Modulation of Antibacterial, Antioxidant, and Anti-Inflammatory Properties by Drying of Prunus domestica L. Plum Juice Extracts




Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Materials
2.3. Drying Procedure
2.4. Physicochemical Properties
2.5. Characterization of Polyphenolic Compounds by UPLC-PDA System
2.6. Bacterial Strains, Growth Media, and Culture Conditions
2.7. Antibacterial Activity
2.8. Cell Cultures
2.9. Cell Viability
2.10. Antioxidant Activity Against Intracellular Reactive Oxygen Species (ROS) Production
2.11. Anti-Inflammatory Activity
2.12. Statistical Analysis
3. Results and Discussion
3.1. Physical Properties
3.2. Polyphenolic Compounds Composition
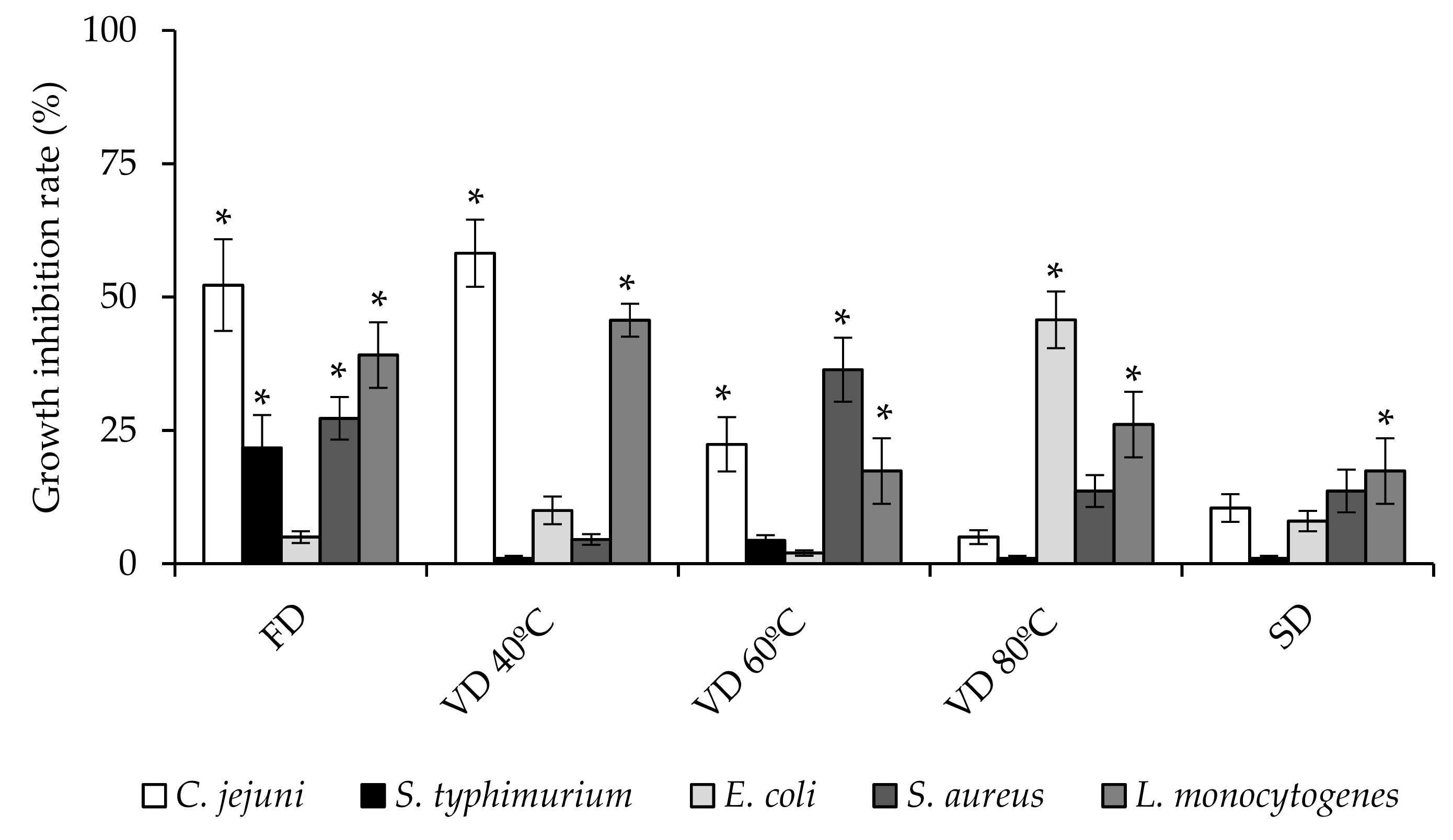
3.3. Antibacterial Activity
3.4. Antioxidant Activity Against Intracellular Reactive Oxygen Species (ROS) Production
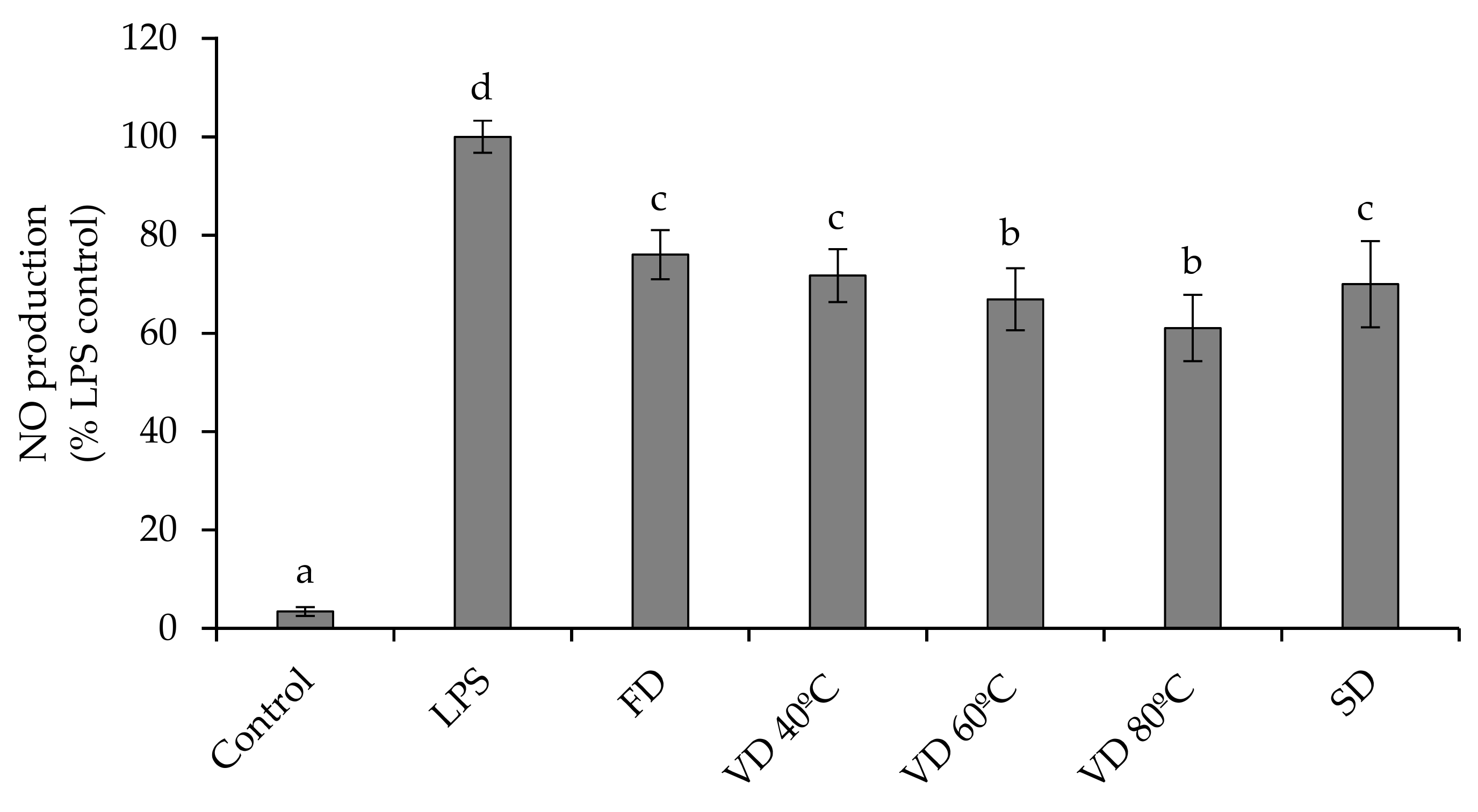
3.5. Anti-Inflammatory Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Noratto, G.; Porter, W.; Byrne, D.; Cisneros-Zevallos, L. Identifying peach and plum polyphenols with chemopreventive potential against estrogen-independent breast cancer cells. J. Agric. Food Chem. 2009, 57, 5219–5226. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization Corporate Statistical Database. Production. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 20 November 2019).
