef]
- Klasing, K. Nutrition and the immune system. Br. Poult. Sci. 2007, 48, 525–537. [Google Scholar] [CrossRef]
- Shattuck, E.C.; Muehlenbein, M.P. Human sickness behavior: Ultimate and proximate explanations. Am. J. Phys. Anthr. 2015, 157, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Velová, H.; Gutowska-Ding, M.W.; Burt, D.W.; Vinkler, M. Toll-Like Receptor Evolution in Birds: Gene Duplication, Pseudogenization, and Diversifying Selection. Mol. Boil. Evol. 2018, 35, 2170–2184. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Aso, H.; Kitazawa, H. Regulation of Toll-Like Receptors-Mediated Inflammation by Immunobiotics in Bovine Intestinal Epitheliocytes: Role of Signaling Pathways and Negative Regulators. Front. Immunol. 2014, 5, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Seifi, K.; Karimi-Torshizi, M.A.; Deldar, H. Probiotics intake from proximal or distal gastrointestinal tract: The investigation on intestinal morphology and performance of Japanese quail. J. Anim. Physiol. Anim. Nutr. 2017, 102, e509–e512. [Google Scholar] [CrossRef] [Green Version]
- Brufau, M.T.; Martín-Venegas, R.; Guerrero-Zamora, A.M.; Pérez-Vendrell, A.M.; Vilà, B.; Ferrer, R. Dietary β-galactomannans have beneficial effects on the intestinal morphology of chickens challenged with Salmonella enterica serovar Enteritidis1. J. Anim. Sci. 2015, 93, 238–246. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Zhou, Z.; Packialakshmi, B.; Makkar, S.; Dridi, S.; Rath, N. Effect of butyrate on immune response of a chicken macrophage cell line. Vet. Immunol. Immunopathol. 2014, 162, 24–32. [Google Scholar] [CrossRef]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y.; Kotani, J. Butyrate and trichostatin A attenuate nuclear factor κB activation and tumor necrosis factor α secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. 2008, 28, 321–328. [Google Scholar] [CrossRef]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.I.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Guo, B.; Gan, Z.; Song, D.; Lu, Z.; Yi, H.; Wu, Y.; Wang, Y.; Du, H. Butyrate upregulates endogenous host defense peptides to enhance disease resistance in piglets via histone deacetylase inhibition. Sci. Rep. 2016, 6, 27070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigley, P. Salmonella enterica in the Chicken: How it has Helped Our Understanding of Immunology in a Non-Biomedical Model Species. Front. Immunol. 2014, 5, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juul-Madsen, H.R.; Viertlböeck, B.; Härtle, S.; Smith, A.L.; Göbel, T.W. Chapter 7—Innate Immune Responses. In Avian Immunology, 2nd ed.; Schat, K.A., Kaspers, B., Kaiser, P., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 121–147. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, I.H. The effects of the supplementation of Bacillus subtilis RX7 and B2A strains on the performance, blood profiles, intestinal Salmonella concentration, noxious gas emission, organ weight and breast meat quality of broiler challenged with Salmonella typhimuri. J. Anim. Physiol. Anim. Nutr. 2014, 99, 326–334. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, I.H. Supplemental effect of probioticBacillus subtilis B2A on productivity, organ weight, intestinal Salmonella microflora, and breast meat quality of growing broiler chicks. Poult. Sci. 2014, 93, 2054–2059. [Google Scholar] [CrossRef]
- Murata, H.; Shimada, N.; Yoshioka, M. Current research on acute phase proteins in veterinary diagnosis: An overview. Vet. J. 2004, 168, 28–40. [Google Scholar] [CrossRef]
