Broader Geographical Distribution of Toscana Virus in the Mediterranean Region Suggests the Existence of Larger Varieties of Sand Fly Vectors

 , and
, and {kind=link}
{kind=link}
Abstract
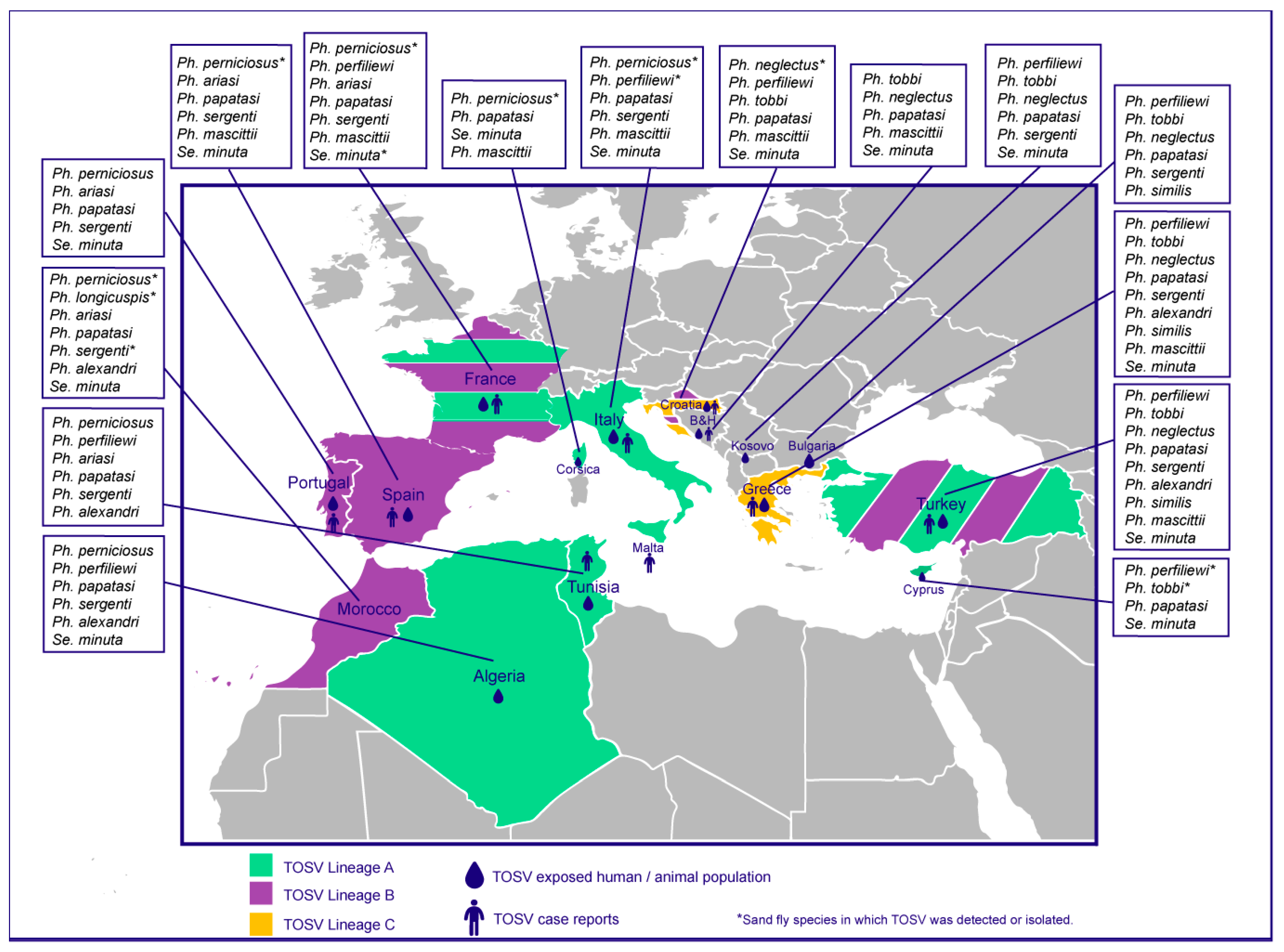
:1. Introduction
2. Toscana Virus
2.1. Overview on Toscana Virus
2.2. Epidemiology of Toscana Virus
2.3. Pathways of TOSV Transmission and Maintenance
3. Sand Flies
3.1. Overview on Sand Flies
3.2. Sandflies as TOSV Vectors
4. Vertebrates as Possible Reservoir or Amplification Host
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Maes, P.; Alkhovsky, S.V.; Bao, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the family Arenaviridae and the order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef] [Green Version]
- Charrel, R.; Bichaud, L.; De Lamballerie, X. Emergence of Toscana virus in the mediterranean area. World J. Virol. 2012, 1, 135. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Modi, G.B. Studies on the biology of phleboviruses in sand flies (Diptera: Psychodidae). I. Experimental infection of the vector. Am. J. Trop. Med. Hyg. 1984, 33, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2018). Arch. Virol. 2018, 163, 2601–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sanchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emerg. Infect. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Braito, A.; Corbisiero, R.; Corradini, S.; Marchi, B.; Sancasciani, N.; Fiorentini, C.; Ciufolini, M.G. Evidence of Toscana virus infections without central nervous system involvement: A serological study. Eur. J. Epidemiol. 1997, 13, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Schultze, D.; Korte, W.; Rafeiner, P.; Niedrig, M. First report of sandfly fever virus infection imported from Malta into Switzerland, October 2011. Euro. Surveill. 2012, 17. [Google Scholar] [CrossRef] [Green Version]
- Jaijakul, S.; Arias, C.A.; Hossain, M.; Arduino, R.C.; Wootton, S.H.; Hasbun, R. Toscana meningoencephalitis: A comparison to other viral central nervous system infections. J. Clin. Virol. 2012, 55, 204–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nougairede, A.; Bichaud, L.; Thiberville, S.D.; Ninove, L.; Zandotti, C.; de Lamballerie, X.; Brouqui, P.; Charrel, R.N. Isolation of Toscana virus from the cerebrospinal fluid of a man with meningitis in Marseille, France, 2010. Vector Borne Zoonotic Dis. 2013, 13, 685–688. [Google Scholar] [CrossRef]
- Karunaratne, K.; Davies, N. Toscana virus meningitis following a holiday in Elba, Italy. Br. J. Hosp. Med. 2018, 79, 292. [Google Scholar] [CrossRef]
- Echevarria, J.M.; de Ory, F.; Guisasola, M.E.; Sanchez-Seco, M.P.; Tenorio, A.; Lozano, A.; Cordoba, J.; Gobernado, M. Acute meningitis due to Toscana virus infection among patients from both the Spanish Mediterranean region and the region of Madrid. J. Clin. Virol. 2003, 26, 79–84. [Google Scholar] [CrossRef]
