Role of SrtA in Pathogenicity of Staphylococcus lugdunensis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Media
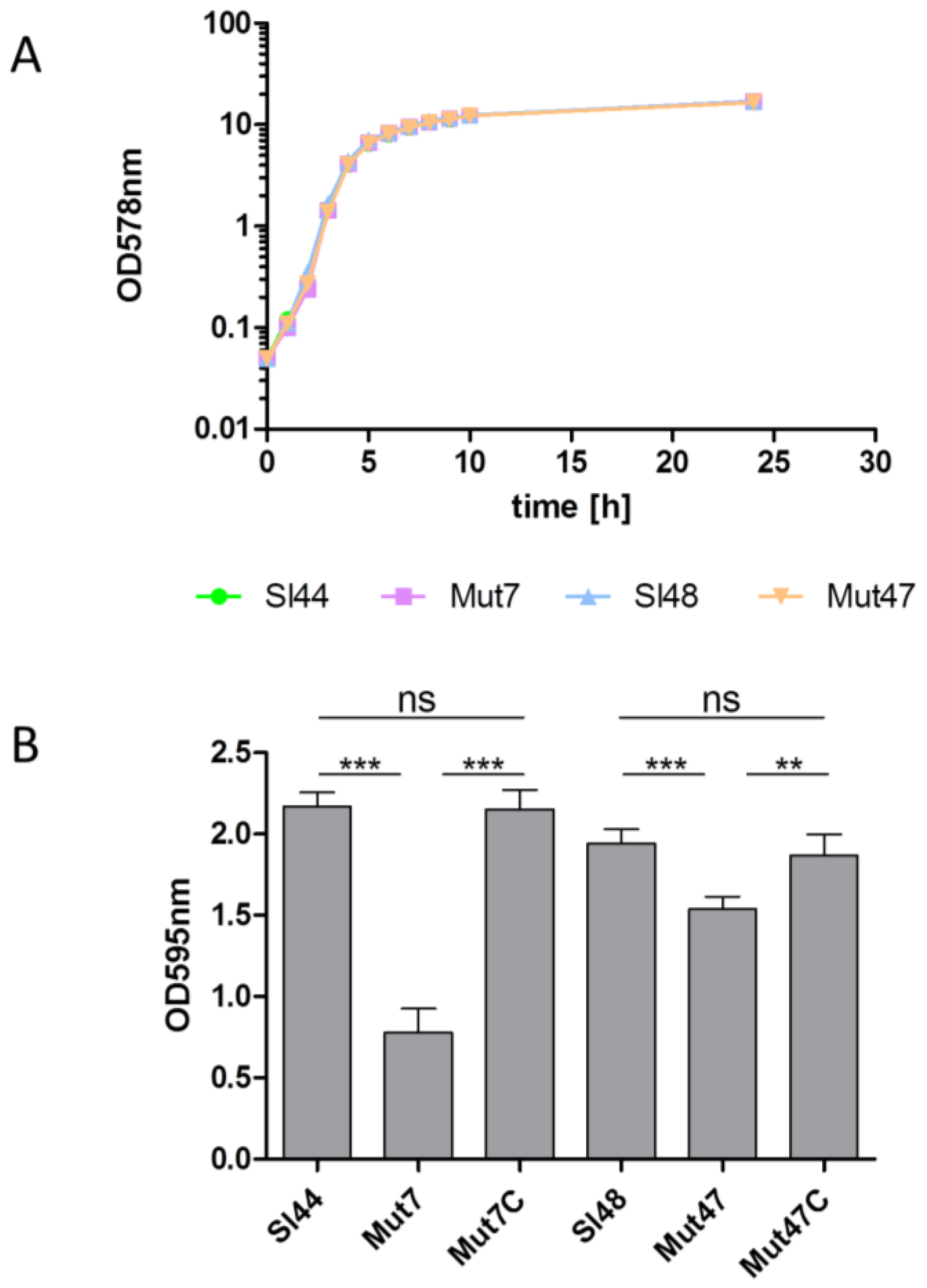
2.2. Growth Characteristics
2.3. Characterization of Agglutination
2.4. Biofilm Formation
2.5. DNA Manipulations and Transformations
2.6. Construction of a srtA-Deficient Mutant
2.7. The Complementation of a srtA-Deficient Mutant
2.8. Cell Protein Preparations
2.9. SDS-PAGE and Ligand Overlay Analysis
2.10. Expression of Recombinant Sortase-A
2.11. Polyclonal Antibodies
2.12. ELISA Adherence assays
2.13. Cell Culture and Flow Cytometric Invasion Assay
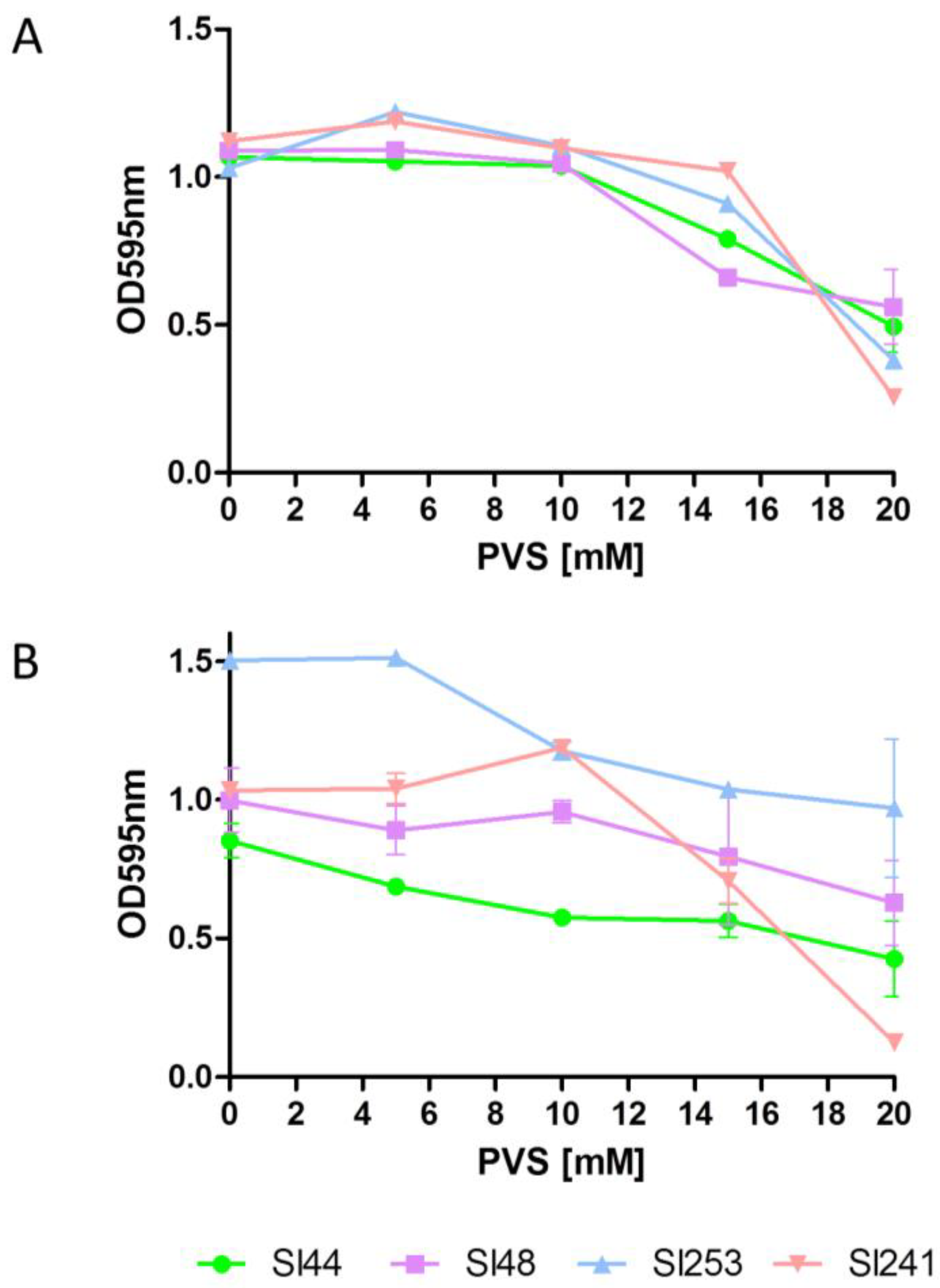
2.14. Sortase Inhibitor PVS
2.15. Hydroxylaminolysis of LPXTG Peptide
2.16. Real-Time Reverse-Transcription PCR (qtRT-PCR)
2.17. Statistical Analysis
3. Results
3.1. Sortase A-Dependent Proteins
3.2. Alignment of Sortase A Sequences
3.3. Generation of Sortase-A Mutants
3.4. Characterisation of the Surface Proteins by SDS Page and Western Blot Experiments
3.5. The Surface Proteomes of the Wild-Type and Mutant Strains Differ in the Absence of the SrtA
3.6. Determination of Growth and Biofilm Formation of the ΔsrtA Mutants
3.7. Recombinant SrtA As Well As Cell Extracts Catalyzes Hydroxylaminolysis
3.8. Agglutination Test and Adherence of ΔsrtA Mutants to Immobilized Fibronectin (Fn), Fibrinogen (Fg), and Vitronectin (Vn)
3.9. Sortase A Inhibition Resulted in Decreased Biofilm Formation and Binding to Fg and Fn
3.10. Hydroxylamine HCl Treatment Decrease Binding to Immobilized Fg and Fn
3.11. Adherence and Invasion to Eucaryotic Cell Lines
3.12. SrtA Influence the Gene Expression of Further Adhesins
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, M.; Bieber, L.; Smyth, R.; Kahlmeter, G. Detection and identification of Staphylococcus lugdunensis are not hampered by use of defibrinated horse blood in blood agar plates. J. Clin. Microbiol 2010, 48, 1987–1988. [Google Scholar] [CrossRef] [Green Version]
- van der Mee-Marquet, N.; Achard, A.; Mereghetti, L.; Danton, A.; Minier, M.; Quentin, R. Staphylococcus lugdunensis infections: High frequency of inguinal area carriage. J. Clin. Microbiol. 2003, 41, 1404–1409. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, U.; Kriegeskorte, A.; Schubert, T.; Peters, G.; Rudack, C.; Pieper, D.H.; Wos-Oxley, M.; Becker, K. The culturome of the human nose habitats reveals individual bacterial fingerprint patterns. Environ. Microbiol. 2016, 18, 2130–2142. [Google Scholar] [CrossRef]