- Bielecka, M.; Smoragiewicz, W.; Siwicki, A.K.; Wójcik, R.; Biedrzycka, E.; Orłowski, A.; Kask, S.; Jankowski, J.; Karska-Wysocki, B.; Ham, D. The Effect of Various Probiotic Strains or Avilamycin Feed Additive on Immune Defense Markers and Acute-Phase Response to Salmonella Infection in Chickens. Probiotics Antimicrob. Proteins 2010, 2, 175–185. [Google Scholar] [CrossRef]
- Pepys, M.B.; Hirschfield, G.M. C-reactive protein: A critical update. J. Clin. Invest. 2003, 111, 1805–1812. [Google Scholar] [CrossRef]
- Thompson, D.; Pepys, M.B.; Wood, S.P. The physiological structure of human C-reactive protein and its complex with phosphocholine. Structure 1999, 7, 169–177. [Google Scholar] [CrossRef]
- Sohail, M.U.; Ijaz, A.; Yousaf, M.S.; Ashraf, K.; Zaneb, H.; Aleem, M.; Rehman, H. Alleviation of cyclic heat stress in broilers by dietary supplementation of mannan-oligosaccharide and Lactobacillus-based probiotic: Dynamics of cortisol, thyroid hormones, cholesterol, C-reactive protein, and humoral immunity. Poult. Sci. 2010, 89, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Garred, P.; Honoré, C.; Ma, Y.J.; Munthe-Fog, L.; Hummelshøj, T. MBL2, FCN1, FCN2 and FCN3—The genes behind the initiation of the lectin pathway of complement. Mol. Immunol. 2009, 46, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Eddie Ip, W.K.; Takahashi, K.; Ezekowitz, R.A.; Stuart, L.M. Mannose-binding lectin and innate immunity. Immunol. Rev. 2009, 230, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Lynge, S.L.; Juul-Madsen, H.R.; Kjærup, R.B.; Okimoto, R.; Abrahamsen, M.S.; Maurischat, S.; Sørensen, P.; Dalgaard, T.S. Broilers with low serum Mannose-binding Lectin show increased fecal shedding of Salmonella enterica serovar Montevideo. Poult. Sci. 2016, 95, 1779–1786. [Google Scholar] [CrossRef]
- Tang, Y.; Ma, D.; Ming, S.; Zhang, L.; Zhou, J.; Shan, G.; Chen, Z.; Lu, X.; Zuo, D. Mannan-binding lectin reduces CpG DNA-induced inflammatory cytokine production by human monocytes. Microbiol. Immunol. 2015, 59, 231–237. [Google Scholar] [CrossRef]
- Kant, R.; de Vos, W.M.; Palva, A.; Satokari, R. Immunostimulatory CpG motifs in the genomes of gut bacteria and their role in human health and disease. J. Med. Microbiol. 2014, 63, 293–308. [Google Scholar] [CrossRef]
- Kopp, Z.A.; Jain, U.; Van Limbergen, J.; Stadnyk, A.W. Do Antimicrobial Peptides and Complement Collaborate in the Intestinal Mucosa? Front. Immunol. 2015, 6, 17. [Google Scholar] [CrossRef]
- Kobayashi, H.; Albarracin, L.; Sato, N.; Kanmani, P.; Kober, A.; Ikeda-Ohtsubo, W.; Suda, Y.; Nochi, T.; Aso, H.; Makino, S.; et al. Modulation of porcine intestinal epitheliocytes immunetranscriptome response by Lactobacillus jensenii TL2937. Benef. Microbes 2016, 7, 769–782. [Google Scholar] [CrossRef]
- Sánchez, B.; González-Rodríguez, I.; Arboleya, S.; Lopez, P.; Suárez, A.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M. The Effects of Bifidobacterium breve on Immune Mediators and Proteome of HT29 Cells Monolayers. BioMed Res. Int. 2015, 2015, 1–6. [Google Scholar]
- Zhan, H.; Dong, X.; Li, L.; Zheng, Y.; Gong, Y.; Zou, X. Effects of dietary supplementation with Clostridium butyricum on laying performance, egg quality, serum parameters, and cecal microflora of laying hens in the late phase of production. Poult. Sci. 2019, 98, 896–903. [Google Scholar] [CrossRef]
- De Kivit, S.; Tobin, M.C.; Forsyth, C.B.; Keshavarzian, A.; Landay, A.L. Regulation of Intestinal Immune Responses through TLR Activation: Implications for Pro- and Prebiotics. Front. Immunol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaide, M.; Edwards, S.V. Molecular Evolution of the Toll-Like Receptor Multigene Family in Birds. Mol. Boil. Evol. 2011, 28, 1703–1715. [Google Scholar] [CrossRef] [Green Version]