- Eitrem, R.; Vene, S.; Niklasson, B. Incidence of sand fly fever among Swedish United Nations soldiers on Cyprus during 1985. Am. J. Trop. Med. Hyg. 1990, 43, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Hemmersbach-Miller, M.; Parola, P.; Charrel, R.N.; Paul Durand, J.; Brouqui, P. Sandfly fever due to Toscana virus: An emerging infection in southern France. Eur. J. Intern. Med. 2004, 15, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.; Charrel, R.N. Emergent Sand Fly–Borne Phleboviruses in the Balkan Region. Emerg. Infect. Dis. 2018, 24, 2324–2330. [Google Scholar] [CrossRef]
- Alkan, C.; Allal-Ikhlef, A.B.; Alwassouf, S.; Baklouti, A.; Piorkowski, G.; de Lamballerie, X.; Izri, A.; Charrel, R.N. Virus isolation, genetic characterization and seroprevalence of Toscana virus in Algeria. Clin. Microbiol. Infect. 2015, 21, 1040. [Google Scholar] [CrossRef] [Green Version]
- Ayhan, N.; Charrel, R.N. Of phlebotomines (sandflies) and viruses: A comprehensive perspective on a complex situation. Curr. Opin. Insect Sci. 2017, 22, 117–124. [Google Scholar] [CrossRef]
- Charrel, R.N.; Izri, A.; Temmam, S.; Delaunay, P.; Toga, I.; Dumon, H.; Marty, P.; de Lamballerie, X.; Parola, P. Cocirculation of 2 genotypes of Toscana virus, southeastern France. Emerg. Infect. Dis. 2007, 13, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Ergunay, K.; Ayhan, N.; Charrel, R.N. Novel and emergent sandfly-borne phleboviruses in Asia Minor: A systematic review. Rev. Med. Virol. 2017, 27. [Google Scholar] [CrossRef]
- Ayhan, N.; Alten, B.; Ivovic, V.; Martinkovic, F.; Kasap, O.E.; Ozbel, Y.; de Lamballerie, X.; Charrel, R.N. Cocirculation of Two Lineages of Toscana Virus in Croatia. Front. Public Health 2017, 5, 336. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Peyrefitte, C.N.; Devetakov, I.; Pastorino, B.; Villeneuve, L.; Bessaud, M.; Stolidi, P.; Depaquit, J.; Segura, L.; Gravier, P.; Tock, F.; et al. Toscana virus and acute meningitis, France. Emerg. Infect. Dis. 2005, 11, 778–780. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Montero, J.; Gamez-Rueda, M.I.; Navarro-Mari, J.M.; de la Rosa-Fraile, M.; Oyonarte-Gomez, S. Infections due to sandfly fever virus serotype Toscana in Spain. Clin. Infect. Dis. 1998, 27, 434–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anagnostou, V.; Papa, A. Seroprevalence of Toscana virus among residents of Aegean Sea islands, Greece. Travel Med. Infect. Dis. 2013, 11, 98–102. [Google Scholar] [CrossRef] [PubMed]
- De Lamballerie, X.; Tolou, H.; Durand, J.P.; Charrel, R.N. Prevalence of Toscana virus antibodies in volunteer blood donors and patients with central nervous system infections in southeastern France. Vector Borne Zoonotic Dis. 2007, 7, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Eitrem, R.; Stylianou, M.; Niklasson, B. High prevalence rates of antibody to three sandfly fever viruses (Sicilian, Naples, and Toscana) among Cypriots. Epidemiol. Infect. 1991, 107, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Brisbarre, N.; Attoui, H.; Gallian, P.; Di Bonito, P.; Giorgi, C.; Cantaloube, J.F.; Biagini, P.; Touinssi, M.; Jordier, F.; de Micco, P. Seroprevalence of Toscana virus in blood donors, France, 2007. Emerg. Infect. Dis. 2011, 17, 941–943. [Google Scholar] [CrossRef]
