Validating Thermal Lethality to Salmonella enterica in Chicken Blood by Simulated Commercial Rendering


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Inoculum Preparation
2.1.1. Preliminary Enumeration of Salmonellae from Non-Inoculated Chicken Blood
2.1.2. Preliminary Experiment to Verify Salmonella Growth for Subsequent Inoculum Preparation
2.1.3. Preliminary Experiment to Identify Optimal Medium for Selective Recovery of Non-Injured and Injured Salmonella Cells from Heat-Treated Chicken Blood
2.2. Sample Inoculation with Salmonella Cocktail
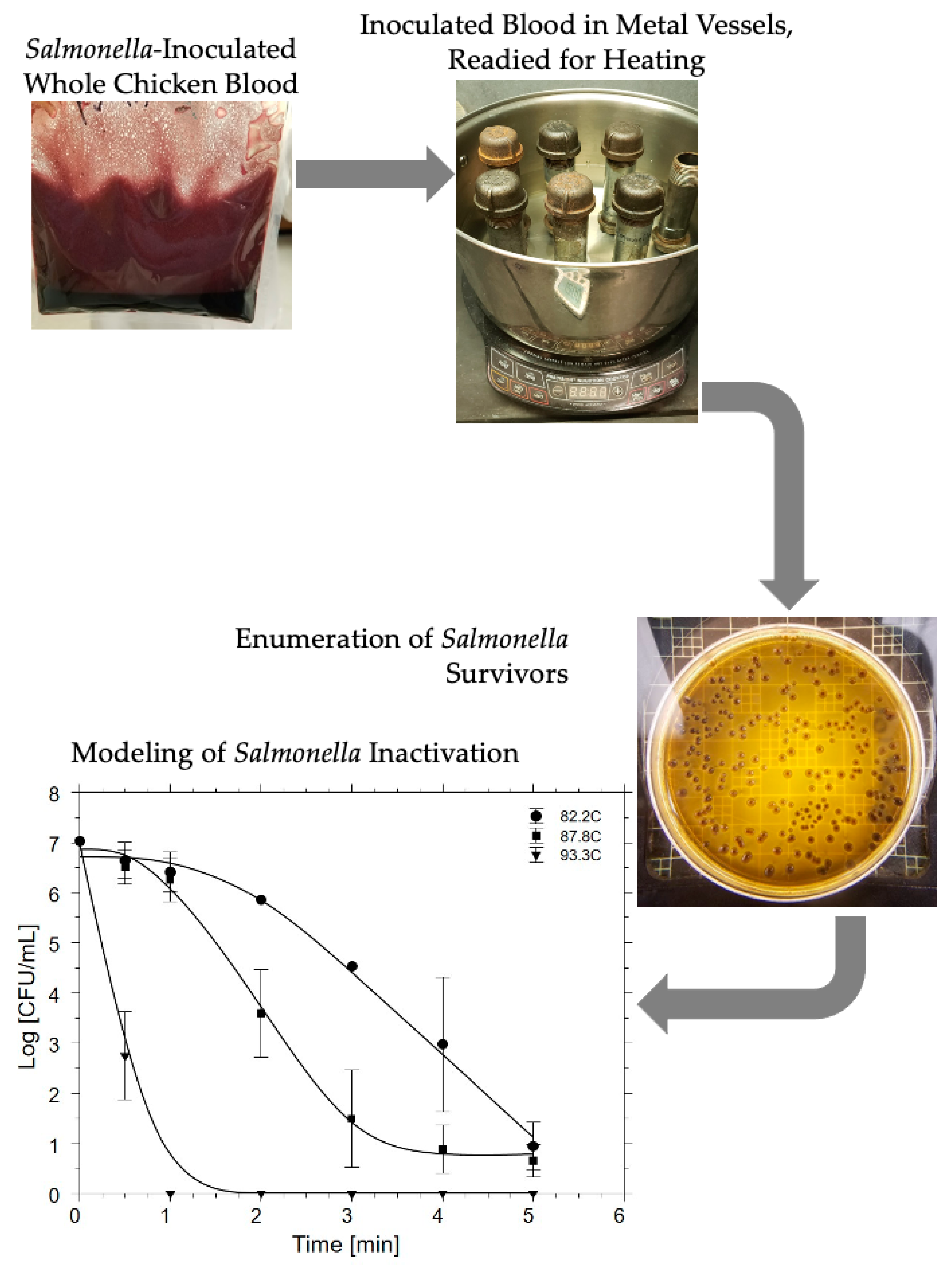
2.3. Lethality to Salmonella Inoculated into Chicken Blood
2.4. Temperature-Dependent Thermal Lethality to Salmonella in Chicken Blood
2.5. Data Analysis
3. Results and Discussion
3.1. Results of Preliminary Experiments Enumerating Salmonellae from Non-Inoculated Chicken Blood and Overnight Growth of Salmonella Isolates for Inoculum Preparation
3.2. Identification of Suitable Medium for Use in Selective Plating and Recovery of Non-Injured and Injured Salmonella Cells from Heated Chicken Blood
3.3. Total Lethality Achieved in Chicken Blood against Salmonella
3.4. Inactivation of S. enterica in Chicken Blood as a Function of Thermal Processing
3.5. Thermal Death Kinetics for Microorganisms in Chicken Blood as a Function of Heating Temperature
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Meeker, D.L.; Hamilton, C.R. An overview of the rendering industry In Essential Rendering: All about the Animal by-Products Industry; Meeker, D.L., Ed.; National Renderers Association: Alexandria, VA, USA, 2006; pp. 1–16. [Google Scholar]
- Pepe, T.; De Dominicis, R.; Esposito, G.; Ventrone, I.; Fratamico, P.M.; Cortesi, M.L. Detection of Campylobacter from poultry carcass skin samples at slaughter in southern Italy. J. Food Prot. 2009, 72, 1718–1721. [Google Scholar] [CrossRef]
- Narvaez-Bravo, C.; Miller, M.F.; Jackson, T.; Jackson, S.; Rodas-Gonzalez, A.; Pond, K.; Echeverry, A.; Brashears, M.M. Salmonella and Escherichia coli O157:H7 prevalence in cattle and on carcasses in a vertically integrated feedlot and harvest plant in Mexico. J. Food Prot. 2013, 76, 786–795. [Google Scholar] [CrossRef]
- Bacon, R.T.; Sofos, J.N.; Belk, K.E.; Hyatt, D.R.; Smith, G.C. Prevalence and antibiotic susceptibility of Salmonella isolated from beef animal hides and carcasses. J. Food Prot. 2002, 65, 284–290. [Google Scholar] [CrossRef]
- Jiang, X. Prevalence and characterization of Salmonella in animal meals collected from rendering operations. J. Food Prot. 2016, 79, 1026–1031. [Google Scholar] [CrossRef]
- Kinley, B.; Rieck, J.; Dawson, P.; Jiang, X. Analysis of Salmonella and enterococci isolated from rendered animal products. Can. J. Microbiol. 2010, 56, 65–73. [Google Scholar] [CrossRef]
