Rules and Exceptions: The Role of Chromosomal ParB in DNA Segregation and Other Cellular Processes


Abstract
:1. Introduction
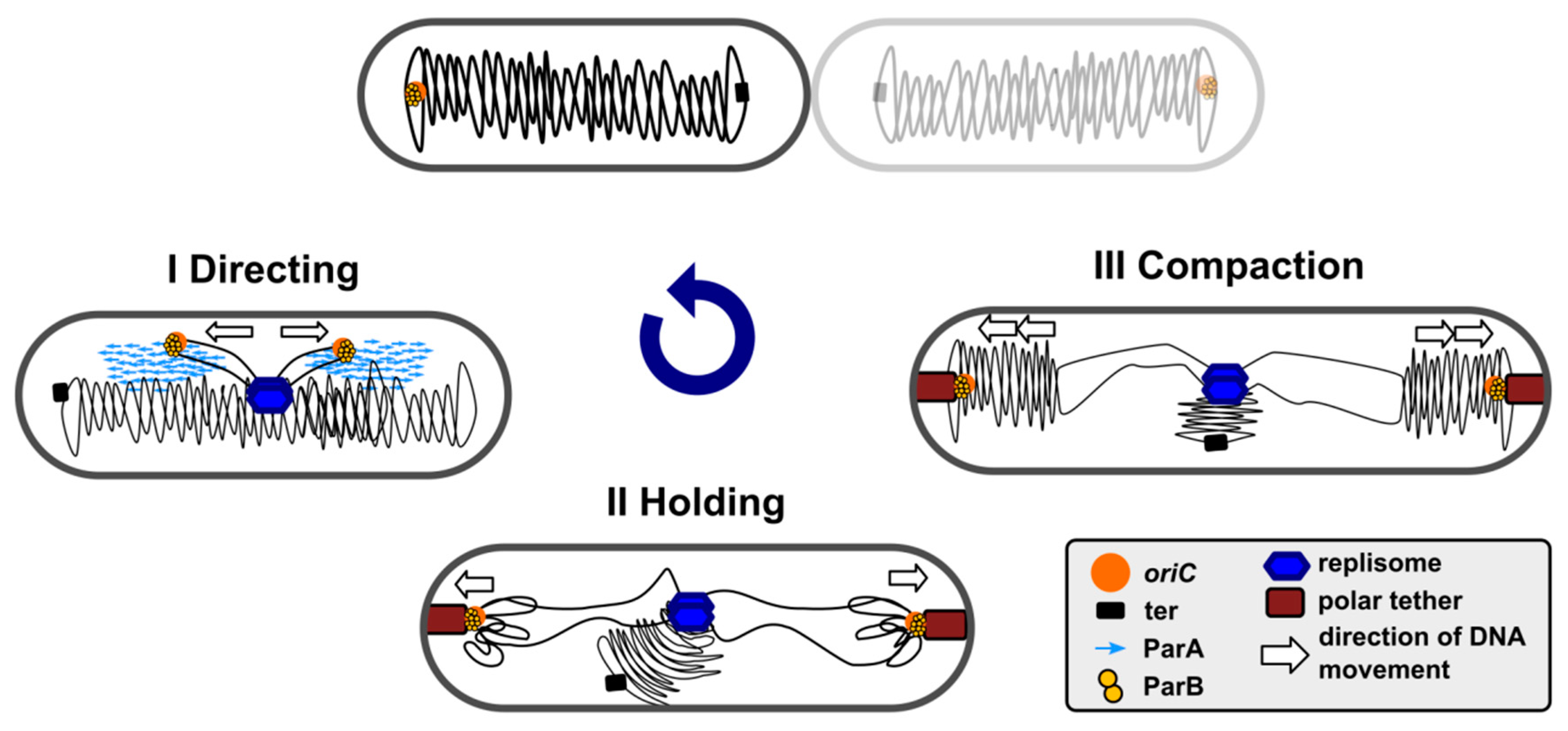
2. ParAB-parS-Driven DNA Segregation—From Plasmids to Chromosomes
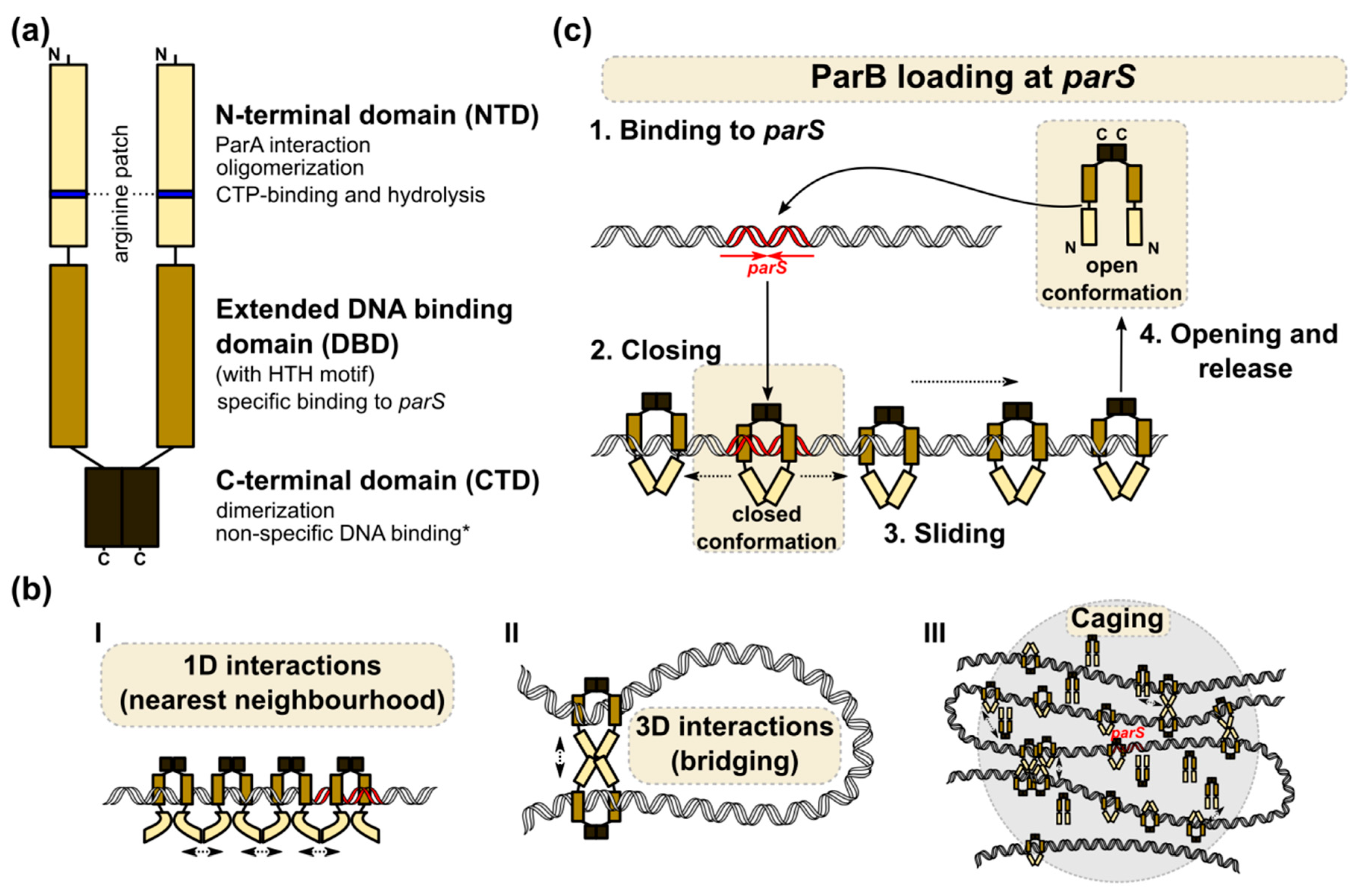
3. The Structure of the ParB–parS Complex
4. ParB Binding to Half-parS: A Novel Aspect of ParB–DNA Interactions?
5. The Role of ParBs in DNA Topology
6. The Role of ParB in the Regulation of Chromosome Replication Initiation
7. Involvement of ParB in The Nucleoid Occlusion
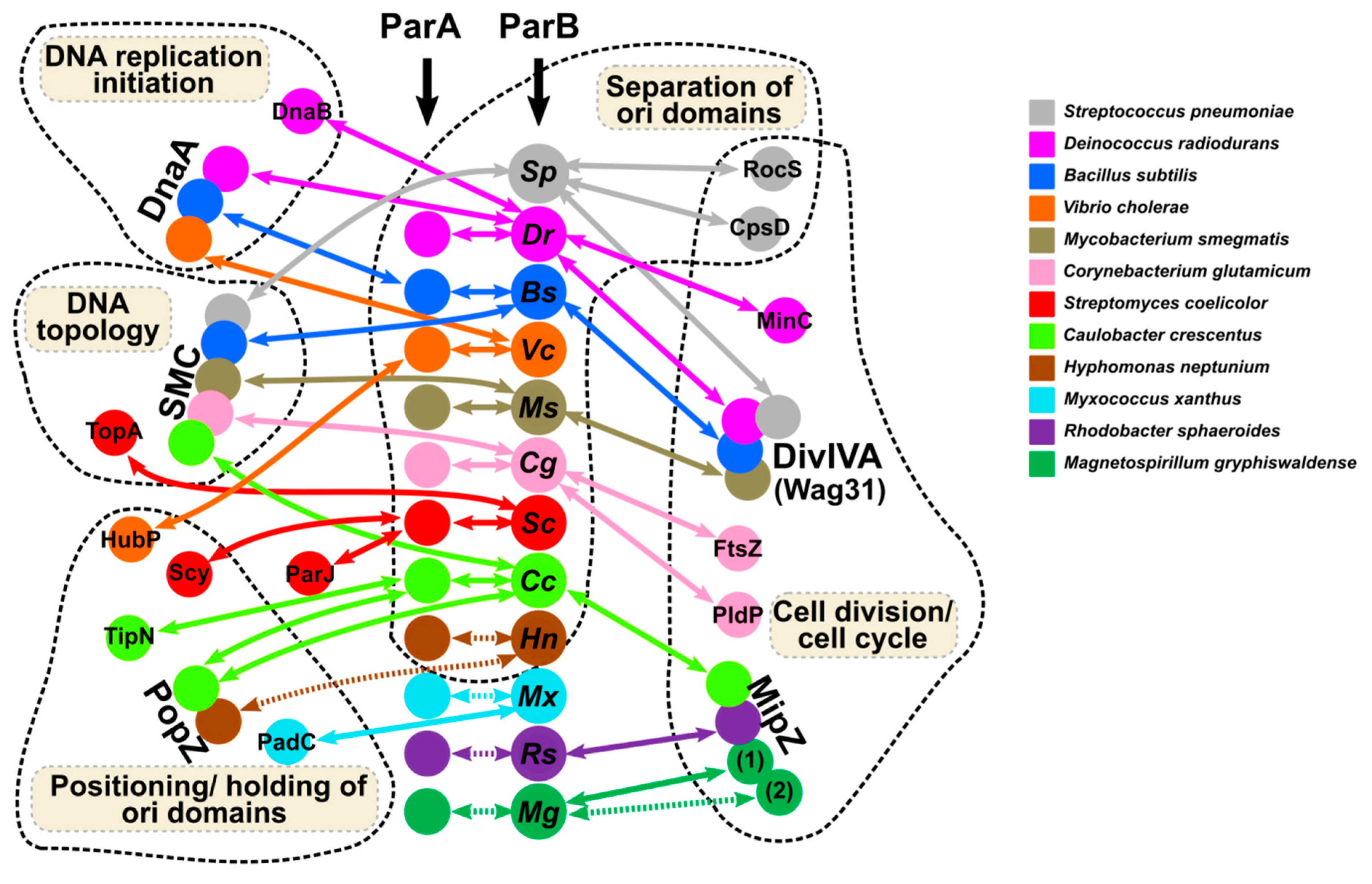
8. The Interactions of ParB with Topological Determinants during Cell Division
9. ParB’s Role in Capsule Formation in S. pneumoniae
10. Impact of ParBs on Gene Expression
11. Conclusions and Future Perspectives
Funding
Conflicts of Interest
References
- Dame, R.T.; Rashid, F.-Z.M.; Grainger, D.C. Chromosome organization in bacteria: Mechanistic insights into genome structure and function. Nat. Rev. Genet. 2019. [Google Scholar] [CrossRef] [PubMed]
- Badrinarayanan, A.; Le, T.B.; Laub, M.T. Bacterial chromosome organization and segregation. Annu. Rev. Cell Dev. Biol. 2015, 31, 171–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorman, C.J. Genome architecture and global gene regulation in bacteria: Making progress towards a unified model? Nat. Rev. Microbiol. 2013, 11, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T.; Tark-Dame, M. Bacterial chromatin: Converging views at different scales. Curr. Opin. Cell Biol. 2016, 40, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Gruber, S.; Errington, J. Recruitment of condensin to replication origin regions by ParB/Spo0J promotes chromosome segregation in B. subtilis. Cell 2009, 137, 685–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, N.L.; Marquis, K.A.; Rudner, D.Z. Recruitment of SMC by ParB-parS organizes the origin region and promotes efficient chromosome segregation. Cell 2009, 137, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.A.; Shapiro, L. An SMC ATPase mutant disrupts chromosome segregation in Caulobacter. Mol. Microbiol. 2011, 82, 1359–1374. [Google Scholar] [CrossRef] [Green Version]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef]
- Le, T.B.K.; Imakaev, M.V.; Mirny, L.A.; Laub, M.T. High-resolution mapping of the spatial organization of a bacterial chromosome. Science 2013, 342, 731–734. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Lamothe, R.; Sherratt, D.J. The bacterial cell cycle, chromosome inheritance and cell growth. Nat. Rev. Microbiol. 2019, 17, 467–478. [Google Scholar] [CrossRef]
- Wang, X.; Montero Llopis, P.; Rudner, D.Z. Organization and segregation of bacterial chromosomes. Nat. Rev. Genet. 2013, 14, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouet, J.-Y.; Stouf, M.; Lebailly, E.; Cornet, F. Mechanisms for chromosome segregation. Curr. Opin. Microbiol. 2014, 22, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Valens, M.; Penaud, S.; Rossignol, M.; Cornet, F.; Boccard, F. Macrodomain organization of the Escherichia coli chromosome. EMBO J. 2004, 23, 4330–4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dame, R.T.; Kalmykowa, O.J.; Grainger, D.C. Chromosomal macrodomains and associated proteins: Implications for DNA organization and replication in Gram-negative bacteria. PLoS Genet. 2011, 7, e1002123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surovtsev, I.V.; Jacobs-Wagner, C. Subcellular organization: A critical feature of bacterial cell replication. Cell 2018, 172, 1271–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marbouty, M.; Le Gall, A.; Cattoni, D.I.; Cournac, A.; Koh, A.; Fiche, J.-B.; Mozziconacci, J.; Murray, H.; Koszul, R.; Nollmann, M. Condensin- and replication-mediated bacterial chromosome folding and origin condensation revealed by Hi-C and super-resolution imaging. Mol. Cell 2015, 59, 588–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niki, H.; Yamaichi, Y.; Hiraga, S. Dynamic organization of chromosomal DNA in Escherichia coli. Genes Dev. 2000, 14, 212–223. [Google Scholar]
- Espeli, O.; Mercier, R.; Boccard, F. DNA dynamics vary according to macrodomain topography in the E. coli chromosome. Mol. Microbiol. 2008, 68, 1418–1427. [Google Scholar] [CrossRef]
- Dorman, C.J. Co-operative roles for DNA supercoiling and nucleoid-associated proteins in the regulation of bacterial transcription. Biochem. Soc. Trans. 2013, 41, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Val, M.-E.; Marbouty, M.; de Lemos Martins, F.; Kennedy, S.P.; Kemble, H.; Bland, M.J.; Possoz, C.; Koszul, R.; Skovgaard, O.; Mazel, D. A checkpoint control orchestrates the replication of the two chromosomes of Vibrio cholerae. Sci. Adv. 2016, 2, e1501914. [Google Scholar] [CrossRef] [Green Version]
- Higgins, N.P. Species-specific supercoil dynamics of the bacterial nucleoid. Biophys. Rev. 2016, 8, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.B.; Laub, M.T. Transcription rate and transcript length drive formation of chromosomal interaction domain boundaries. EMBO J. 2016, 35, 1582–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, D.F.; Grainger, D.C.; Busby, S.J. Effects of nucleoid-associated proteins on bacterial chromosome structure and gene expression. Curr. Opin. Microbiol. 2010, 13, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Lioy, V.S.; Cournac, A.; Marbouty, M.; Duigou, S.; Mozziconacci, J.; Espéli, O.; Boccard, F.; Koszul, R. Multiscale structuring of the E. coli chromosome by nucleoid-associated and condensin proteins. Cell 2018, 172, 771–783. [Google Scholar] [CrossRef]
- Danilova, O.; Reyes-Lamothe, R.; Pinskaya, M.; Sherratt, D.; Possoz, C. MukB colocalizes with the oriC region and is required for organization of the two Escherichia coli chromosome arms into separate cell halves. Mol. Microbiol. 2007, 65, 1485–1492. [Google Scholar] [CrossRef] [Green Version]
- Petrushenko, Z.M.; Cui, Y.; She, W.; Rybenkov, V.V. Mechanics of DNA bridging by bacterial condensin MukBEF in vitro and in singulo. EMBO J. 2010, 29, 1126–1135. [Google Scholar] [CrossRef]
- Strunnikov, A.V. SMC complexes in bacterial chromosome condensation and segregation. Plasmid 2006, 55, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Le, T.B.K.; Lajoie, B.R.; Dekker, J.; Laub, M.T.; Rudner, D.Z. Condensin promotes the juxtaposition of DNA flanking its loading site in Bacillus subtilis. Genes Dev. 2015, 29, 1661–1675. [Google Scholar] [CrossRef] [Green Version]