- Ergunay, K.; Aydogan, S.; Ilhami Ozcebe, O.; Cilek, E.E.; Hacioglu, S.; Karakaya, J.; Ozkul, A.; Us, D. Toscana virus (TOSV) exposure is confirmed in blood donors from Central, North and South/Southeast Anatolia, Turkey. Zoonoses Public Health 2012, 59, 148–154. [Google Scholar] [CrossRef]
- Papa, A.; Andriotis, V.; Tzilianos, M. Prevalence of Toscana virus antibodies in residents of two Ionian islands, Greece. Travel Med. Infect. Dis. 2010, 8, 302–304. [Google Scholar] [CrossRef]
- Santos, L.; Cardoso, M.J.; Marinho, A.S.; Guimaraes, T.; Sarmento, A. [Seroprevalence survey of Toscana virus infection in Oporto region]. Acta Med. Port. 2011, 24, 479–482. [Google Scholar]
- Terrosi, C.; Olivieri, R.; Bianco, C.; Cellesi, C.; Cusi, M.G. Age-dependent seroprevalence of Toscana virus in central Italy and correlation with the clinical profile. Clin. Vaccine Immunol. 2009, 16, 1251–1252. [Google Scholar] [CrossRef] [Green Version]
- Punda-Polic, V.; Jeroncic, A.; Mohar, B.; Sisko Kraljevic, K. Prevalence of Toscana virus antibodies in residents of Croatia. Clin. Microbiol. Infect. 2012, 18, E200–E203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturi, G.; Marchi, A.; Fiorentini, C.; Ramadani, N.; Quaglio, G.; Kalaveshi, A.; Bertinato, L.; Putoto, G.; Benedetti, E.; Rezza, G.; et al. Prevalence of antibodies to phleboviruses and flaviviruses in Peja, Kosovo. Clin. Microbiol. Infect. 2011, 17, 1180–1182. [Google Scholar] [CrossRef] [Green Version]
- Hukic, M.; Salimovic-Besic, I. Sandfly—Pappataci fever in Bosnia and Herzegovina: The new-old disease. Bosn. J. Basic Med. Sci. 2009, 9, 39–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christova, I.; Panayotova, E.; Trifonova, I.; Taseva, E.; Gladnishka, T.; Ivanova, V. Serologic evidence of widespread Toscana virus infection in Bulgaria. J. Infect. Public Health 2019. [Google Scholar] [CrossRef] [PubMed]
- Fezaa, O.; Bahri, O.; Alaya Bouafif, N.B.; Triki, H.; Bouattour, A. Seroprevalence of Toscana virus infection in Tunisia. Int. J. Infect. Dis. 2013, 17, e1172–e1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakhria, S.; Bichaud, L.; Mensi, M.; Salez, N.; Dachraoui, K.; Thirion, L.; Cherni, S.; Chelbi, I.; De Lamballerie, X.; Zhioua, E.; et al. Co-circulation of Toscana virus and Punique virus in northern Tunisia: A microneutralisation-based seroprevalence study. PLoS Negl. Trop. Dis. 2013, 7, e2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesh, R.B.; Lubroth, J.; Guzman, H. Simulation of arbovirus overwintering: Survival of Toscana virus (Bunyaviridae: Phlebovirus) in its natural sand fly vector Phlebotomus perniciosus. Am. J. Trop. Med. Hyg. 1992, 47, 574–581. [Google Scholar] [CrossRef]
- Maroli, M.; Ciufolini, M.G.; Verani, P. Vertical transmission of Toscana virus in the sandfly, Phlebotomus perniciosus, via the second gonotrophic cycle. Med. Vet. Entomol. 1993, 7, 283–286. [Google Scholar] [CrossRef]
- Tesh, R.B.; Modi, G.B. Maintenance of Toscana virus in Phlebotomus perniciosus by vertical transmission. Am. J. Trop. Med. Hyg. 1987, 36, 189–193. [Google Scholar] [CrossRef]