- Michalska, A.; Wojdyło, A.; Lech, K.; Łysiak, G.P.; Figiel, A. Physicochemical properties of whole fruit plum powders obtained using different drying technologies. Food Chem. 2016, 207, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Michalska, A.; Wojdyło, A.; Majerska, J.; Lech, K.; Brzezowska, J. Qualitative and quantitative evaluation of heat-induced changes in polyphenols and antioxidant capacity in Prunus domestica L. by-products. Molecules 2019, 24, 3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalska, A.; Wojdyło, A.; Łysiak, G.P.; Figiel, A. Chemical composition and antioxidant properties of powders obtained from different plum juice formulations. Int. J. Mol. Sci. 2017, 18, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figiel, A.; Michalska, A. Overall quality of fruits and vegetables products affected by the drying processes with the assistance of vacuum-microwaves. Int. J. Mol. Sci. 2017, 18, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karam, M.C.; Petit, J.; Zimmer, D.; Djantou, E.B.; Scher, J. Effects of drying and grinding in production of fruit and vegetable powders: A review. J. Food Eng. 2016, 188, 32–49. [Google Scholar] [CrossRef]
- Ratti, C. Hot air and freeze-drying of high-value foods: A review. J. Food Eng. 2001, 49, 311–319. [Google Scholar] [CrossRef]
- Lewicki, P.P. Design of hot air drying for better foods. Trends Food Sci. Technol. 2006, 17, 153–163. [Google Scholar] [CrossRef]
- Alamilla-Beltrán, L.; Chanona-Pérez, J.J.; Jiménez-Aparicio, A.R.; Gutiérrez-López, G.F. Description of morphological changes of particles along spray drying. J. Food Eng. 2005, 67, 179–184. [Google Scholar] [CrossRef]
- Horszwald, A.; Julien, H.; Andlauer, W. Characterisation of Aronia powders obtained by different drying processes. Food Chem. 2013, 141, 2858–2863. [Google Scholar] [CrossRef]
- Michalska, A.; Lech, K. The effect of carrier quantity and drying method on the physical properties of apple juice powders. Beverages 2018, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Majerska, J.; Michalska, A.; Figiel, A. A review of new directions in managing fruit and vegetable processing by-products. Trends Food Sci. Technol. 2019, 88, 207–219. [Google Scholar] [CrossRef]
- Sójka, M.; Kołodziejczyk, K.; Milala, J.; Abadias, M.; Viñas, I.; Guyot, S.; Baron, A. Composition and properties of the polyphenolic extracts obtained from industrial plum pomaces. J. Funct. Foods 2015, 12, 168–178. [Google Scholar] [CrossRef]
- Valtierra-Rodriguez, D.; Heredia, N.L.; Garcia, S.; Sanchez, E. Reduction of Campylobacter jejuni and Campylobacter coli in poultry skin by fruit extracts. J. Food Prot. 2010, 73, 477–482. [Google Scholar] [CrossRef]
- Khallouki, F.; Haubner, R.; Erben, G.; Ulrich, C.M.; Owen, R.W. Phytochemical composition and antioxidant capacity of various botanical parts of the fruits of Prunus domestica L. from the Lorraine region of Europe. Food Chem. 2012, 133, 697–706. [Google Scholar] [CrossRef]
- Kaulmann, A.; Legay, S.; Schneider, Y.J.; Hoffmann, L.; Bohn, T. Inflammation related responses of intestinal cells to plum and cabbage digesta with differential carotenoid and polyphenol profiles following simulated gastrointestinal digestion. Mol. Nutr. Food Res. 2016, 60, 992–1005. [Google Scholar] [CrossRef]