- Trussart, M.; Yus, E.; Martinez, S.; Baù, D.; Tahara, Y.O.; Pengo, T.; Widjaja, M.; Kretschmer, S.; Swoger, J.; Djordjevic, S.; et al. Defined chromosome structure in the genome-reduced bacterium Mycoplasma pneumoniae. Nat. Commun. 2017, 8, 14665. [Google Scholar] [CrossRef]
- Vallet-Gely, I.; Boccard, F. Chromosomal organization and segregation in Pseudomonas aeruginosa. PLoS Genet. 2013, 9, e1003492. [Google Scholar] [CrossRef] [Green Version]
- Harms, A.; Treuner-Lange, A.; Schumacher, D.; Søgaard-Andersen, L. Tracking of chromosome and replisome dynamics in Myxococcus xanthus reveals a novel chromosome arrangement. PLoS Genet. 2013, 9, e1003802. [Google Scholar] [CrossRef] [Green Version]
- Viollier, P.H.; Thanbichler, M.; McGrath, P.T.; West, L.; Meewan, M.; McAdams, H.H.; Shapiro, L. Rapid and sequential movement of individual chromosomal loci to specific subcellular locations during bacterial DNA replication. Proc. Natl. Acad. Sci. USA 2004, 101, 9257–9262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umbarger, M.A.; Toro, E.; Wright, M.A.; Porreca, G.J.; Baù, D.; Hong, S.-H.; Fero, M.J.; Zhu, L.J.; Marti-Renom, M.A.; McAdams, H.H.; et al. The three-dimensional architecture of a bacterial genome and its alteration by genetic perturbation. Mol. Cell 2011, 44, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Marbouty, M.; Cournac, A.; Flot, J.-F.; Marie-Nelly, H.; Mozziconacci, J.; Koszul, R. Metagenomic chromosome conformation capture (meta3C) unveils the diversity of chromosome organization in microorganisms. ELife 2014, 3, e03318. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.J.; Ottesen, J.R.; Youngren, B.; Austin, S.J.; Hansen, F.G. The Escherichia coli chromosome is organized with the left and right chromosome arms in separate cell halves. Mol. Microbiol. 2006, 62, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, X.; Possoz, C.; Sherratt, D.J. The two Escherichia coli chromosome arms locate to separate cell halves. Genes Dev. 2006, 20, 1727–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordström, K.; Molin, S.; Aagaard-Hansen, H. Partitioning of plasmid R1 in Escherichia coli: I. Kinetics of loss of plasmid derivatives deleted of the par region. Plasmid 1980, 4, 215–227. [Google Scholar] [CrossRef]
- Austin, S.; Abeles, A. Partition of unit-copy miniplasmids to daughter cells. II. The partition region of miniplasmid P1 encodes an essential protein and a centromere-like site at which it acts. J. Mol. Biol. 1983, 169, 373–387. [Google Scholar] [CrossRef]
- Abeles, A.L.; Friedman, S.A.; Austin, S.J. Partition of unit-copy miniplasmids to daughter cells. III. The DNA sequence and functional organization of the P1 partition region. J. Mol. Biol. 1985, 185, 261–272. [Google Scholar] [CrossRef]
- Gerdes, K.; Larsen, J.E.; Molin, S. Stable inheritance of plasmid R1 requires two different loci. J. Bacteriol. 1985, 161, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, K.; Møller-Jensen, J.; Jensen, R.B. Plasmid and chromosome partitioning: Surprises from phylogeny. Mol. Microbiol. 2000, 37, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Howard, M.; Szardenings, F. Pushing and pulling in prokaryotic DNA segregation. Cell 2010, 141, 927–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, F. The partition system of multidrug resistance plasmid TP228 includes a novel protein that epitomizes an evolutionarily distinct subgroup of the ParA superfamily. Mol. Microbiol. 2000, 37, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Motallebi-Veshareh, M.; Rouch, D.A.; Thomas, C.M. A family of ATPases involved in active partitioning of diverse bacterial plasmids. Mol. Microbiol. 1990, 4, 1455–1463. [Google Scholar] [CrossRef]
- Koonin, E.V. A superfamily of ATPases with diverse functions containing either classical or deviant ATP-binding motif. J. Mol. Biol. 1993, 229, 1165–1174. [Google Scholar] [CrossRef]
- Møller-Jensen, J.; Borch, J.; Dam, M.; Jensen, R.B.; Roepstorff, P.; Gerdes, K. Bacterial mitosis: ParM of plasmid R1 moves plasmid DNA by an actin-like insertional polymerization mechanism. Mol. Cell 2003, 12, 1477–1487. [Google Scholar] [CrossRef]
- Larsen, R.A.; Cusumano, C.; Fujioka, A.; Lim-Fong, G.; Patterson, P.; Pogliano, J. Treadmilling of a prokaryotic tubulin-like protein, TubZ, required for plasmid stability in Bacillus thuringiensis. Genes Dev. 2007, 21, 1340–1352. [Google Scholar] [CrossRef] [Green Version]
- Delbrück, H.; Ziegelin, G.; Lanka, E.; Heinemann, U. An Src homology 3-like domain is responsible for dimerization of the repressor protein KorB encoded by the promiscuous IncP plasmid RP4. J. Biol. Chem. 2002, 277, 4191–4198. [Google Scholar] [CrossRef] [Green Version]
- Khare, D.; Ziegelin, G.; Lanka, E.; Heinemann, U. Sequence-specific DNA binding determined by contacts outside the helix-turn-helix motif of the ParB homolog KorB. Nat. Struct. Mol. Biol. 2004, 11, 656–663. [Google Scholar] [CrossRef]
- Schumacher, M.A.; Mansoor, A.; Funnell, B.E. Structure of a four-way bridged ParB-DNA complex provides insight into P1 segrosome assembly. J. Biol. Chem. 2007, 282, 10456–10464. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, M.A.; Piro, K.M.; Xu, W. Insight into F plasmid DNA segregation revealed by structures of SopB and SopB-DNA complexes. Nucleic Acids Res. 2010, 38, 4514–4526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golovanov, A.P.; Barillà, D.; Golovanova, M.; Hayes, F.; Lian, L.-Y. ParG, a protein required for active partition of bacterial plasmids, has a dimeric ribbon-helix-helix structure. Mol. Microbiol. 2003, 50, 1141–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murayama, K.; Orth, P.; de la Hoz, A.B.; Alonso, J.C.; Saenger, W. Crystal structure of omega transcriptional repressor encoded by Streptococcus pyogenes plasmid pSM19035 at 1.5 A resolution. J. Mol. Biol. 2001, 314, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Barillà, D.; Carmelo, E.; Hayes, F. The tail of the ParG DNA segregation protein remodels ParF polymers and enhances ATP hydrolysis via an arginine finger-like motif. Proc. Natl. Acad. Sci. USA 2007, 104, 1811–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, L.; Xu, W.; Kumaraswami, M.; Schumacher, M.A. Plasmid protein TubR uses a distinct mode of HTH-DNA binding and recruits the prokaryotic tubulin homolog TubZ to effect DNA partition. Proc. Natl. Acad. Sci. USA 2010, 107, 11763–11768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, J.C.; Funnell, B.E. Plasmid partition mechanisms in Plasmids. In Biology and Impact in Biotechnology and Discovery; Tolmasky, M.E., Alonso, J.A., Eds.; ASM Press: Washington, DC, USA, 2015; pp. 135–155. [Google Scholar]