- Frank, K.L.; Del Pozo, J.L.; Patel, R. From clinical microbiology to infection pathogenesis: How daring to be different works for Staphylococcus lugdunensis. Clin. Microbiol. Rev. 2008, 21, 111–133. [Google Scholar] [CrossRef] [Green Version]
- Anguera, I.; Del, R.A.; Miro, J.M.; Matinez-Lacasa, X.; Marco, F.; Guma, J.R.; Quaglio, G.; Claramonte, X.; Moreno, A.; Mestres, C.A.; et al. Staphylococcus lugdunensis infective endocarditis: Description of 10 cases and analysis of native valve, prosthetic valve, and pacemaker lead endocarditis clinical profiles. Heart 2005, 91, e10. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Steinbacher, T.; Peters, G.; Heilmann, C.; Becker, K. The adhesive properties of the Staphylococcus lugdunensis multifunctional autolysin AtlL and its role in biofilm formation and internalization. Int. J. Med. Microbiol. 2015, 305, 129–139. [Google Scholar] [CrossRef]
- Marlinghaus, L.; Becker, K.; Korte, M.; Neumann, S.; Gatermann, S.G.; Szabados, F. Construction and characterization of three knockout mutants of the fbl gene in Staphylococcus lugdunensis. APMIS 2012, 120, 108–116. [Google Scholar] [CrossRef]
- Nilsson, M.; Bjerketorp, J.; Wiebensjo, A.; Ljungh, A.; Frykberg, L.; Guss, B. A von Willebrand factor-binding protein from Staphylococcus lugdunensis. FEMS Microbiol. Lett. 2004, 234, 155–161. [Google Scholar] [CrossRef]
- Nilsson, M.; Bjerketorp, J.; Guss, B.; Frykberg, L. A fibrinogen-binding protein of Staphylococcus lugdunensis. FEMS Microbiol. Lett. 2004, 241, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Donvito, B.; Etienne, J.; Denoroy, L.; Greenland, T.; Benito, Y.; Vandenesch, F. Synergistic hemolytic activity of Staphylococcus lugdunensis is mediated by three peptides encoded by a non-agr genetic locus. Infect. Immun. 1997, 65, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, C. Adhesion mechanisms of staphylococci. Adv. Exp. Med. Biol. 2011, 715, 105–123. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Patti, J.M.; Höök, M. Microbial adhesins recognizing extracellular matrix macromolecules. Curr. Opin. Cell. Biol. 1994, 6, 752–758. [Google Scholar] [CrossRef]
- Navarre, W.W.; Schneewind, O. Proteolytic cleavage and cell wall anchoring at the LPXTG motif of surface proteins in gram-positive bacteria. Mol. Microbiol. 1994, 14, 115–121. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, G.; Ton-That, H.; Schneewind, O. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 1999, 285, 760–763. [Google Scholar] [CrossRef]