- Keestra, A.M.; De Zoete, M.R.; Bouwman, L.I.; Vaezirad, M.M.; Van Putten, J.P. Unique features of chicken Toll-like receptors. Dev. Comp. Immunol. 2013, 41, 316–323. [Google Scholar] [CrossRef]
- Keestra, A.M.; De Zoete, M.R.; Van Aubel, R.A.M.H.; Van Putten, J.P.M. The central leucine-rich repeat region of chicken TLR16 dictates unique ligand specificity and species-specific interaction with TLR2. J. Immunol. 2007, 178, 7110–7119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, M.; Matsuo, A.; Shingai, M.; Shida, K.; Ishii, A.; Funami, K.; Suzuki, Y.; Oshiumi, H.; Matsumoto, M.; Seya, T. Combinational recognition of bacterial lipoproteins and peptidoglycan by chicken Toll-like receptor 2 subfamily. Dev. Comp. Immunol. 2008, 32, 147–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownlie, R.; Allan, B. Avian toll-like receptors. Cell Tissue Res. 2011, 343, 121–130. [Google Scholar] [CrossRef]
- Keestra, A.M.; De Zoete, M.R.; Bouwman, L.I.; Van Putten, J.P.M. Chicken TLR21 Is an Innate CpG DNA Receptor Distinct from Mammalian TLR9. J. Immunol. 2010, 185, 460–467. [Google Scholar] [CrossRef] [Green Version]
- De Zoete, M.R.; Bouwman, L.I.; Keestra, A.M.; Van Putten, J.P.M. Cleavage and activation of a Toll-like receptor by microbial proteases. Proc. Natl. Acad. Sci. USA 2011, 108, 4968–4973. [Google Scholar] [CrossRef] [Green Version]
- Kannaki, T.R.; Reddy, M.R.; Verma, P.C.; Shanmugam, M. Differential Toll-like receptor (TLR) mRNA expression patterns during chicken embryological development. Anim. Biotechnol. 2015, 26, 130–135. [Google Scholar] [CrossRef]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–144. [Google Scholar] [CrossRef]
- Bai, S.P.; Wu, A.M.; Ding, X.M.; Lei, Y.; Bai, J.; Zhang, K.Y.; Chio, J.S. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 2013, 92, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, P.; Lee, C.H.; Cosby, D.E.; Cox, N.A.; Kim, W.K. Effect of probiotics on fecal excretion, colonization in internal organs and immune gene expression in the ileum of laying hens challenged with SalmonellaEnteritidis. Poult. Sci. 2019, 98, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Yitbarek, A.; Echeverry, H.; Munyaka, P.; Rodriguez-Lecompte, J.C. Innate immune response of pullets fed diets supplemented with prebiotics and synbiotics. Poult. Sci. 2015, 94, 1802–1811. [Google Scholar] [CrossRef] [PubMed]
- Karaffova, V.; Marcinkova, E.; Bobikova, K.; Herich, R.; Revajova, V.; Stasova, D.; Kavulova, A.; Levkutova, M.; Levkut, M., Jr.; Laukova, A.; et al. TLR4 and TLR21 expression, MIF, IFN-beta, MD-2, CD14 activation, and sIgA production in chickens administered with EFAL41 strain challenged with Campylobacter jejuni. Folia Microbiol. 2017, 62, 89–97. [Google Scholar] [CrossRef]
- Pinto, M.G.V.; Gomez, M.R.; Seifert, S.; Watzl, B.; Holzapfel, W.H.; Franz, C.M. Lactobacilli stimulate the innate immune response and modulate the TLR expression of HT29 intestinal epithelial cells in vitro. Int. J. Food Microbiol. 2009, 133, 86–93. [Google Scholar] [CrossRef]
- De Kivit, S.; Van Hoffen, E.; Korthagen, N.; Garssen, J.; Willemsen, L.E. Apical TLR ligation of intestinal epithelial cells drives a Th1-polarized regulatory or inflammatory type effector response in vitro. Immunobiol. 2011, 216, 518–527. [Google Scholar] [CrossRef]
- Lee, J.; Mo, J.-H.; Katakura, K.; Alkalay, I.; Rucker, A.N.; Liu, Y.-T.; Lee, H.-K.; Shen, C.; Cojocaru, G.; Shenouda, S.; et al. Maintenance of colonic homeostasis by distinctive apical TLR9 signalling in intestinal epithelial cells. Nat. Cell. Biol. 2006, 8, 1327–1336. [Google Scholar] [CrossRef]