- Ge, B.; Lafon, P.C.; Carter, P.J.; McDermott, S.D.; Abbott, J.; Glenn, A.; Ayers, S.L.; Friedman, S.L.; Paige, J.C.; Wagner, D.D.; et al. Retrospective analysis of Salmonella, Campylobacter, Escherichia coli, and Enterococcus in animal feed ingredients. FoodBorne Pathog. Dis. 2013, 10, 684–691. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Title 21: Food and Drugs, Part 507: Current Good Manufacturing Practice, Hazard Analysis, and Risk-Based Preventive Controls for Food for Animals. Available online: https://www.ecfr.gov/cgi-bin/text-idx?SID=b4e880ff425c66edad94b50e64f286a3&mc=true&node=pt21.6.507&rgn=div5 (accessed on 23 September 2020).
- Consortium of Food Process Validation Experts. Validation of antimicrobial interventions for small and very small processors: A how-to guide to develop and conduct validations. Food Prot. Trends 2013, 33, 95–104. [Google Scholar]
- Ceylan, E.; Bautista, D.A. Evaluating Pediococcus acidilactici and Enterococcus faecium NRRL B-2354 as a thermal surrogate microorganisms for Salmonella for in-plant validation studies of low-moisture pet food products. J. Food Prot. 2015, 78, 934–939. [Google Scholar] [CrossRef]
- Bianchini, A.; Stratton, J.; Weier, S.; Hartter, T.; Plattner, B.; Rokey, G.; Hertzel, G.; Gompa, L.; Martinez, B.; Eskridge, K.M. Use of Enterococcus faecium as a surrogate for Salmonella enterica during extrusion of a balanced carbohydrate-protein meal. J. Food Prot. 2014, 77, 75–82. [Google Scholar] [CrossRef]
- Jones-Ibarra, A.M.; Acuff, G.R.; Alvarado, C.Z.; Taylor, T.M. Validation of thermal lethality against Salmonella enterica in poultry offal during rendering. J. Food Prot. 2017, 80, 1422–1428. [Google Scholar] [CrossRef]
- Gurtler, J.B.; Kornacki, J.R. Comparison of supplements to enhance recovery of heat-injured Salmonella from egg albumen. Lett. Appl. Microbiol. 2009, 49, 503–509. [Google Scholar] [CrossRef]
- Geeraerd, A.H.; Valdramidis, V.P.; van Impe, J.F. GInaFiT, a freeware tool to assess non-log-linear microbial survivor curves. Int. J. Food Microbiol. 2005, 102, 95–105. [Google Scholar] [CrossRef]
- Russell, S.M.; Fletcher, D.L.; Pancorbo, O.C.; Merka, W.C. Effect of lactic acid fermentation on bacterial pathogens and indicator organisms in broiler processing waste. Poult. Sci. 1993, 72, 1573–1576. [Google Scholar] [CrossRef]
- USDA-FSIS. Performance standards for the production of certain meat and poultry products. Fed. Regist. 1999, 64, 732–749. [Google Scholar]
- FDA. Compliance Policy Guide, Section 690.800: Salmonella in food for animals. Fed. Regist. 2013, 78, 42526–42527. [Google Scholar]
- Valdramidis, V.P.; Bernaerts, K.; Van Impe, J.F.; Geeraerd, A.H. An alternative approach to non-log-linear thermal microbial inactivation: Modelling the number of log cycles reduction with respect to temperature. Food Technol. Biotechnol. 2005, 43, 321–327. [Google Scholar]
- Vaddella, V.; Pitesky, M.; Cao, W.; Govinthasamy, V.; Shi, J.; Pandey, P. Assessing Salmonella typhimurium persistence in poultry carcasses under multiple thermal conditions consistent with composting and wet rendering. Poult. Sci. 2016, 95, 705–714. [Google Scholar] [CrossRef]