- Ton-That, H.; Liu, G.; Mazmanian, S.K.; Faull, K.F.; Schneewind, O. Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the LPXTG motif. Proc. Natl. Acad. Sci. USA 1999, 96, 12424–12429. [Google Scholar] [CrossRef] [Green Version]
- Perry, A.M.; Ton-That, H.; Mazmanian, S.K.; Schneewind, O. Anchoring of surface proteins to the cell wall of Staphylococcus aureus. III. Lipid II is an in vivo peptidoglycan substrate for sortase-catalyzed surface protein anchoring. J. Biol. Chem. 2002, 277, 16241–16248. [Google Scholar] [CrossRef] [Green Version]
- Ruzin, A.; Severin, A.; Ritacco, F.; Tabei, K.; Singh, G.; Bradford, P.A.; Siegel, M.M.; Projan, S.J.; Shlaes, D.M. Further evidence that a cell wall precursor [C(55)-MurNAc-(peptide)-GlcNAc] serves as an acceptor in a sorting reaction. J. Bacteriol. 2002, 184, 2141–2147. [Google Scholar] [CrossRef] [Green Version]
- Spirig, T.; Weiner, E.M.; Clubb, R.T. Sortase enzymes in Gram-positive bacteria. Mol. Microbiol. 2011, 82, 1044–1059. [Google Scholar] [CrossRef]
- Oshida, T.; Sugai, M.; Komatsuzawa, H.; Hong, Y.M.; Suginaka, H.; Tomasz, A. A Staphylococcus aureus autolysin that has an N-acetylmuramoyl-L-alanine amidase domain and an endo-beta-N-acetylglucosaminidase domain: Cloning, sequence analysis, and characterization. Proc. Natl. Acad. Sci. USA 1995, 92, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Liu, G.; Jensen, E.R.; Lenoy, E.; Schneewind, O. Staphylococcus aureus sortase mutants defective in the display of surface proteins and in the pathogenesis of animal infections. Proc. Natl. Acad. Sci. USA 2000, 97, 5510–5515. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Ton-That, H.; Su, K.; Schneewind, O. An iron-regulated sortase anchors a class of surface protein during Staphylococcus aureus pathogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 2293–2298. [Google Scholar] [CrossRef] [Green Version]
- Heilbronner, S.; Hanses, F.; Monk, I.R.; Speziale, P.; Foster, T.J. Sortase A promotes virulence in experimental Staphylococcus lugdunensis endocarditis. Microbiology 2013, 159, 2141–2152. [Google Scholar] [CrossRef] [Green Version]
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737. [Google Scholar] [CrossRef] [Green Version]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In vitro cultivation of human tumors: Establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef]