- De Kivit, S.; Kraneveld, A.D.; Knippels, L.M.; Van Kooyk, Y.; Garssen, J.; Willemsen, L.E. Intestinal Epithelium-Derived Galectin-9 Is Involved in the Immunomodulating Effects of Nondigestible Oligosaccharides. J. Innate Immun. 2013, 5, 625–638. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Alkie, T.N.; Hodgins, D.C.; Shojadoost, B.; Sharif, S. Characterization of host responses induced by Toll-like receptor ligands in chicken cecal tonsil cells. Vet. Immunol. Immunopathol. 2016, 174, 19–25. [Google Scholar] [CrossRef]
- Bouladoux, N.; Hall, J.; Grainger, J.R.; Dos Santos, L.M.; Kann, M.; Nagarajan, V.; Verthelyi, D.; Belkaid, Y. Regulatory role of suppressive motifs from commensal DNA. Mucosal Immunol. 2012, 5, 623–634. [Google Scholar] [CrossRef]
- He, H.; Genovese, K.J.; Swaggerty, C.L.; MacKinnon, K.M.; Kogut, M.H. Co-stimulation with TLR3 and TLR21 ligands synergistically up-regulates Th1-cytokine IFN-γ and regulatory cytokine IL-10 expression in chicken monocytes. Dev. Comp. Immunol. 2012, 36, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, K.T.; Reddy, M.R.; Raveendranathan, D.N.; Murugesan, S.; Chatterjee, R.N.; Ullengala, R.; Haunshi, S. Differential expression of Toll-like receptor mRNA in White Leghorn and indigenous chicken of India. Vet. Res. Commun. 2010, 34, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Abasht, B.; Kaiser, M.G.; Van Der Poel, J.; Lamont, S.J. Genetic lines differ in Toll-like receptor gene expression in spleens of chicks inoculated with Salmonella enterica serovar Enteritidis. Poult. Sci. 2009, 88, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Sławińska, A.; D’Andrea, M.; Pilla, F.; Bednarczyk, M.; Siwek, M. Expression profiles of Toll-like receptors 1, 2 and 5 in selected organs of commercial and indigenous chickens. J. Appl. Genet. 2013, 54, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broom, L.J. Host⁻Microbe Interactions and Gut Health in Poultry-Focus on Innate Responses. Microorganisms 2019, 7, 139. [Google Scholar] [CrossRef] [Green Version]
- Neulen, M.L.; Viertlboeck, B.C.; Straub, C.; Gobel, T.W. Identification of novel chicken CD4(+) CD3(-) blood population with NK cell like features. Dev. Comp. Immunol. 2015, 49, 72–78. [Google Scholar] [CrossRef]
- Zitti, B.; Bryceson, Y.T. Natural killer cells in inflammation and autoimmunity. Cytokine Growth Factor Rev. 2018, 42, 37–46. [Google Scholar] [CrossRef]
- Qiu, Y.; Jiang, Z.; Hu, S.; Wang, L.; Ma, X.; Yang, X. Lactobacillus plantarum Enhanced IL-22 Production in Natural Killer (NK) Cells That Protect the Integrity of Intestinal Epithelial Cell Barrier Damaged by Enterotoxigenic Escherichia coli. Int. J. Mol. Sci. 2017, 18, 2409. [Google Scholar] [CrossRef] [Green Version]
- Fenzl, L.; Göbel, T.W.; Neulen, M.-L. γδ T cells represent a major spontaneously cytotoxic cell population in the chicken. Dev. Comp. Immunol. 2017, 73, 175–183. [Google Scholar] [CrossRef]
- Walliser, I.; Gobel, T.W. Chicken IL-17A is expressed in alphabeta and gammadelta T cell subsets and binds to a receptor present on macrophages, and T cells. Dev. Comp. Immunol. 2018, 81, 44–53. [Google Scholar] [CrossRef]