- Hooshmand, S.; Kumar, A.; Zhang, J.Y.; Johnson, S.A.; Chaid, S.C.; Arjmandi, B.H. Evidence for anti-inflammatory and antioxidative properties of dried plum polyphenols in macrophage RAW 264.7 cells. Food Funct. 2015, 6, 1719–1725. [Google Scholar] [CrossRef]
- Treutter, D.; Wang, D.; Farag, M.A.; Argueta Baires, G.D.; Ruhmann, S.; Neumuller, M. Diversity of phenolic profiles in the fruit skin of Prunus domestica plums and related species. J. Agric. Food Chem. 2012, 60, 12011–12019. [Google Scholar] [CrossRef]
- Figiel, A. Drying kinetics and quality of beetroots dehydrated by combination of convective and vacuum-microwave methods. J. Food Eng. 2010, 98, 461–470. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmianski, J.; Bielicki, P. Polyphenolic composition, antioxidant activity, and polyphenoloxidase (PPO) activity of quince (Cydonia oblonga Miller) varieties. J. Agric. Food Chem. 2013, 61, 2762–2772. [Google Scholar] [CrossRef]
- Silvan, J.M.; Mingo, E.; Hidalgo, M.; de Pascual-Teresa, S.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Antibacterial activity of a grape seed extract and its fractions against Campylobacter spp. Food Control 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Silvan, J.M.; Zorraquin-Peña, I.; Gonzalez de Llano, D.; Moreno-Arribas, M.V.; Martinez-Rodriguez, A.J. Antibacterial activity of glutathione-stabilized silver nanoparticles against Campylobacter multidrug-resistant strains. Front. Microbiol. 2018, 9, 458. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.A.; Cordero-Herrera, I.; Bravo, L.; Ramos, S.; Goya, L. Cocoa flavanols show beneficial effects in cultured pancreatic beta cells and liver cells to prevent the onset of type 2 diabetes. Food Res. Int. 2014, 63, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Boonyai, P.; Howes, T.; Bhandari, B. Applications of the cyclone stickiness test for characterization of stickiness in food powders. Dry. Technol. 2006, 24, 703–709. [Google Scholar] [CrossRef]
- Rahman, M.S.; Labuza, T.P. Water activity and food preservation. In Handbook of Food Preservation; Rahman, S., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 447–476. [Google Scholar]
- Michalska, A.; Wojdyło, A.; Honke, J.; Ciska, E.; Andlauer, W. Drying-induced physico-chemical changes in cranberry products. Food Chem. 2018, 240, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Martillanes, S.; Rocha-Pimienta, J.; Cabrera-Bañegil, M.; Martín-Vertedor, D.; Delgado-Adámez, J. Application of phenolic compounds for food preservation: Food additive and active packaging. In Phenolic Compounds Biological Activity; Soto-Hernández, M., Ed.; IntechOpen: London, UK, 2017; pp. 39–57. [Google Scholar]
- Ku, S.K.; Kwak, S.; Kwon, O.J.; Bae, J.S. Hyperoside inhibits high-glucose-induced vascular inflammation in vitro and in vivo. Inflammation 2014, 37, 1389–1400. [Google Scholar] [CrossRef]
- Xing, H.Y.; Liu, Y.; Chen, J.H.; Sun, F.J.; Shi, H.Q.; Xia, P.Y. Hyperoside attenuates hydrogen peroxide-induced L02 cell damage via MAPK-dependent Keap-Nrf-ARE signaling pathway. Biochem. Biophys. Commun. 2011, 410, 759–765. [Google Scholar] [CrossRef]