- Schumacher, M.A. Bacterial plasmid partition machinery: A minimalist approach to survival. Curr. Opin. Struct. Biol. 2012, 22, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, N.; Yoshikawa, H. Genes and their organization in the replication origin region of the bacterial chromosome. Mol. Microbiol. 1992, 6, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Livny, J.; Yamaichi, Y.; Waldor, M.K. Distribution of centromere-like parS sites in bacteria: Insights from comparative genomics. J. Bacteriol. 2007, 189, 8693–8703. [Google Scholar] [CrossRef] [Green Version]
- Gal-Mor, O.; Borovok, I.; Av-Gay, Y.; Cohen, G.; Aharonowitz, Y. Gene organization in the trxA/B-oriC region of the Streptomyces coelicolor chromosome and comparison with other eubacteria. Gene 1998, 217, 83–90. [Google Scholar] [CrossRef]
- Lin, D.C.; Grossman, A.D. Identification and characterization of a bacterial chromosome partitioning site. Cell 1998, 92, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Yamaichi, Y.; Niki, H. Active segregation by the Bacillus subtilis partitioning system in Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 14656–14661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrin-Estevenon, A.-M.; Pasta, F.; Lane, D. The parAB gene products of Pseudomonas putida exhibit partition activity in both P. putida and Escherichia coli. Mol. Microbiol. 2002, 43, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Bartosik, A.A.; Lasocki, K.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. ParB of Pseudomonas aeruginosa: Interactions with its partner ParA and its target parS and specific effects on bacterial growth. J. Bacteriol. 2004, 186, 6983–6998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemon, K.P.; Grossman, A.D. Localization of bacterial DNA polymerase: Evidence for a factory model of replication. Science 1998, 282, 1516–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawitzke, J.; Austin, S. An analysis of the factory model for chromosome replication and segregation in bacteria. Mol. Microbiol. 2001, 40, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Bartosik, A.A.; Jagura-Burdzy, G. Bacterial chromosome segregation. Acta Biochim. Pol. 2005, 52, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Breier, A.M.; Grossman, A.D. Whole-genome analysis of the chromosome partitioning and sporulation protein Spo0J (ParB) reveals spreading and origin-distal sites on the Bacillus subtilis chromosome. Mol. Microbiol. 2007, 64, 703–718. [Google Scholar] [CrossRef]
- Ireton, K.; Gunther, N.W.; Grossman, A.D. spo0J is required for normal chromosome segregation as well as the initiation of sporulation in Bacillus subtilis. J. Bacteriol. 1994, 176, 5320–5329. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.S.; Grossman, A.D. The chromosome partitioning proteins Soj (ParA) and Spo0J (ParB) contribute to accurate chromosome partitioning, separation of replicated sister origins, and regulation of replication initiation in Bacillus subtilis. Mol. Microbiol. 2006, 60, 853–869. [Google Scholar] [CrossRef]
- Minnen, A.; Attaiech, L.; Thon, M.; Gruber, S.; Veening, J.-W. SMC is recruited to oriC by ParB and promotes chromosome segregation in Streptococcus pneumoniae. Mol. Microbiol. 2011, 81, 676–688. [Google Scholar] [CrossRef] [Green Version]
- Böhm, K.; Giacomelli, G.; Schmidt, A.; Imhof, A.; Koszul, R.; Marbouty, M.; Bramkamp, M. Chromosome organization by a conserved condensin-ParB system in the actinobacterium Corynebacterium glutamicum. bioRxiv 2019. [Google Scholar] [CrossRef]
- Donovan, C.; Schwaiger, A.; Krämer, R.; Bramkamp, M. Subcellular localization and characterization of the ParAB system from Corynebacterium glutamicum. J. Bacteriol. 2010, 192, 3441–3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakimowicz, D.; Brzostek, A.; Rumijowska-Galewicz, A.; Zydek, P.; Dołzbłasz, A.; Smulczyk-Krawczyszyn, A.; Zimniak, T.; Wojtasz, L.; Zawilak-Pawlik, A.; Kois, A.; et al. Characterization of the mycobacterial chromosome segregation protein ParB and identification of its target in Mycobacterium smegmatis. Microbiol. Read. Engl. 2007, 153, 4050–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santi, I.; McKinney, J.D. Chromosome organization and replisome dynamics in Mycobacterium smegmatis. mBio 2015, 6, e01999-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakimowicz, D.; Chater, K.; Zakrzewska-Czerwínska, J. The ParB protein of Streptomyces coelicolor A3(2) recognizes a cluster of parS sequences within the origin-proximal region of the linear chromosome. Mol. Microbiol. 2002, 45, 1365–1377. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Calcutt, M.J.; Schmidt, F.J.; Chater, K.F. Partitioning of the linear chromosome during sporulation of Streptomyces coelicolor A3(2) involves an oriC-linked parAB locus. J. Bacteriol. 2000, 182, 1313–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakimowicz, D.; Zydek, P.; Kois, A.; Zakrzewska-Czerwińska, J.; Chater, K.F. Alignment of multiple chromosomes along helical ParA scaffolding in sporulating Streptomyces hyphae. Mol. Microbiol. 2007, 65, 625–641. [Google Scholar] [CrossRef]
- Donczew, M.; Mackiewicz, P.; Wróbel, A.; Flärdh, K.; Zakrzewska-Czerwińska, J.; Jakimowicz, D. ParA and ParB coordinate chromosome segregation with cell elongation and division during Streptomyces sporulation. Open Biol. 2016, 6, 150263. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.T.; Stevenson, C.E.; Som, N.F.; Thanapipatsiri, A.; Jalal, A.S.B.; Le, T.B.K. Permissive zones for the centromere-binding protein ParB on the Caulobacter crescentus chromosome. Nucleic Acids Res. 2018, 46, 1196–1209. [Google Scholar] [CrossRef] [Green Version]
- Toro, E.; Hong, S.-H.; McAdams, H.H.; Shapiro, L. Caulobacter requires a dedicated mechanism to initiate chromosome segregation. Proc. Natl. Acad. Sci. USA 2008, 105, 15435–15440. [Google Scholar] [CrossRef] [Green Version]