- Veldhoen, M. Interleukin 17 is a chief orchestrator of immunity. Nat. Immunol. 2017, 18, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-M.; Jang, H.; Jang, S.-E.; Han, M.; Kim, D.-H. Lactobacillus fermentum IM12 attenuates inflammation in mice by inhibiting NF-κB-STAT3 signalling pathway. Benef. Microbes 2017, 8, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Cash, H.A.; Farmer, S.; Keller, D. Inactivated probiotic Bacillus coagulans GBI-30 induces complex immune activating, anti-inflammatory, and regenerative markers in vitro. J. Inflamm. Res. 2017, 10, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Iliev, I.D.; Spadoni, I.; Mileti, E.; Matteoli, G.; Sonzogni, A.M.; Sampietro, G.M.; Foschi, D.; Caprioli, F.; Viale, G.; Rescigno, M. Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells. Gut 2009, 58, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Spadoni, I.; Iliev, I.D.; Rossi, G.; Rescigno, M. Dendritic cells produce TSLP that limits the differentiation of Th17 cells, fosters Treg development, and protects against colitis. Mucosal Immunol. 2012, 5, 184–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson-Lindbom, B.; Svensson, M.; Pabst, O.; Palmqvist, C.; Marquez, G.; Förster, R.; Agace, W.W. Functional specialization of gut CD103+ dendritic cells in the regulation of tissue-selective T cell homing. J. Exp. Med. 2005, 202, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, N.E.; Eberl, M. Human γδ T-Cell Control of Mucosal Immunity and Inflammation. Front. Immunol. 2018, 9, 985. [Google Scholar] [CrossRef]
- Adel-Patient, K.; Wavrin, S.; Bernard, H.; Meziti, N.; Ah-Leung, S.; Wal, J.M. Oral tolerance and Treg cells are induced in BALB/c mice after gavage with bovine β-lactoglobulin. Allergy 2011, 66, 1312–1321. [Google Scholar] [CrossRef]
- Tsuji, M.; Komatsu, N.; Kawamoto, S.; Suzuki, K.; Kanagawa, O.; Honjo, T.; Hori, S.; Fagarasan, S. Preferential generation of follicular B helper T cells from Foxp3+ T cells in gut Peyer’s patches. Science 2009, 323, 1488–1492. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, X.; He, B.; Hoang, T.K.; Taylor, C.M.; Blanchard, E.; Freeborn, J.; Park, S.; Luo, M.; Couturier, J.; et al. Lactobacillus reuteri DSM 17938 feeding of healthy newborn mice regulates immune responses while modulating gut microbiota and boosting beneficial metabolites. Am. J. Physiol. Liver Physiol. 2019, 317, G824–G838. [Google Scholar] [CrossRef]
- Sang, Y.; Blecha, F. Antimicrobial peptides and bacteriocins: Alternatives to traditional antibiotics. Anim. Heal. Res. Rev. 2008, 9, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Andreu, D.; Rivas, L. Animal antimicrobial peptides: An overview. Biopolymers 1998, 47, 415–433. [Google Scholar] [CrossRef]
- Sumi, C.D.; Yang, B.W.; Yeo, I.-C.; Hahm, Y.T. Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Can. J. Microbiol. 2015, 61, 93–103. [Google Scholar] [CrossRef]
- Grant, A.; Gay, C.G.; Lillehoj, H.S. Bacillusspp.as direct-fed microbial antibiotic alternatives to enhance growth, immunity, and gut health in poultry. Avian Pathol. 2018, 47, 339–351. [Google Scholar] [CrossRef]
- Ben Lagha, A.; Haas, B.; Gottschalk, M.; Grenier, D. Antimicrobial potential of bacteriocins in poultry and swine production. Vet. Res. 2017, 48, 22. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Genovese, K.J.; He, H.; Swaggerty, C.L.; Jiang, Y. Modulation of Chicken Intestinal Immune Gene Expression by Small Cationic Peptides as Feed Additives during the First Week Posthatch. Clin. Vaccine Immunol. 2013, 20, 1440–1448. [Google Scholar] [CrossRef] [Green Version]
- Cuperus, T.; Coorens, M.; Van Dijk, A.; Haagsman, H.P. Avian host defense peptides. Dev. Comp. Immunol. 2013, 41, 352–369. [Google Scholar] [CrossRef]