- Peleg, M. Microbial survivor curves—The reality of flat “shoulders” and absolute thermal death times. Food Res. Int. 2000, 33, 531–538. [Google Scholar] [CrossRef]
- Murphy, R.L.; Woerner, D.R.; Geornaras, I.; Martin, J.N.; Yang, H.; Sofos, J.N.; Delmore, R.J.; Belk, K.E. Determining process lethality of Escherichia coli O157:H7 and Salmonella in heat treated, rendered oil. Meat Muscle Biol. 2016, 1, 124. [Google Scholar] [CrossRef]
- Ramirez-Hernandez, A.; Inestroza, B.; Parks, A.; Brashears, M.M.; Sanchez-Plata, M.X.; Echeverry, A. Thermal inactivation of Salmonella in high-fat rendering meat products. J. Food Prot. 2018, 81, 54–58. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Golden, M.H.; Whiting, R.C. Differentiation of the effects of pH and lactic or acetic acid concentration on the kinetics of Listeria monocytogenes inactivation. J. Food Prot. 1993, 56, 474–478. [Google Scholar] [CrossRef]
- Jackson, T.C.; Hardin, M.D.; Acuff, G.R. Heat resistance of Escherichia coli O157:H7 in a nutrient medium and in ground beef patties as influenced by storage and holding temperatures. J. Food Prot. 1996, 59, 230–237. [Google Scholar] [CrossRef]
- Juneja, V.K.; Eblen, B.S.; Marks, H.M. Modeling non-linear survival curves to calculate thermal inactivation of Salmonella in poultry of different fat levels. Int. J. Food Microbiol. 2001, 70, 37–51. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heating Temperature (°C) | ||||
|---|---|---|---|---|
| 82.2 | 87.8 | 93.3 | p > F; Pooled SE 2 | |
| D-value 1 | 0.61 ± 0.18 A (0.22, 1.06) | 0.41 ± 0.19 A,B (0.00, 0.88) | 0.12 ± 0.02 B,C (0.04, 0.21) | 0.023; 0.13 |
| t4D | 4.12 ± 0.19 A (3.32, 4.92) | 2.42 ± 0.38 B (0.80, 4.04) | 0.50 ± 0.08 C (0.17, 0.83) | 0.0001; 0.35 |
| Model Parameter 1 | 82.2 °C | 87.8 °C | 93.3 °C | p > F; Pooled SE 2 |
|---|---|---|---|---|
| SL (min) | 1.62 ± 0.12 A | 0.71 ± 0.09 B | 0.0 ± 0.0 C | <0.0001; 0.125 |
| kmax (1/min) | 3.80 ± 0.29 | 5.59 ± 0.47 | 19.71 ± 0.04 | |
| Nres (log CFU/mL) | -- 3 | 0.77 ± 0.17 | -- | |
| N0 (log CFU/mL) | 6.71 ± 0.16 | 6.87 ± 0.19 | 7.03 ± 0.01 | 0.089; 0.117 |
| RMSE | 0.26 | 0.23 | 0.01 | |
| Adjusted R2 | 0.99 | 0.99 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong de la Rosa, C.; Daniels, K.A.; Moreira, R.G.; Kerth, C.R.; Taylor, T.M. Validating Thermal Lethality to Salmonella enterica in Chicken Blood by Simulated Commercial Rendering. Microorganisms 2020, 8, 2009. https://doi.org/10.3390/microorganisms8122009
Wong de la Rosa C, Daniels KA, Moreira RG, Kerth CR, Taylor TM. Validating Thermal Lethality to Salmonella enterica in Chicken Blood by Simulated Commercial Rendering. Microorganisms. 2020; 8(12):2009. https://doi.org/10.3390/microorganisms8122009
Chicago/Turabian StyleWong de la Rosa, Caleb, Kourtney A. Daniels, Rosana G. Moreira, Chris R. Kerth, and Thomas M. Taylor. 2020. "Validating Thermal Lethality to Salmonella enterica in Chicken Blood by Simulated Commercial Rendering" Microorganisms 8, no. 12: 2009. https://doi.org/10.3390/microorganisms8122009