- Mohl, D.A.; Easter, J.; Gober, J.W. The chromosome partitioning protein, ParB, is required for cytokinesis in Caulobacter crescentus. Mol. Microbiol. 2001, 42, 741–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, A.; Raßbach, A.; Pulpetta, R.L.; van Teeseling, M.C.F.; Heinrich, K.; Sobetzko, P.; Serrania, J.; Becker, A.; Thanbichler, M. Two-step chromosome segregation in the stalked budding bacterium Hyphomonas neptunium. Nat. Commun. 2019, 10, 3290. [Google Scholar] [CrossRef] [PubMed]
- Dubarry, N.; Pasta, F.; Lane, D. ParABS systems of the four replicons of Burkholderia cenocepacia: New chromosome centromeres confer partition specificity. J. Bacteriol. 2006, 188, 1489–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.-L.; Dubarry, N.; Passot, F.M.; Kamgoué, A.; Murray, H.; Lane, D.; Pasta, F. Orderly Replication and segregation of the four replicons of Burkholderia cenocepacia J2315. PLoS Genet. 2016, 12, e1006172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passot, F.M.; Calderon, V.; Fichant, G.; Lane, D.; Pasta, F. Centromere binding and evolution of chromosomal partition systems in the Burkholderiales. J. Bacteriol. 2012, 194, 3426–3436. [Google Scholar] [CrossRef] [Green Version]
- Pillet, F.; Passot, F.M.; Pasta, F.; Anton Leberre, V.; Bouet, J.-Y. Analysis of ParB-centromere interactions by multiplex SPR imaging reveals specific patterns for binding ParB in six centromeres of Burkholderiales chromosomes and plasmids. PLoS ONE 2017, 12, e0177056. [Google Scholar] [CrossRef]
- Iniesta, A.A. ParABS system in chromosome partitioning in the bacterium Myxococcus xanthus. PLoS ONE 2014, 9, e86897. [Google Scholar] [CrossRef] [Green Version]
- Lagage, V.; Boccard, F.; Vallet-Gely, I. Regional control of chromosome segregation in Pseudomonas aeruginosa. PLoS Genet. 2016, 12, e1006428. [Google Scholar] [CrossRef]
- Kawalek, A.; Bartosik, A.A.; Glabski, K.; Jagura-Burdzy, G. Pseudomonas aeruginosa partitioning protein ParB acts as a nucleoid-associated protein binding to multiple copies of a parS-related motif. Nucleic Acids Res. 2018, 46, 4592–4606. [Google Scholar] [CrossRef] [Green Version]
- Jecz, P.; Bartosik, A.A.; Glabski, K.; Jagura-Burdzy, G. A single parS sequence from the cluster of four sites closest to oriC is necessary and sufficient for proper chromosome segregation in Pseudomonas aeruginosa. PLoS ONE 2015, 10, e0120867. [Google Scholar] [CrossRef] [Green Version]
- Bartosik, A.A.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. ParB deficiency in Pseudomonas aeruginosa destabilizes the partner protein ParA and affects a variety of physiological parameters. Microbiology 2009, 155, 1080–1092. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.A.; Bignell, C.R.; Zeng, W.; Jones, A.C.; Thomas, C.M. Chromosome loss from par mutants of Pseudomonas putida depends on growth medium and phase of growth. Microbiology 2002, 148, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Yamaichi, Y.; Fogel, M.A.; McLeod, S.M.; Hui, M.P.; Waldor, M.K. Distinct centromere-like parS sites on the two chromosomes of Vibrio spp. J. Bacteriol. 2007, 189, 5314–5324. [Google Scholar] [CrossRef] [Green Version]
- Kadoya, R.; Baek, J.H.; Sarker, A.; Chattoraj, D.K. Participation of chromosome segregation protein ParAI of Vibrio cholerae in chromosome replication. J. Bacteriol. 2011, 193, 1504–1514. [Google Scholar] [CrossRef] [Green Version]
- Charaka, V.K.; Misra, H.S. Functional characterization of the role of the chromosome I partitioning system in genome segregation in Deinococcus radiodurans. J. Bacteriol. 2012, 194, 5739–5748. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Angelov, A.; Pham, V.T.T.; Leis, B.; Liebl, W. Characterization of chromosomal and megaplasmid partitioning loci in Thermus thermophilus HB27. BMC Genomics 2015, 16, 317. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Random chromosome partitioning in the polyploid bacterium Thermus thermophilus HB27. G3 Genes Genomes Genet. 2019, 9, 1249–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Mori, Y.; Ichinose, C.; Niki, H.; Ogura, T.; Kato, A.; Hiraga, S. Purification and characterization of SopA and SopB proteins essential for F plasmid partitioning. J. Biol. Chem. 1989, 264, 15535–15541. [Google Scholar] [PubMed]
- Hayes, F.; Radnedge, L.; Davis, M.A.; Austin, S.J. The homologous operons for P1 and P7 plasmid partition are autoregulated from dissimilar operator sites. Mol. Microbiol. 1994, 11, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Hester, C.M.; Lutkenhaus, J. Soj (ParA) DNA binding is mediated by conserved arginines and is essential for plasmid segregation. Proc. Natl. Acad. Sci. USA 2007, 104, 20326–20331. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.-H.; Yen, C.-Y.; Chen, B.-W.; Lin, M.-G.; Wang, L.-H.; Tang, K.-Z.; Hsiao, C.-D.; Sun, Y.-J. Crystal structures of HpSoj–DNA complexes and the nucleoid-adaptor complex formation in chromosome segregation. Nucleic Acids Res. 2019, 47, 2113–2129. [Google Scholar] [CrossRef] [Green Version]
- Ptacin, J.L.; Lee, S.F.; Garner, E.C.; Toro, E.; Eckart, M.; Comolli, L.R.; Moerner, W.E.; Shapiro, L. A spindle-like apparatus guides bacterial chromosome segregation. Nat. Cell Biol. 2010, 12, 791–798. [Google Scholar] [CrossRef]
- Figge, R.M.; Easter, J.; Gober, J.W. Productive interaction between the chromosome partitioning proteins, ParA and ParB, is required for the progression of the cell cycle in Caulobacter crescentus. Mol. Microbiol. 2003, 47, 1225–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosik, A.A.; Glabski, K.; Jecz, P.; Lasocki, K.; Mikosa, M.; Plochocka, D.; Thomas, C.M.; Jagura-Burdzy, G. Dissection of the region of Pseudomonas aeruginosa ParA that is important for dimerization and interactions with its partner ParB. Microbiology 2014, 160, 2406–2420. [Google Scholar] [CrossRef] [PubMed]
- Ringgaard, S.; Zon, J.; van Howard, M.; Gerdes, K. Movement and equipositioning of plasmids by ParA filament disassembly. Proc. Natl. Acad. Sci. USA 2009, 106, 19369–19374. [Google Scholar] [CrossRef] [Green Version]
- Fogel, M.A.; Waldor, M.K. A dynamic, mitotic-like mechanism for bacterial chromosome segregation. Genes Dev. 2006, 20, 3269–3282. [Google Scholar] [CrossRef] [Green Version]
- Hui, M.P.; Galkin, V.E.; Yu, X.; Stasiak, A.Z.; Stasiak, A.; Waldor, M.K.; Egelman, E.H. ParA2, a Vibrio cholerae chromosome partitioning protein, forms left-handed helical filaments on DNA. Proc. Natl. Acad. Sci. USA 2010, 107, 4590–4595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.C.; Surovtsev, I.V.; Beltran, B.G.; Huang, F.; Bewersdorf, J.; Jacobs-Wagner, C. Evidence for a DNA-relay mechanism in ParABS-mediated chromosome segregation. Elife 2014, 3, e02758. [Google Scholar] [CrossRef] [PubMed]