- Fogh, J.; Wright, W.C.; Loveless, J.D. Absence of HeLa cell contamination in 169 cell lines derived from human tumors. J. Natl. Cancer Inst. 1977, 58, 209–214. [Google Scholar] [CrossRef]
- Khan, S.A.; Novick, R.P. Terminal nucleotide sequences of Tn551, a transposon specifying erythromycin resistance in Staphylococcus aureus: Homology with Tn3. Plasmid 1980, 4, 148–154. [Google Scholar] [CrossRef]
- Palma, M.; Nozohoor, S.; Schennings, T.; Heimdahl, A.; Flock, J.I. Lack of the extracellular 19-kilodalton fibrinogen-binding protein from Staphylococcus aureus decreases virulence in experimental wound infection. Infect. Immun. 1996, 64, 5284–5289. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Becker, K.; von Eiff, C.; Schrenzel, J.; Peters, G.; Herrmann, M. Identification and characterization of a novel 38.5-kilodalton cell surface protein of Staphylococcus aureus with extended-spectrum binding activity for extracellular matrix and plasma proteins. J. Bacteriol. 2001, 183, 6778–6786. [Google Scholar] [CrossRef] [Green Version]
- Tuchscherr, L.; Löffler, B.; Buzzola, F.R.; Sordelli, D.O. Staphylococcus aureus adaptation to the host and persistence: Role of loss of capsular polysaccharide expression. Future Microbiol. 2010, 5, 1823–1832. [Google Scholar] [CrossRef]
- Hussain, M.; Haggar, A.; Heilmann, C.; Peters, G.; Flock, J.I.; Herrmann, M. Insertional inactivation of Eap in Staphylococcus aureus strain Newman confers reduced staphylococcal binding to fibroblasts. Infect. Immun. 2002, 70, 2933–2940. [Google Scholar] [CrossRef] [Green Version]
- Quentmeier, H.; Zaborski, M.; Drexler, H.G. The human bladder carcinoma cell line 5637 constitutively secretes functional cytokines. Leuk. Res. 1997, 21, 343–350. [Google Scholar] [CrossRef]
- Sinha, B.; Francois, P.; Que, Y.A.; Hussain, M.; Heilmann, C.; Moreillon, P.; Lew, D.; Krause, K.H.; Peters, G.; Herrmann, M. Heterologously expressed Staphylococcus aureus fibronectin-binding proteins are sufficient for invasion of host cells. Infect. Immun. 2000, 68, 6871–6878. [Google Scholar] [CrossRef] [Green Version]
- Juuti, K.M.; Sinha, B.; Werbick, C.; Peters, G.; Kuusela, P.I. Reduced adherence and host cell invasion by methicillin-resistant Staphylococcus aureus expressing the surface protein Pls. J. Infect. Dis. 2004, 189, 1574–1584. [Google Scholar] [CrossRef] [Green Version]