- Peyrefitte, C.N.; Grandadam, M.; Bessaud, M.; Andry, P.-E.; Fouque, F.; Caro, V.; Diancourt, L.; Schuffenecker, I.; Pagès, F.; Tolou, H.; et al. Diversity of Phlebotomus perniciosus in Provence, Southeastern France: Detection of Two Putative New Phlebovirus Sequences. Vector Borne Zoonotic Dis. 2013, 13, 630–636. [Google Scholar] [CrossRef]
- Zhioua, E.; Moureau, G.; Chelbi, I.; Ninove, L.; Bichaud, L.; Derbali, M.; Champs, M.; Cherni, S.; Salez, N.; Cook, S.; et al. Punique virus, a novel phlebovirus, related to sandfly fever Naples virus, isolated from sandflies collected in Tunisia. J. Gen. Virol. 2010, 91, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Alwassouf, S.; Piorkowski, G.; Bichaud, L.; Tezcan, S.; Dincer, E.; Ergunay, K.; Ozbel, Y.; Alten, B.; de Lamballerie, X.; et al. Isolation, genetic characterization, and seroprevalence of Adana virus, a novel phlebovirus belonging to the Salehabad virus complex, in Turkey. J. Virol. 2015, 89, 4080–4091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remoli, M.E.; Fortuna, C.; Marchi, A.; Bucci, P.; Argentini, C.; Bongiorno, G.; Maroli, M.; Gradoni, L.; Gramiccia, M.; Ciufolini, M.G. Viral isolates of a novel putative phlebovirus in the Marche Region of Italy. Am. J. Trop. Med. Hyg. 2014, 90, 760–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jancarova, M.; Bichaud, L.; Hlavacova, J.; Priet, S.; Ayhan, N.; Spitzova, T.; Volf, P.; Charrel, R.N. Experimental infection of sand flies by Massilia virus and viral transmission by co-feeding on sugar meal. Viruses 2019, 11, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ready, P. Biology of Phlebotomine Sand Flies as Vectors of Disease Agents. Annu. Rev. Entomol. 2013, 58, 227–250. [Google Scholar] [CrossRef]
- Rueda, L.M.; Pecor, J.E.; Wolkoff, M.; Pecor, D.; Benyamin, S.; Bousses, P.; Debboun, M. New records, distribution, and updated checklists of old world Phlebotomine sand flies, with emphasis on Africa, southwest Asia, and central Asia. US Army Med. Dep. J. 2017, 1–17, 6–85. [Google Scholar]
- Abonnenc, E. Les Phlébotomes de la Région Éthiopienne (Diptera, Psychodidae); Service Central de Documentation de l’ORSTOM: Bondy, France, 1972; pp. 1–290. [Google Scholar]
- Muller, G.; Schlein, Y. Nectar and honeydew feeding of Phlebotomus papatasi in a focus of Leishmania major in Neot Hakikar oasis. J. Vector Ecol. 2004, 29, 154–158. [Google Scholar]
- Alten, B.; Maia, C.; Afonso, O.; Campino, L.; Jiménez, M.; González, E.; Molina, R.; Bañuls, A.L.; Prudhomme, J.; Vergnes, B.; et al. Seasonal dynamics of phlebotomine sand fly species proven vectors of Mediterranean leishmaniasis caused by Leishmania infantum. PLoS Negl. Trop. Dis. 2016, 10, e0004458. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- Lane, R. Sandflies (Phlebotominae). In Medical Insects and Arachnids; Springer: Dordrecht, Germany, 1993; pp. 78–119. [Google Scholar]