- Vecchiarelli, A.G.; Mizuuchi, K.; Funnell, B.E. Surfing biological surfaces: Exploiting the nucleoid for partition and transport in bacteria. Mol. Microbiol. 2012, 86, 513–523. [Google Scholar] [CrossRef]
- Hwang, L.C.; Vecchiarelli, A.G.; Han, Y.-W.; Mizuuchi, M.; Harada, Y.; Funnell, B.E.; Mizuuchi, K. ParA-mediated plasmid partition driven by protein pattern self-organization. EMBO J. 2013, 32, 1238–1249. [Google Scholar] [CrossRef] [Green Version]
- Kisner, J.R.; Kuwada, N.J. Nucleoid-mediated positioning and transport in bacteria. Curr. Genet. 2019. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.C.; Hwang, L.C. Reconstitutions of plasmid partition systems and their mechanisms. Plasmid 2017, 91, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusiak, M.; Gapczyńska, A.; Płochocka, D.; Thomas, C.M.; Jagura-Burdzy, G. Binding and spreading of ParB on DNA determine its biological function in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 3342–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, T.A.; Butler, P.J.G.; Löwe, J. Structural analysis of the chromosome segregation protein Spo0J from Thermus thermophilus. Mol. Microbiol. 2004, 53, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-W.; Lin, M.-H.; Chu, C.-H.; Hsu, C.-E.; Sun, Y.-J. Insights into ParB spreading from the complex structure of Spo0J and parS. Proc. Natl. Acad. Sci. USA 2015, 112, 6613–6618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, T.G.W.; Wang, X.; Song, D.; Etson, C.M.; van Oijen, A.M.; Rudner, D.Z.; Loparo, J.J. ParB spreading requires DNA bridging. Genes Dev. 2014, 28, 1228–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funnell, B.E. ParB partition proteins: Complex formation and spreading at bacterial and plasmid centromeres. Front. Mol. Biosci. 2016, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Fisher, G.L.; Pastrana, C.L.; Higman, V.A.; Koh, A.; Taylor, J.A.; Butterer, A.; Craggs, T.; Sobott, F.; Murray, H.; Crump, M.P.; et al. The structural basis for dynamic DNA binding and bridging interactions which condense the bacterial centromere. ELife 2017, 6, e28086. [Google Scholar] [CrossRef]
- Taylor, J.A.; Pastrana, C.L.; Butterer, A.; Pernstich, C.; Gwynn, E.J.; Sobott, F.; Moreno-Herrero, F.; Dillingham, M.S. Specific and non-specific interactions of ParB with DNA: Implications for chromosome segregation. Nucleic Acids Res. 2015, 43, 719–731. [Google Scholar] [CrossRef]
- Madariaga-Marcos, J.; Pastrana, C.L.; Fisher, G.L.; Dillingham, M.S.; Moreno-Herrero, F. ParB dynamics and the critical role of the CTD in DNA condensation unveiled by combined force-fluorescence measurements. ELife 2019, 8, e43812. [Google Scholar] [CrossRef]
- Soh, Y.-M.; Davidson, I.F.; Zamuner, S.; Basquin, J.; Bock, F.P.; Taschner, M.; Veening, J.-W.; Rios, P.D.L.; Peters, J.-M.; Gruber, S. Self-organization of parS centromeres by the ParB CTP hydrolase. Science 2019, 366, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Lynch, A.S.; Wang, J.C. SopB protein-mediated silencing of genes linked to the sopC locus of Escherichia coli F plasmid. Proc. Natl. Acad. Sci. USA 1995, 92, 1896–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodionov, O.; Łobocka, M.; Yarmolinsky, M. Silencing of genes flanking the P1 plasmid centromere. Science 1999, 283, 546–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukaszewicz, M.; Kostelidou, K.; Bartosik, A.A.; Cooke, G.D.; Thomas, C.M.; Jagura-Burdzy, G. Functional dissection of the ParB homologue (KorB) from IncP-1 plasmid RK2. Nucleic Acids Res. 2002, 30, 1046–1055. [Google Scholar] [CrossRef]
- Bingle, L.E.H.; Macartney, D.P.; Fantozzi, A.; Manzoor, S.E.; Thomas, C.M. Flexibility in repression and cooperativity by KorB of broad host range IncP-1 plasmid RK2. J. Mol. Biol. 2005, 349, 302–316. [Google Scholar] [CrossRef]
- Mohl, D.A.; Gober, J.W. Cell cycle-dependent polar localization of chromosome partitioning proteins in Caulobacter crescentus. Cell 1997, 88, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.C.-H.; Levin, P.A.; Grossman, A.D. Bipolar localization of a chromosome partition protein in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 1997, 94, 4721–4726. [Google Scholar] [CrossRef] [Green Version]
- Debaugny, R.E.; Sanchez, A.; Rech, J.; Labourdette, D.; Dorignac, J.; Geniet, F.; Palmeri, J.; Parmeggiani, A.; Boudsocq, F.; Anton Leberre, V.; et al. A conserved mechanism drives partition complex assembly on bacterial chromosomes and plasmids. Mol. Syst. Biol. 2018, 14, e8516. [Google Scholar] [CrossRef]
- Attaiech, L.; Minnen, A.; Kjos, M.; Gruber, S.; Veening, J.-W. The ParB-parS chromosome segregation system modulates competence development in Streptococcus pneumoniae. mBio 2015, 6, e00662. [Google Scholar] [CrossRef] [Green Version]
- Glaser, P.; Sharpe, M.E.; Raether, B.; Perego, M.; Ohlsen, K.; Errington, J. Dynamic, mitotic-like behavior of a bacterial protein required for accurate chromosome partitioning. Genes Dev. 1997, 11, 1160–1168. [Google Scholar] [CrossRef] [Green Version]
- Baek, J.H.; Rajagopala, S.V.; Chattoraj, D.K. Chromosome segregation proteins of Vibrio cholerae as transcription regulators. mBio 2014, 5, e01061-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, A.S.B.; Pastrana, C.L.; Tran, N.T.; Stevenson, C.E.; Lawson, D.M.; Moreno-Herrero, F.; Le, T.B.K. Structural and biochemical analyses of Caulobacter crescentus ParB reveal the role of its N-terminal domain in chromosome segregation. bioRxiv 2019, 816959. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Rodrigues, K.; Graham, T.G.W.; Loparo, J.J. A network of cis and trans interactions is required for ParB spreading. Nucleic Acids Res. 2017, 45, 7106–7117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, B.N.; Dean, R. The evidence of large-scale DNA-induced compaction in the Mycobacterial chromosomal ParB. J. Mol. Biol. 2011, 413, 901–907. [Google Scholar] [CrossRef]
- Broedersz, C.P.; Wang, X.; Meir, Y.; Loparo, J.J.; Rudner, D.Z.; Wingreen, N.S. Condensation and localization of the partitioning protein ParB on the bacterial chromosome. Proc. Natl. Acad. Sci. USA 2014, 111, 8809–8814. [Google Scholar] [CrossRef] [Green Version]
- Audibert, S.; Gac, N.T.-L.; Rech, J.; Turlan, C.; Bouet, J.-Y.; Bystricky, K.; Lane, D. Role of centromere sites in activation of ParB proteins for partition complex assembly. bioRxiv 2019, 862136. [Google Scholar] [CrossRef]