- Sinha, B.; Francois, P.P.; Nüße, O.; Foti, M.; Hartford, O.M.; Vaudaux, P.; Foster, T.J.; Lew, D.P.; Herrmann, M.; Krause, K.H. Fibronectin-binding protein acts as Staphylococcus aureus invasin via fibronectin bridging to integrin α5β1. Cell. Microbiol. 1999, 1, 101–117. [Google Scholar] [CrossRef]
- Hussain, M.; Schafer, D.; Juuti, K.M.; Peters, G.; Haslinger-Loffler, B.; Kuusela, P.I.; Sinha, B. Expression of Pls (plasmin sensitive) in Staphylococcus aureus negative for pls reduces adherence and cellular invasion and acts by steric hindrance. J. Infect. Dis. 2009, 200, 107–117. [Google Scholar] [CrossRef]
- Pinsky, B.A.; Samson, D.; Ghafghaichi, L.; Baron, E.J.; Banaei, N. Comparison of real-time PCR and conventional biochemical methods for identification of Staphylococcus lugdunensis. J. Clin. Microbiol. 2009, 47, 3472–3477. [Google Scholar] [CrossRef] [Green Version]
- Heilbronner, S.; Holden, M.T.; van Tonder, A.; Geoghegan, J.A.; Foster, T.J.; Parkhill, J.; Bentley, S.D. Genome sequence of Staphylococcus lugdunensis N920143 allows identification of putative colonization and virulence factors. FEMS Microbiol. Lett. 2011, 322, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, H.; Ishikawa, S.; Nagamune, T. Design of Ca2+-independent Staphylococcus aureus sortase A mutants. Biotechnol. Bioeng. 2012, 109, 2955–2961. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, D.S.; Oh, M.N.; Chung, S.C.; Lee, J.S.; Oh, K.B. Inhibition of the bacterial surface protein anchoring transpeptidase sortase by isoquinoline alkaloids. Biosci. Biotechnol. Biochem. 2004, 68, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Maresso, A.W.; Schneewind, O. Sortase as a target of anti-infective therapy. Pharmacol. Rev. 2008, 60, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Que, Y.A.; Francois, P.; Haefliger, J.A.; Entenza, J.M.; Vaudaux, P.; Moreillon, P. Reassessing the role of Staphylococcus aureus clumping factor and fibronectin-binding protein by expression in Lactococcus lactis. Infect. Immun. 2001, 69, 6296–6302. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, I.M.; Mazmanian, S.K.; Schneewind, O.; Verdrengh, M.; Bremell, T.; Tarkowski, A. On the role of Staphylococcus aureus sortase and sortase-catalyzed surface protein anchoring in murine septic arthritis. J. Infect. Dis. 2002, 185, 1417–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsson, M.; Petersson, A.C.; Ljungh, A. Serum and tissue protein binding and cell surface properties of Staphylococcus lugdunensis. J. Med. Microbiol. 1993, 38, 96–102. [Google Scholar] [CrossRef]