- Orhan, I.; Özçelik, B.; Kartal, M.; Özdeveci, B.; Duman, H. HPLC quantification of vitexine-2″-O-rhamnoside and hyperoside in three Crataegus species and their antimicrobial and antiviral activities. Chromatographia 2007, 66, 153–157. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, F.; Feng, W.; Qiu, X.; Liu, Y.; Yang, B.; Chen, Y.; Xia, P. Hyperoside inhibits biofilm formation of Pseudomonas aeruginosa. Exp. Ther. Med. 2017, 14, 1647–1652. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo Cavalcante de Araújo, D.; Diego da Silva, T.; Harand, W.; Sampaio de Andrade Lima, C.; Paulo Ferreira Neto, J.; de Azevedo Ramos, B.; Alves Rocha, T.; da Silva Alves, H.; Sobrinho de Sousa, R.; Paula de Oliveira, A.; et al. Bioguided purification of active compounds from leaves of Anadenanthera colubrina var. cebil (Griseb.) Altschul. Biomolecules 2019, 9, 590. [Google Scholar] [CrossRef] [Green Version]
- Fattouch, S.; Caboni, P.; Coroneo, V.; Tuberoso, C.I.; Angioni, A.; Dessi, S.; Marzouki, N.; Cabras, P. Antimicrobial activity of Tunisian quince (Cydonia oblonga Miller) pulp and peel polyphenolic extracts. J. Agric. Food Chem. 2007, 55, 963–969. [Google Scholar] [CrossRef]
- Park, S.F. The physiology of Campylobacter species and its relevance to their role as foodborne pathogens. Int. J. Food Microbiol. 2002, 74, 177–188. [Google Scholar] [CrossRef]
- Mingo, E.; Silvan, J.M.; Martinez-Rodriguez, A.J. Selective antibacterial effect on Campylobacter of a winemaking waste extract (WWE) as a source of active phenolic compounds. LWT Food Sci. Technol. 2016, 68, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.S.; Satsu, H.; Bae, M.J.; Totsuka, M.; Shimizu, M. Catechol groups enable reactive oxygen species scavenging-mediated suppression of PKD-NFkappaB-IL-8 signaling pathway by chlorogenic and caffeic acids in human intestinal cells. Nutrients 2018, 9, 165. [Google Scholar] [CrossRef]
- Bouayed, J.; Rammal, H.; Dicko, A.; Younos, C.; Soulimani, R. Chlorogenic acid, a polyphenol from Prunus domestica (Mirabelle), with coupled anxiolytic and antioxidant effects. J. Neurol. Sci. 2007, 262, 77–84. [Google Scholar] [CrossRef]
- Kim, H.; Hoon Pan, J.; Kim, S.H.; Lee, J.H.; Park, J.W. Chlorogenic acid ameliorates alcohol-induced liver injuries through scavenging reactive oxygen species. Biochimie 2018, 150, 131–138. [Google Scholar] [CrossRef]
- Liang, N.; Dupuis, J.H.; Yada, R.Y.; Kitts, D.D. Chlorogenic acid isomers directly interact with Keap 1-Nrf2 signalling in Caco-2 cells. Mol. Cell Biochem. 2019, 457, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Conforti, F.; Menichini, F. Phenolic compounds from plants as nitric oxide production inhibitors. Curr. Med. Chem. 2011, 18, 1137–1145. [Google Scholar] [CrossRef]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef]
- Banerjee, N.; Kim, H.; Talcott, S.T.; Turner, N.D.; Byrne, D.H.; Mertens-Talcott, S.U. Plum polyphenols inhibit colorectal aberrant crypt foci formation in rats: Potential role of the miR-143/protein kinase B/mammalian target of rapamycin axis. Nutr. Res. 2016, 36, 1105–1113. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Moisture Content (%) | Water Activity (-) | Colour | |||||
|---|---|---|---|---|---|---|---|
| L* | a* | b* | C* | h* | |||