- Walter, J.-C.; Walliser, N.-O.; David, G.; Dorignac, J.; Geniet, F.; Palmeri, J.; Parmeggiani, A.; Wingreen, N.S.; Broedersz, C.P. Looping and clustering model for the organization of protein-DNA complexes on the bacterial genome. N. J. Phys. 2018, 20, e035002. [Google Scholar] [CrossRef]
- Sanchez, A.; Cattoni, D.I.; Walter, J.-C.; Rech, J.; Parmeggiani, A.; Nollmann, M.; Bouet, J.-Y. Stochastic Self-Assembly of ParB proteins builds the bacterial DNA segregation apparatus. Cell Syst. 2015, 1, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Jalal, A.S.B.; Tran, N.T.; Le, T.B.K. ParB spreading on DNA requires cytidine triphosphate in vitro. bioRxiv 2019, 865972. [Google Scholar] [CrossRef] [Green Version]
- Osorio-Valeriano, M.; Altegoer, F.; Steinchen, W.; Urban, S.; Liu, Y.; Bange, G.; Thanbichler, M. ParB-type DNA segregation proteins are CTP-dependent molecular switches. Cell 2019, 179, 1512–1524.e15. [Google Scholar] [CrossRef]
- Jalal, A.S.B.; Tran, N.T.; Stevenson, C.E.; Tan, X.; Lawson, D.M.; Le, T.B.K. Evolving a new protein-DNA interface via sequential introduction of permissive and specificity-switching mutations. bioRxiv 2019, 724823. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinforma. Oxf. Engl. 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, F.; Ryan, D.P.; Grüning, B.; Bhardwaj, V.; Kilpert, F.; Richter, A.S.; Heyne, S.; Dündar, F.; Manke, T. deepTools2: A next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 2016, 44, W160–W165. [Google Scholar] [CrossRef] [PubMed]
- Yuen, K.C.; Gerton, J.L. Taking cohesin and condensin in context. PLoS Genet. 2018, 14, e1007118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnen, A.; Bürmann, F.; Wilhelm, L.; Anchimiuk, A.; Diebold-Durand, M.-L.; Gruber, S. Control of Smc coiled coil architecture by the ATPase heads facilitates targeting to chromosomal ParB/parS and release onto flanking DNA. Cell Rep. 2016, 14, 2003–2016. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Brandão, H.B.; Le, T.B.K.; Laub, M.T.; Rudner, D.Z. Bacillus subtilis SMC complexes juxtapose chromosome arms as they travel from origin to terminus. Science 2017, 355, 524–527. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T. At the heart of the chromosome: SMC proteins in action. Nat. Rev. Mol. Cell Biol. 2006, 7, 311–322. [Google Scholar] [CrossRef]
- Tran, N.T.; Laub, M.T.; Le, T.B.K. SMC Progressively aligns chromosomal arms in Caulobacter crescentus but is antagonized by convergent transcription. Cell Rep. 2017, 20, 2057–2071. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, L.; Bürmann, F.; Minnen, A.; Shin, H.-C.; Toseland, C.P.; Oh, B.-H.; Gruber, S. SMC condensin entraps chromosomal DNA by an ATP hydrolysis dependent loading mechanism in Bacillus subtilis. eLife 2015, 4, e06659. [Google Scholar] [CrossRef]
- Hassler, M.; Shaltiel, I.A.; Kschonsak, M.; Simon, B.; Merkel, F.; Thärichen, L.; Bailey, H.J.; Macošek, J.; Bravo, S.; Metz, J.; et al. Structural basis of an asymmetric condensin ATPase cycle. Mol. Cell 2019, 74, 1175–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, S. SMC complexes sweeping through the chromosome: Going with the flow and against the tide. Curr. Opin. Microbiol. 2018, 42, 96–103. [Google Scholar] [CrossRef]
- Nolivos, S.; Upton, A.L.; Badrinarayanan, A.; Müller, J.; Zawadzka, K.; Wiktor, J.; Gill, A.; Arciszewska, L.; Nicolas, E.; Sherratt, D. MatP regulates the coordinated action of topoisomerase IV and MukBEF in chromosome segregation. Nat. Commun. 2016, 7, 10466. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Clevenger, A.L.; Ritchey, J.W.; Zgurskaya, H.I.; Rybenkov, V.V. Pseudomonas aeruginosa condensins support opposite differentiation states. J. Bacteriol. 2016, 198, 2936–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panas, M.W.; Jain, P.; Yang, H.; Mitra, S.; Biswas, D.; Wattam, A.R.; Letvin, N.L.; Jacobs, W.R. Noncanonical SMC protein in Mycobacterium smegmatis restricts maintenance of Mycobacterium fortuitum plasmids. Proc. Natl. Acad. Sci. USA 2014, 111, 13264–13271. [Google Scholar] [CrossRef] [Green Version]
- Szafran, M.; Skut, P.; Ditkowski, B.; Ginda, K.; Chandra, G.; Zakrzewska-Czerwińska, J.; Jakimowicz, D. Topoisomerase I (TopA) is recruited to ParB complexes and is required for proper chromosome organization during Streptomyces coelicolor sporulation. J. Bacteriol. 2013, 195, 4445–4455. [Google Scholar] [CrossRef] [Green Version]
- Champoux, J.J. DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef] [Green Version]
- Murray, H.; Errington, J. Dynamic control of the DNA replication initiation protein DnaA by Soj/ParA. Cell 2008, 135, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Scholefield, G.; Whiting, R.; Errington, J.; Murray, H. Spo0J regulates the oligomeric state of Soj to trigger its switch from an activator to an inhibitor of DNA replication initiation. Mol. Microbiol. 2011, 79, 1089–1100. [Google Scholar] [CrossRef]
- Scholefield, G.; Errington, J.; Murray, H. Soj/ParA stalls DNA replication by inhibiting helix formation of the initiator protein DnaA. EMBO J. 2012, 31, 1542–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, G.K.; Kota, S.; Misra, H.S. Characterisation of ParB encoded on multipartite genome in Deinococcus radiodurans and their roles in radioresistance. Microbiol. Res. 2019, 223, 22–32. [Google Scholar] [CrossRef]
- Perry, S.E.; Edwards, D.H. The Bacillus subtilis DivIVA protein has a sporulation-specific proximity to Spo0J. J. Bacteriol. 2006, 188, 6039–6043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Osorio Valeriano, M.; Harms, A.; Søgaard-Andersen, L.; Thanbichler, M. Bactofilin-mediated organization of the ParABS chromosome segregation system in Myxococcus xanthus. Nat. Commun. 2017, 8, 1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginda, K.; Bezulska, M.; Ziółkiewicz, M.; Dziadek, J.; Zakrzewska-Czerwińska, J.; Jakimowicz, D. ParA of Mycobacterium smegmatis co-ordinates chromosome segregation with the cell cycle and interacts with the polar growth determinant DivIVA. Mol. Microbiol. 2013, 87, 998–1012. [Google Scholar] [CrossRef]
- Pióro, M.; Małecki, T.; Portas, M.; Magierowska, I.; Trojanowski, D.; Sherratt, D.; Zakrzewska-Czerwińska, J.; Ginda, K.; Jakimowicz, D. Competition between DivIVA and the nucleoid for ParA binding promotes segrosome separation and modulates mycobacterial cell elongation. Mol. Microbiol. 2019, 111, 204–220. [Google Scholar] [CrossRef]