- Oh, K.B.; Oh, M.N.; Kim, J.G.; Shin, D.S.; Shin, J. Inhibition of sortase-mediated Staphylococcus aureus adhesion to fibronectin via fibronectin-binding protein by sortase inhibitors. Appl. Microbiol. Biotechnol. 2006, 70, 102–106. [Google Scholar] [CrossRef]
- Sibbald, M.J.; Yang, X.M.; Tsompanidou, E.; Qu, D.; Hecker, M.; Becher, D.; Buist, G.; van Dijl, J.M. Partially overlapping substrate specificities of staphylococcal group A sortases. Proteomics 2012, 12, 3049–3062. [Google Scholar] [CrossRef] [Green Version]
- Weidenmaier, C.; Kokai-Kun, J.F.; Kristian, S.A.; Chanturiya, T.; Kalbacher, H.; Gross, M.; Nicholson, G.; Neumeister, B.; Mond, J.J.; Peschel, A. Role of teichoic acids in Staphylococcus aureus nasal colonization, a major risk factor in nosocomial infections. Nat. Med. 2004, 10, 243–245. [Google Scholar] [CrossRef]
- van Pijkeren, J.P.; Canchaya, C.; Ryan, K.A.; Li, Y.; Claesson, M.J.; Sheil, B.; Steidler, L.; O’Mahony, L.; Fitzgerald, G.F.; van Sinderen, S.; et al. Comparative and functional analysis of sortase-dependent proteins in the predicted secretome of Lactobacillus salivarius UCC118. Appl. Environ. Microbiol. 2006, 72, 4143–4153. [Google Scholar] [CrossRef] [Green Version]
- Nobbs, A.H.; Vajna, R.M.; Johnson, J.R.; Zhang, Y.; Erlandsen, S.L.; Oli, M.W.; Kreth, J.; Brady, L.J.; Herzberg, M.C. Consequences of a sortase A mutation in Streptococcus gordonii. Microbiology 2007, 153, 4088–4097. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.W.; Lima, B.P.; Herbomel, G.G.; Gopinath, T.; McDonald, L.; Shyne, M.T.; Lee, J.K.; Kreth, J.; Ross, K.F.; Veglia, G.; et al. An intramembrane sensory circuit monitors sortase A-mediated processing of streptococcal adhesins. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Sudheesh, P.S.; Crane, S.; Cain, K.D.; Strom, M.S. Sortase inhibitor phenyl vinyl sulfone inhibits Renibacterium salmoninarum adherence and invasion of host cells. Dis. Aquat. Organ. 2007, 78, 115–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ton-That, H.; Schneewind, O. Anchor structure of staphylococcal surface proteins. IV. Inhibitors of the cell wall sorting reaction. J. Biol. Chem. 1999, 274, 24316–24320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, I.M.; Mazmanian, S.K.; Schneewind, O.; Bremell, T.; Tarkowski, A. The role of Staphylococcus aureus sortase A and sortase B in murine arthritis. Microbes Infect. 2003, 5, 775–780. [Google Scholar] [CrossRef]