- Alten, B.; Ozbel, Y.; Ergunay, K.; Kasap, O.; Cull, B.; Antoniou, M.; Velo, E.; Prudhomme, J.; Molina, R.; Bañuls, A.; et al. Sampling strategies for phlebotomine sand flies (Diptera: Psychodidae) in Europe. Bull. Entomol. Res. 2015, 105, 664–678. [Google Scholar] [CrossRef]
- Fischer, D.; Moeller, P.; Thomas, S.; Naucke, T.; Beierkuhnlein, C. Combining Climatic Projections and Dispersal Ability: A Method for Estimating the Responses of Sandfly Vector Species to Climate Change. PLoS Negl. Trop. Dis. 2011, 5, e1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naucke, T.; Menn, B.; Massberg, D.; Lorentz, S. Sandflies and leishmaniasis in Germany. Parasitol. Res. 2008, 103, 65–68. [Google Scholar] [CrossRef]
- Verani, P.; Lopes, M.; Nicoletti, L.; Balducci, M. Studies on Phlebotomus-transmitted viruses in Italy. I. Isolation and characterization of a sandfly fever Naples-like virus. Arboviruses Mediterr. Ctries. Stuttg. Gustav Fischer Verl. 1980, Suppl. 9, 195–201. [Google Scholar]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.; Amaducci, L.; Coluzzi, M.; et al. Ecology of viruses isolated from sand flies in Italy and characterized of a new Phlebovirus (Arabia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Remoli, M.E.; Jimenez, M.; Fortuna, C.; Benedetti, E.; Marchi, A.; Genovese, D.; Gramiccia, M.; Molina, R.; Ciufolini, M.G. Phleboviruses detection in Phlebotomus perniciosus from a human leishmaniasis focus in South-West Madrid region, Spain. Parasit. Vectors 2016, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Es-Sette, N.; Nourlil, J.; Hamdi, S.; Mellouki, F.; Lemrani, M. First detection of Toscana virus RNA from sand flies in the genus Phlebotomus (Diptera: Phlebotomidae) naturally infected in Morocco. J. Med. Entomol. 2012, 49, 1507–1509. [Google Scholar] [CrossRef]
- Es-Sette, N.; Ajaoud, M.; Anga, L.; Mellouki, F.; Lemrani, M. Toscana virus isolated from sandflies, Morocco. Parasit. Vectors 2015, 8, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Charrel, R.N.; Izri, A.; Temmam, S.; de Lamballerie, X.; Parola, P. Toscana virus RNA in Sergentomyia minuta files. Emerg. Infect. Dis. 2006, 12, 1299–1300. [Google Scholar] [CrossRef]
- Ergunay, K.; Kasap, O.E.; Orsten, S.; Oter, K.; Gunay, F.; Yoldar, A.Z.; Dincer, E.; Alten, B.; Ozkul, A. Phlebovirus and Leishmania detection in sandflies from eastern Thrace and Northern Cyprus. Parasit. Vectors 2014, 7, 575. [Google Scholar] [CrossRef]
- Sanbonmatsu-Gamez, S.; Perez-Ruiz, M.; Collao, X.; Sanchez-Seco, M.P.; Morillas-Marquez, F.; de la Rosa-Fraile, M.; Navarro-Mari, J.M.; Tenorio, A. Toscana virus in Spain. Emerg. Infect. Dis. 2005, 11, 1701–1707. [Google Scholar] [CrossRef]