- Nourikyan, J.; Kjos, M.; Mercy, C.; Cluzel, C.; Morlot, C.; Noirot-Gros, M.-F.; Guiral, S.; Lavergne, J.-P.; Veening, J.-W.; Grangeasse, C. Autophosphorylation of the bacterial tyrosine-kinase CpsD connects capsule synthesis with the cell cycle in Streptococcus pneumoniae. PLoS Genet. 2015, 11, e1005518. [Google Scholar] [CrossRef] [Green Version]
- Fadda, D.; Santona, A.; D’Ulisse, V.; Ghelardini, P.; Ennas, M.G.; Whalen, M.B.; Massidda, O. Streptococcus pneumoniae DivIVA: Localization and interactions in a MinCD-free context. J. Bacteriol. 2007, 189, 1288–1298. [Google Scholar] [CrossRef] [Green Version]
- Mercy, C.; Ducret, A.; Slager, J.; Lavergne, J.-P.; Freton, C.; Nagarajan, S.N.; Garcia, P.S.; Noirot-Gros, M.-F.; Dubarry, N.; Nourikyan, J.; et al. RocS drives chromosome segregation and nucleoid protection in Streptococcus pneumoniae. Nat. Microbiol. 2019, 4, 1661–1670. [Google Scholar] [CrossRef]
- Kois-Ostrowska, A.; Strzałka, A.; Lipietta, N.; Tilley, E.; Zakrzewska-Czerwińska, J.; Herron, P.; Jakimowicz, D. Unique function of the bacterial chromosome segregation machinery in apically growing Streptomyces—targeting the chromosome to new hyphal tubes and its anchorage at the tips. PLoS Genet. 2016, 12, e1006488. [Google Scholar] [CrossRef]
- Ditkowski, B.; Troć, P.; Ginda, K.; Donczew, M.; Chater, K.F.; Zakrzewska-Czerwińska, J.; Jakimowicz, D. The actinobacterial signature protein ParJ (SCO1662) regulates ParA polymerization and affects chromosome segregation and cell division during Streptomyces sporulation. Mol. Microbiol. 2010, 78, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Ditkowski, B.; Holmes, N.; Rydzak, J.; Donczew, M.; Bezulska, M.; Ginda, K.; Kedzierski, P.; Zakrzewska-Czerwińska, J.; Kelemen, G.H.; Jakimowicz, D. Dynamic interplay of ParA with the polarity protein, Scy, coordinates the growth with chromosome segregation in Streptomyces coelicolor. Open Biol. 2013, 3, 130006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shebelut, C.W.; Guberman, J.M.; Teeffelen, S.; van Yakhnina, A.A.; Gitai, Z. Caulobacter chromosome segregation is an ordered multistep process. Proc. Natl. Acad. Sci. USA 2010, 107, 14194–14198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, G.R.; Comolli, L.R.; Zhu, J.; Eckart, M.; Koenig, M.; Downing, K.H.; Moerner, W.E.; Earnest, T.; Shapiro, L. A polymeric protein anchors the chromosomal origin/ParB complex at a bacterial cell pole. Cell 2008, 134, 945–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ptacin, J.L.; Gahlmann, A.; Bowman, G.R.; Perez, A.M.; von Diezmann, A.R.S.; Eckart, R.M.; Moerner, W.E.; Shapiro, L. Bacterial scaffold directs pole-specific centromere segregation. Proc. Natl. Acad. Sci. USA 2014, 111, 2046–2055. [Google Scholar] [CrossRef] [Green Version]
- Thanbichler, M.; Shapiro, L. MipZ, A spatial regulator coordinating chromosome segregation with cell division in Caulobacter. Cell 2006, 126, 147–162. [Google Scholar] [CrossRef] [Green Version]
- Schofield, W.B.; Lim, H.C.; Jacobs-Wagner, C. Cell cycle coordination and regulation of bacterial chromosome segregation dynamics by polarly localized proteins. EMBO J. 2010, 29, 3068–3081. [Google Scholar] [CrossRef]
- Charaka, V.; Kota, S.; Misra, H. ParA encoded on chromosome i of Deinococcus radiodurans requires its cognate ParB and centromere for its dynamics. Proc. Indian Natl. Sci. Acad. 2014, 80, 663. [Google Scholar] [CrossRef]
- Maurya, G.K.; Modi, K.; Misra, H.S. Divisome and segrosome components of Deinococcus radiodurans interact through cell division regulatory proteins. Microbiol. Read. Engl. 2016, 162, 1321–1334. [Google Scholar] [CrossRef]
- Chaudhary, R.; Gupta, A.; Kota, S.; Misra, H.S. N-terminal domain of DivIVA contributes to its dimerization and interaction with genome segregation proteins in a radioresistant bacterium Deinococcus radiodurans. Int. J. Biol. Macromol. 2019, 128, 12–21. [Google Scholar] [CrossRef]
- Yamaichi, Y.; Bruckner, R.; Ringgaard, S.; Möll, A.; Cameron, D.E.; Briegel, A.; Jensen, G.J.; Davis, B.M.; Waldor, M.K. A multidomain hub anchors the chromosome segregation and chemotactic machinery to the bacterial pole. Genes Dev. 2012, 26, 2348–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubarry, N.; Willis, C.R.; Ball, G.; Lesterlin, C.; Armitage, J.P. In vivo imaging of the segregation of the 2 chromosomes and the cell division proteins of Rhodobacter sphaeroides reveals an unexpected role for MipZ. mBio 2019, 10, e02515–e02518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toro-Nahuelpan, M.; Corrales-Guerrero, L.; Zwiener, T.; Osorio-Valeriano, M.; Müller, F.-D.; Plitzko, J.M.; Bramkamp, M.; Thanbichler, M.; Schüler, D. A gradient-forming MipZ protein mediating the control of cell division in the magnetotactic bacterium Magnetospirillum gryphiswaldense. Mol. Microbiol. 2019, 112, 1423–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errington, J.; Wu, L.J. Cell cycle machinery in Bacillus subtilis. Subcell. Biochem. 2017, 84, 67–101. [Google Scholar] [PubMed] [Green Version]
- van Baarle, S.; Bramkamp, M. The MinCDJ system in Bacillus subtilis prevents minicell formation by promoting divisome disassembly. PLoS ONE 2010, 5, e9850. [Google Scholar] [CrossRef] [Green Version]
- Sievers, J.; Raether, B.; Perego, M.; Errington, J. Characterization of the parB-like yyaA gene of Bacillus subtilis. J. Bacteriol. 2002, 184, 1102–1111. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.J.; Ishikawa, S.; Kawai, Y.; Oshima, T.; Ogasawara, N.; Errington, J. Noc protein binds to specific DNA sequences to coordinate cell division with chromosome segregation. EMBO J. 2009, 28, 1940–1952. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.W.; Wu, L.J.; Errington, J. Nucleoid occlusion protein Noc recruits DNA to the bacterial cell membrane. EMBO J. 2015, 34, 491–501. [Google Scholar] [CrossRef]
- Hajduk, I.V.; Mann, R.; Rodrigues, C.D.A.; Harry, E.J. The ParB homologs, Spo0J and Noc, together prevent premature midcell Z ring assembly when the early stages of replication are blocked in Bacillus subtilis. Mol. Microbiol. 2019, 112, 766–784. [Google Scholar] [CrossRef]
- Hammond, L.R.; White, M.L.; Eswara, P.J. ¡vIVA la DivIVA! J. Bacteriol. 2019, 201, e00245-19. [Google Scholar] [CrossRef]