- Lee, M.O.; Jang, H.-J.; Rengaraj, D.; Yang, S.-Y.; Han, J.Y.; Lamont, S.J.; Womack, J.E. Tissue expression and antibacterial activity of host defense peptides in chicken. BMC Vet. Res. 2016, 12, 231. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Sunkara, L.T. Avian Antimicrobial Host Defense Peptides: From Biology to Therapeutic Applications. Pharmaceuticals 2014, 7, 220–246. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, K.T.; Verma, P.; Reddy, M.R. Differential gene expression of antimicrobial peptides beta defensins in the gastrointestinal tract of Salmonella serovar Pullorum infected broiler chickens. Vet. Res. Commun. 2012, 36, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Rengaraj, D.; Truong, A.D.; Lillehoj, H.S.; Han, J.Y.; Hong, Y.H. Expression and regulation of avian beta-defensin 8 protein in immune tissues and cell lines of chickens. Asian-Australas. J. Anim. Sci. 2018, 31, 1516–1524. [Google Scholar] [CrossRef] [PubMed]
- Coorens, M.; Van Dijk, A.; Bikker, F.; Veldhuizen, E.J.A.; Haagsman, H.P. Importance of Endosomal Cathelicidin Degradation To Enhance DNA-Induced Chicken Macrophage Activation. J. Immunol. 2015, 195, 3970–3977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Hou, C.; Wang, G.; Jia, H.; Yu, H.; Zeng, X.; Thacker, P.A.; Zhang, G.; Qiao, S. Lactobacillus reuteri I5007 Modulates Intestinal Host Defense Peptide Expression in the Model of IPEC-J2 Cells and Neonatal Piglets. Nutrients 2017, 9, 559. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Metabolic Pathway | Known Functions | Putative Role on Inflammation |
|---|---|---|
| Acute Phase Proteins | ||
| Haptoglobin | ↓Free hemoglobin, ↓oxidative damage | Anti-inflammatory |
| Ceruloplasmin | ↓Neutrophils, ↑peroxide scavenging | Anti-inflammatory |
| C-Reactive protein | Activation of complement | Pro-inflammatory |
| Mannose-binding lectin | Activation of complement, reduction of CpG-ODN motifs, increase of phagocytosis against pathogens | Pro-inflammatory |
| Complement | ||
| Complement proteins in the epithelium | Induction of phagocytosis, cytolysis, and inflammatory response | Pro-inflammatory |
| TLR | ||
| TLR2 and 4 | Detection of LPS, inflammatory response through the NF-κB pathway | Pro-inflammatory |
| TLR21 basolateral | Detection of CpG-ODN motifs, activation of NF-κB pathway | Pro-inflammatory |
| TLR21 apical | Detection of CpG-ODN motifs, inactivation of NF-κB pathway | Anti-inflammatory |
| Immune cells | ||
| NK cells | Kill infected cells, cytotoxicity mediation, maintenance of homeostasis, cell barrier integrity | Pro- and anti-inflammatory |
| γδ T cells | Cytotoxicity, ↑IL17a | Pro-inflammatory |
| Treg cells | ↑Tolerance, ↑IL10, ↑TGF-β, ↑sIgA, ↓hypersensitivity to food | Anti-inflammatory |
| Antimicrobial peptides | ||
| Expression of HDP | ↑Antimicrobial activity | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarradas, J.; Tous, N.; Esteve-Garcia, E.; Brufau, J. The Control of Intestinal Inflammation: A Major Objective in the Research of Probiotic Strains as Alternatives to Antibiotic Growth Promoters in Poultry. Microorganisms 2020, 8, 148. https://doi.org/10.3390/microorganisms8020148
Tarradas J, Tous N, Esteve-Garcia E, Brufau J. The Control of Intestinal Inflammation: A Major Objective in the Research of Probiotic Strains as Alternatives to Antibiotic Growth Promoters in Poultry. Microorganisms. 2020; 8(2):148. https://doi.org/10.3390/microorganisms8020148
Chicago/Turabian StyleTarradas, Joan, Núria Tous, Enric Esteve-Garcia, and Joaquim Brufau. 2020. "The Control of Intestinal Inflammation: A Major Objective in the Research of Probiotic Strains as Alternatives to Antibiotic Growth Promoters in Poultry" Microorganisms 8, no. 2: 148. https://doi.org/10.3390/microorganisms8020148