- Bichaud, L.; Izri, A.; Lamballerie, X.; Moureau, G.; Charrel, R. First detection of Toscana virus in Corsica, France. Clin. Microbiol. Infect. 2014, 20, O101–O104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bichaud, L.; Dachraoui, K.; Piorkowski, G.; Chelbi, I.; Moureau, G.; Cherni, S.; De Lamballerie, X.; Sakhria, S.; Charrel, R.N.; Zhioua, E. Toscana virus isolated from sandflies, Tunisia. Emerg. Infect. Dis. 2013, 19, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Chalghaf, B.; Chemkhi, J.; Mayala, B.; Harrabi, M.; Benie, G.B.; Michael, E.; Ben Salah, A. Ecological niche modeling predicting the potential distribution of Leishmania vectors in the Mediterranean basin: Impact of climate change. Parasit. Vectors 2018, 11, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Koch, L.K.; Kochmann, J.; Klimpel, S.; Cunze, S. Modeling the climatic suitability of leishmaniasis vector species in Europe. Sci. Rep. 2017, 7, 13325. [Google Scholar] [CrossRef] [PubMed]
- Mencke, N. The importance of canine leishmaniosis in non-endemic areas, with special emphasis on the situation in Germany. Berl. Munch. Tierarztl. Wochenschr. 2011, 124, 434–442. [Google Scholar]
- Depaquit, J.; Grandadam, M.; Fouque, F.; Andry, P.-E.; Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: A review. Euro. Surveill. 2010, 15, 19507. [Google Scholar]
- European Centre for Disease Prevention and Control. Phlebotomine Sand Flies Maps. Available online: https://www.ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/phlebotomine-maps (accessed on 1 October 2019).
- Senghor, M.W.; Niang, A.A.; Depaquit, J.; Ferte, H.; Faye, M.N.; Elguero, E.; Gaye, O.; Alten, B.; Perktas, U.; Cassan, C.; et al. Transmission of Leishmania infantum in the Canine Leishmaniasis Focus of Mont-Rolland, Senegal: Ecological, Parasitological and Molecular Evidence for a Possible Role of Sergentomyia Sand Flies. PLoS Negl. Trop. Dis. 2016, 10, e0004940. [Google Scholar] [CrossRef]
- Jaouadi, K.; Ghawar, W.; Salem, S.; Gharbi, M.; Bettaieb, J.; Yazidi, R.; Harrabi, M.; Hamarsheh, O.; Ben Salah, A. First report of naturally infected Sergentomyia minuta with Leishmania major in Tunisia. Parasit. Vectors 2015, 8, 649. [Google Scholar] [CrossRef] [Green Version]
- Maia, C.; Depaquit, J. Can Sergentomyia (Diptera, Psychodidae) play a role in the transmission of mammal-infecting Leishmania? Parasite 2016, 23, 55. [Google Scholar] [CrossRef] [Green Version]
- Bravo-Barriga, D.; Parreira, R.; Maia, C.; Afonso, M.O.; Blanco-Ciudad, J.; Serrano, F.J.; Perez-Martin, J.E.; Gomez-Gordo, L.; Campino, L.; Reina, D.; et al. Detection of Leishmania DNA and blood meal sources in phlebotomine sand flies (Diptera: Psychodidae) in western of Spain: Update on distribution and risk factors associated. Acta Trop. 2016, 164, 414–424. [Google Scholar] [CrossRef]
- Alwassouf, S.; Christodoulou, V.; Bichaud, L.; Ntais, P.; Mazeris, A.; Antoniou, M.; Charrel, R.N. Seroprevalence of Sandfly-Borne Phleboviruses Belonging to Three Serocomplexes (Sandfly fever Naples, Sandfly fever Sicilian and Salehabad) in Dogs from Greece and Cyprus Using Neutralization Test. PLoS Negl. Trop. Dis. 2016, 10, e0005063. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Mari, J.M.; Palop-Borras, B.; Perez-Ruiz, M.; Sanbonmatsu-Gamez, S. Serosurvey study of Toscana virus in domestic animals, Granada, Spain. Vector Borne Zoonotic Dis. 2011, 11, 583–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovao, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Alwassouf, S.; Cristovao, J.M.; Ayhan, N.; Pereira, A.; Charrel, R.N.; Campino, L. Serological association between Leishmania infantum and sand fly fever Sicilian (but not Toscana) virus in sheltered dogs from southern Portugal. Parasit. Vectors 2017, 10, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahmani, M.; Alwassouf, S.; Grech-Angelini, S.; Marie, J.L.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Corsica, France. Parasit. Vectors 2016, 9, 381. [Google Scholar] [CrossRef] [Green Version]
- Sakhria, S.; Alwassouf, S.; Fares, W.; Bichaud, L.; Dachraoui, K.; Alkan, C.; Zoghlami, Z.; De Lamballerie, X.; Zhioua, E.; Charrel, R.N. Presence of sandfly-borne phleboviruses of two antigenic complexes (Sandfly fever Naples virus and Sandfly fever Sicilian virus) in two different bio-geographical regions of Tunisia demonstrated by microneutralisation-based seroprevalence study in dogs. Parasit. Vectors 2014, 7, 476. [Google Scholar]
- Tahir, D.; Alwassouf, S.; Loudahi, A.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Kabylia (Algeria). Clin. Microbiol. Infect. 2016, 22, e16–e17. [Google Scholar] [CrossRef] [Green Version]
- Ayhan, N.; Sherifi, K.; Taraku, A.; Berxholi, K.; Charrel, R.N. High Rates of Neutralizing Antibodies to Toscana and Sandfly Fever Sicilian Viruses in Livestock, Kosovo. Emerg. Infect. Dis. 2017, 23, 989–992. [Google Scholar] [CrossRef]
- Dincer, E.; Karapinar, Z.; Oktem, M.; Ozbaba, M.; Ozkul, A.; Ergunay, K. Canine Infections and Partial S Segment Sequence Analysis of Toscana Virus in Turkey. Vector Borne Zoonotic Dis. 2016, 16, 611–618. [Google Scholar] [CrossRef]
- Dincer, E.; Gargari, S.; Ozkul, A.; Ergunay, K. Potential animal reservoirs of Toscana virus and coinfections with Leishmania infantum in Turkey. Am. J. Trop. Med. Hyg. 2015, 92, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Hacioglu, S.; Dincer, E.; Isler, C.T.; Karapinar, Z.; Ataseven, V.S.; Ozkul, A.; Ergunay, K. A Snapshot Avian Surveillance Reveals West Nile Virus and Evidence of Wild Birds Participating in Toscana Virus Circulation. Vector Borne Zoonotic Dis. 2017, 17, 698–708. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayhan, N.; Prudhomme, J.; Laroche, L.; Bañuls, A.-L.; Charrel, R.N. Broader Geographical Distribution of Toscana Virus in the Mediterranean Region Suggests the Existence of Larger Varieties of Sand Fly Vectors. Microorganisms 2020, 8, 114. https://doi.org/10.3390/microorganisms8010114
Ayhan N, Prudhomme J, Laroche L, Bañuls A-L, Charrel RN. Broader Geographical Distribution of Toscana Virus in the Mediterranean Region Suggests the Existence of Larger Varieties of Sand Fly Vectors. Microorganisms. 2020; 8(1):114. https://doi.org/10.3390/microorganisms8010114
Chicago/Turabian StyleAyhan, Nazli, Jorian Prudhomme, Lison Laroche, Anne-Laure Bañuls, and Remi N. Charrel. 2020. "Broader Geographical Distribution of Toscana Virus in the Mediterranean Region Suggests the Existence of Larger Varieties of Sand Fly Vectors" Microorganisms 8, no. 1: 114. https://doi.org/10.3390/microorganisms8010114