- Weiss, W.J.; Lenoy, E.; Murphy, T.; Tardio, L.; Burgio, P.; Projan, S.J.; Schneewind, O.; Alksne, L. Effect of srtA and srtB gene expression on the virulence of Staphylococcus aureus in animal models of infection. J. Antimicrob. Chemother. 2004, 53, 480–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Kim, H.Y.; Schneewind, O.; Missiakas, D. Identifying protective antigens of Staphylococcus aureus, a pathogen that suppresses host immune responses. FASEB J. 2011, 25, 3605–3612. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Relevant Genotype or Plasmid | Properties | Reference or Source |
|---|---|---|---|
| S. lugdunensis strains | |||
| Sl48 | Clinical isolate | Germany b | |
| Sl44 | Clinical isolate | Germany b | |
| Mut7 | Sl48 srtA::EmR | Sl48 deficient in sortase-A | This study |
| Mut47 | Sl44 srtA::EmR | Sl44 deficient in sortase-A | This study |
| SL241 | Clinical isolate | Germany b | |
| SL253 | Clinical isolate | Germany b | |
| S. aureus strain | |||
| S. aureus Cowan 1 (ATCC 12598) | Reference isolate from septic arthritis | ATCC | |
| E. coli strains | |||
| DH5α | supE44ΔlacU169 (ϕ80 lacZΔM15) hsdR17 recA1 end A1 gyrA96 thi-1 relA1 | Cloning host | Stratagene |
| TG1 | supE hsdΔ5 thiΔ(lac-proAB) F’(traD36 proAB+ lacIq lacZΔM15) | Cloning host | Stratagene |
| DH5α (pBT37) | pBT9atlL::EmR | Shuttle vector pBT9 containing atlL::EmR | This study |
| Eukaryotic strains | |||
| EA.hy 926 cells | [25] | ||
| A549 fibroblast | [26] | ||
| Human bladder carcinoma cell line 5637 | [27] | ||
| Primer | Sequence (5′–3′) | Reference |
|---|---|---|
| srtA1FH | AAAAAGCTTTAAGAAAGCTAAAAAAATGACATAGTTG | This study |
| srtA1RE | AAAGAATTCCTCCAATAATGGTCATCAATTGGTTTGTCC | This study |
| srtA2FE | AAGAATTCTATTTATAGCAGAACAGATTAAATAATTGTAG | This study |
| srtA2RB | AAAGGATCCCATCTGAGTCAA GACTACTAGCAAGTGG | This study |
| Ery-EF, | ATATATCGATTAGGGACCTCTTTAGC | [28] |
| Ery-ER | ATATATCGATATCATGAGTATTGTCCG | [28] |
| SrtA1FH | AAAAAGCTTTAAGAAAGCTAAAAAAATGACATAGTTG | This study |
| SrtA2RB | AAAGGATCCCATCTGAGTCAAGACTACTAGCAAGTGG | This study |
| srtAF | CTCGGATCCAAACCTCATATTGATAGTTATTTACATGAC | This study |
| srtAR | CTCGGTACCTTATTTAATCTGTTCTGCTATAAATATTTTACGC | This study |
| RTFblF | GAAGCAACAACGCAGAACAA | [38] |
| RTFblR | TGCTTGTGCCTCGCTATTTA | [38] |
| RT16SF | CAGCTCGTGTCGTGAGATGT | [38] |
| RT16SR | TAGCACGTGTGTAGCCCAAA | [38] |
| RTvWbF | GGACCAGGTGAAGGTGATGT | This study |