| FD | 7.34 ± 0.14 d | 0.289 ± 0.001 d | 42.84 ± 0.13 b | 23.37 ± 0.1 d | 10.73 ± 0.03 d | 25.72 ± 0.10 c | 24.67 ± 0.13 b |
| VD 40 °C | 3.74 ± 0.17 c | 0.227 ± 0.002 b | 40.28 ± 0.05 a | 17.73 ± 0.04 c | 7.64 ± 0.01 b | 19.31 ± 0.03 b | 23.33 ± 0.05 a |
| VD 60 °C | 7.17 ± 0.04 d | 0.243 ± 0.001 c | 43.10 ± 0.67 b | 16.94 ± 0.61 b | 8.33 ± 0.10 c | 18.88 ± 0.59 b | 26.19 ± 0.56 c |
| VD 80 °C | 3.29 ± 0.08 b | 0.242 ± 0.001 c | 40.40 ± 0.09 a | 13.56 ± 0.02 a | 6.64 ± 0.02 a | 15.11 ± 0.03 a | 26.09 ± 0.07 c |
| SPD | 2.44 ± 0.06 a | 0.146 ± 0.002 a | 45.76 ± 0.02 c | 24.97 ± 0.06 e | 13.15 ± 0.03 e | 28.22 ± 0.04 d | 27.78 ± 0.09 d |
| Compound | FD | VD 40 °C | VD 60 °C | VD 80 °C | SPD |
|---|---|---|---|---|---|
| Phenolic acids | |||||
| Neochlorogenic acid | 10.25 ± 0.12 c | 10.18 ± 0.02 c | 8.85 ± 0.11 b | 6.45 ± 0.02 a | 11.04 ± 0.33 c |
| 3-feruoylquinic acid | 10.51 ± 0.06 c | 9.96 ± 0.11 c | 8.51 ± 0.36 b | 5.42 ± 0.15 a | 11.69 ± 0.28 d |
| 3-O-p-coumaroylquinic acid | 4.41 ± 0.09 ab | 4.73 ± 0.11 b | 4.59 ± 0.15 ab | 4.05 ± 0.21 a | 5.30 ± 0.14 c |
| Chlorogenic acid | 4.14 ± 0.01 c | 3.96 ± 0.01 bc | 3.58 ± 0.02 ab | 3.36 ± 0.12 a | 4.70 ± 0.04 d |
| Methyl 3-caffeoylquinate | 0.25 ± 0.01 a | 0.37 ± 0.01 ab | 0.55 ± 0.02 b | 7.08 ± 0.11 d | 3.92 ± 0.06 c |
| Total phenolic acids | 29.56 | 29.20 | 26.08 | 26.36 | 36.65 |
| Flavonols | |||||
| Quercetin-3-O-rutinoside | 1.25 ± 0.01 c | 0.09 ± 0.01 a | 0.24 ± 0.04 b | 1.09 ± 0.16 c | 1.88 ± 0.17 d |
| Quercetin-3-O-galactoside | 5.01 ± 0.05 d | 1.29 ± 0.01 b | 1.41 ± 0.01 c | 1.47 ± 0.06 c | 0.73 ± 0.04 a |
| Quercetin-3-O-glucoside | 0.73 ± 0.01 a | 5.24 ± 0.02 c | 5.14 ± 0.07 c | 4.58 ± 0.09 b | 6.11 ± 0.14 d |
| Quercetin-3-O-(6′′acetylgalactoside) | 1.03 ± 0.01 c | 1.04 ± 0.01 c | 0.89 ± 0.11 b | 0.61 ± 0.01 a | 1.21 ± 0.09 d |
| Total flavonols | 8.02 | 7.66 | 7.68 | 7.75 | 9.93 |
| Anthocyanins | |||||
| Cyanidin-3-O-glucoside | 0.27 ± 0.02 c | 0.26 ± 0.01 c | 0.24 ± 0.04 b | 0.17 ± 0.03 a | 0.28 ± 0.01 c |
| Cyanidin-3-O-rutinoside | 0.70 ± 0.01 bc | 0.70 ± 0.00 bc | 0.66 ± 0.11 ab | 0.53 ± 0.03 a | 0.78 ± 0.04 bc |
| Peonidin-3-O-rutinoside | 0.0058 ± 0.0001 b | 0.0058 ± 0.0002 b | 0.0055 ± 0.0009 b | 0.0044 ± 0.0007 a | 0.0065 ± 0.0003 c |
| Total anthocyanins | 0.97 | 0.96 | 0.90 | 0.70 | 1.07 |
| Total polyphenolics content | 38.55 | 37.82 | 34.66 | 34.81 | 47.65 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvan, J.M.; Michalska-Ciechanowska, A.; Martinez-Rodriguez, A.J. Modulation of Antibacterial, Antioxidant, and Anti-Inflammatory Properties by Drying of Prunus domestica L. Plum Juice Extracts. Microorganisms 2020, 8, 119. https://doi.org/10.3390/microorganisms8010119
Silvan JM, Michalska-Ciechanowska A, Martinez-Rodriguez AJ. Modulation of Antibacterial, Antioxidant, and Anti-Inflammatory Properties by Drying of Prunus domestica L. Plum Juice Extracts. Microorganisms. 2020; 8(1):119. https://doi.org/10.3390/microorganisms8010119
Chicago/Turabian StyleSilvan, Jose Manuel, Anna Michalska-Ciechanowska, and Adolfo J. Martinez-Rodriguez. 2020. "Modulation of Antibacterial, Antioxidant, and Anti-Inflammatory Properties by Drying of Prunus domestica L. Plum Juice Extracts" Microorganisms 8, no. 1: 119. https://doi.org/10.3390/microorganisms8010119