| RTvWbR | GCCGCTGATTTTCGTGTAAT | This study |
| Genetic Identifiers (GN) | Annotation | Cleavage Motif | Size (aa) | Predicted Protein Size (kDa) | NCBI BLAST Hit (Protein 1, Strain 2, Length 3) |
|---|---|---|---|---|---|
| SLUG_00890 SLGD_00061 | IsdB | LPATG | 690 | 76.9 | Surface protein SasI, HKU09-01 |
| SLUG_00930 SLGD_00065 SasE | IsdJ | LPNTG | 646 | 71.5 | Cell surface protein IsdA, HKU09-01 LPXTG cell wall surface anchor protein, M23590 |
| SLUG_02990 SLGD_00301 | SlsF | LPASG | 659 | 73.4 | Predicted cell-wall-anchored protein SasF, HKU09-01 |
| SLUG_03480 SLGD_00351 | SlsA | LPDTG | 1930 | 207.3 | Cell wall associated biofilm protein, HKU09-01, 3799 |
| SLUG_03490 SEVCU139_1800 SLGD_00352 | SlsD 4 | LPATG | 1619 | 175.8 | Putative serine-aspartate repeat protein F, VCU139, 2190 Putative uncharacterized protein, HKU09-01, 1136 |
| SLUG_03850 SLGD_00389 HMPREF0790_1688 | Slsc | LPETG | 190 | 21 | LPXTG protein, HKU09-01 Cell wall surface anchor family protein, M23590, 196 |
| SLUG_04710 SEVCU139_1680 SLGD_00473 | SlsE | LPETG | 3459 | 364 | Gram-positive signal peptide protein, VCU139, 2988 Hypothetical membrane protein, HKU09-01, 3232 |
| SLUG_04760 SLGD_00478 | SlsB | LPNTG | 277 | 30.6 | Putative uncharacterized protein, HKU09-01 |
| SLUG_22400 SLGD_02322 bca PE | SlsG | LPDTG | 2079 | 222.1 | Putative uncharacterized protein, HKU09-01, 2886 C protein alpha-antigen, VCU139, 2031 |
| SLUG_16350 HMPREF0790_0533 SLGD_01633 | Fbl | LPKTG | 881 | 94.2 | Clumping factor A, M23590, 857 Clumping factor A (fragment), VCU139, 688 Methicillin-resistant surface protein, HKU09-01, 701 |
| SLUG_23290 SLGD_02429 | vWbF | LPETG | 1869 | 209.4 | Von Willebrand factor-binding protein, HKU09-01, 2194 |
| Strain | fbl a | vWbF a |
|---|---|---|
| Mut47 vs. Sl48 | 1.88 ± 0.12 | 2.35 ± 0.25 |
| Mut7 vs. Sl44 | 1.99 ± 0.18 | 1.68 ± 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, M.; Kohler, C.; Becker, K. Role of SrtA in Pathogenicity of Staphylococcus lugdunensis. Microorganisms 2020, 8, 1975. https://doi.org/10.3390/microorganisms8121975
Hussain M, Kohler C, Becker K. Role of SrtA in Pathogenicity of Staphylococcus lugdunensis. Microorganisms. 2020; 8(12):1975. https://doi.org/10.3390/microorganisms8121975
Chicago/Turabian StyleHussain, Muzaffar, Christian Kohler, and Karsten Becker. 2020. "Role of SrtA in Pathogenicity of Staphylococcus lugdunensis" Microorganisms 8, no. 12: 1975. https://doi.org/10.3390/microorganisms8121975