- Bergé, M.; Viollier, P.H. End-in-sight: Cell polarization by the polygamic organizer PopZ. Trends Microbiol. 2018, 26, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Marczynski, G.T.; Petit, K.; Patel, P. Crosstalk regulation between bacterial chromosome replication and chromosome partitioning. Front. Microbiol. 2019, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Ausmees, N.; Kuhn, J.R.; Jacobs-Wagner, C. The bacterial cytoskeleton: An intermediate filament-like function in cell shape. Cell 2003, 115, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Refes, Y.; He, B.; Corrales-Guerrero, L.; Steinchen, W.; Panis, G.; Viollier, P.H.; Bange, G.; Thanbichler, M. Molecular architecture of the DNA-binding sites of the P-loop ATPases MipZ and ParA from Caulobacter crescentus. bioRxiv 2019, 766287. [Google Scholar] [CrossRef]
- Kiekebusch, D.; Michie, K.A.; Essen, L.-O.; Löwe, J.; Thanbichler, M. Localized dimerization and nucleoid binding drive gradient formation by the bacterial cell division inhibitor MipZ. Mol. Cell 2012, 46, 245–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tettelin, H.; Nelson, K.E.; Paulsen, I.T.; Eisen, J.A.; Read, T.D.; Peterson, S.; Heidelberg, J.; DeBoy, R.T.; Haft, D.H.; Dodson, R.J.; et al. Complete genome sequence of a virulent isolate of Streptococcus pneumoniae. Science 2001, 293, 498–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.M.; Smith, C.A.; Shingler, V.; Cross, M.A.; Hussain, A.A.; Pinkney, M. Regulation of replication and maintenance functions of broad host-range plasmid RK2. Basic Life Sci. 1985, 30, 261–276. [Google Scholar]
- Jagura-Burdzy, G.; Kostelidou, K.; Pole, J.; Khare, D.; Jones, A.; Williams, D.R.; Thomas, C.M. IncC of broad-host-range plasmid RK2 modulates KorB transcriptional repressor activity in vivo and operator binding in vitro. J. Bacteriol. 1999, 181, 2807–2815. [Google Scholar] [CrossRef] [Green Version]
- Kulinska, A.; Godziszewska, J.; Wojciechowska, A.; Ludwiczak, M.; Jagura-Burdzy, G. Global transcriptional regulation of backbone genes in broad-host-range plasmid RA3 from the IncU group involves segregation protein KorB (ParB family). Appl. Environ. Microbiol. 2016, 82, 2320–2335. [Google Scholar] [CrossRef] [Green Version]
- Bartosik, A.A.; Glabski, K.; Jecz, P.; Mikulska, S.; Fogtman, A.; Koblowska, M.; Jagura-Burdzy, G. Transcriptional profiling of ParA and ParB mutants in actively dividing cells of an opportunistic human pathogen Pseudomonas aeruginosa. PLoS ONE 2014, 9, e87276. [Google Scholar] [CrossRef]
- Lasocki, K.; Bartosik, A.A.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. Deletion of the parA (soj) homologue in Pseudomonas aeruginosa causes ParB instability and affects growth rate, chromosome segregation, and motility. J. Bacteriol. 2007, 189, 5762–5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawalek, A.; Glabski, K.; Bartosik, A.A.; Fogtman, A.; Jagura-Burdzy, G. Increased ParB level affects expression of stress response, adaptation and virulence operons and potentiates repression of promoters adjacent to the high affinity binding sites parS3 and parS4 in Pseudomonas aeruginosa. PLoS ONE 2017, 12, e0181726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, S.; Wright, A. Entropy as the driver of chromosome segregation. Nat. Rev. Microbiol. 2010, 8, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.; Mulder, B. Entropy-driven spatial organization of highly confined polymers: Lessons for the bacterial chromosome. Proc. Natl. Acad. Sci. USA 2006, 103, 12388–12393. [Google Scholar] [CrossRef] [Green Version]
- Weber, P.M.; Moessel, F.; Paredes, G.F.; Viehboeck, T.; Vischer, N.O.E.; Bulgheresi, S. A Bidimensional segregation mode maintains symbiont chromosome orientation toward its host. Curr. Biol. 2019, 29, 3018–3028. [Google Scholar] [CrossRef] [Green Version]
- Jakimowicz, D.; Mouz, S.; Zakrzewska-Czerwinska, J.; Chater, K.F. Developmental control of a parAB promoter leads to formation of sporulation-associated ParB complexes in Streptomyces coelicolor. J. Bacteriol. 2006, 188, 1710–1720. [Google Scholar] [CrossRef] [Green Version]
- Casart, Y.; Gamero, E.; Rivera-Gutierrez, S.; González-y-Merchand, J.A.; Salazar, L. par genes in Mycobacterium bovis and Mycobacterium smegmatis are arranged in an operon transcribed from “SigGC” promoters. BMC Microbiol. 2008, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Manuse, S.; Fleurie, A.; Zucchini, L.; Lesterlin, C.; Grangeasse, C. Role of eukaryotic-like serine/threonine kinases in bacterial cell division and morphogenesis. FEMS Microbiol. Rev. 2016, 40, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Vinardell, J.-M.; Lipa, P.; Karaś, M. Hanks-type serine/threonine protein kinases and phosphatases in bacteria: Roles in signaling and adaptation to various environments. Int. J. Mol. Sci. 2018, 19, e2872. [Google Scholar] [CrossRef] [Green Version]
- Carabetta, V.J.; Cristea, I.M. Regulation, function, and detection of protein acetylation in bacteria. J. Bacteriol. 2017, 199, e00107–e00117. [Google Scholar] [CrossRef] [Green Version]
- Baronian, G.; Ginda, K.; Berry, L.; Cohen-Gonsaud, M.; Zakrzewska-Czerwińska, J.; Jakimowicz, D.; Molle, V. Phosphorylation of Mycobacterium tuberculosis ParB participates in regulating the ParABS chromosome segregation system. PLoS ONE 2015, 10, e0119907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, Y.; Upadhyay, S.; Khan, S.; Nagarajan, S.N.; Forti, F.; Nandicoori, V.K. Protein kinase B (PknB) of Mycobacterium tuberculosis is essential for growth of the pathogen in vitro as well as for survival within the host. J. Biol. Chem. 2014, 289, 13858–13875. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | parS Sites Total/in ori Domain/Consensus | par Genes | Anucleate Cells in parB Mutant | Other Phenotypes associated with Mutation in parB Gene |
|---|---|---|---|---|
| Gram-positive: | ||||
| Firmicutes | ||||
| Bacillus subtilis | 10/8/TGTTNCACGTGAAACA [68] | non-essential | 1%–2% (wt 0.02%) [69] | Defect in sporulation, elongated cells, increased amount of DNA per cell, overreplication: >2 foci corresponding to ori in 35.4% cells (wt 15.3%), disturbed replication control and ori separation [70], impaired SMC loading (lack of foci) [6] |
| Streptococcus pneumoniae | 4/4/AGTTTCACGTGAAACT [71] | non-essential, no parA | 0.8% at 30 °C to 3.5% at 37 °C (wt 0%) [71] | No apparent growth defects, mild perturbations in chromosome segregation, decreased SMC loading near origin [71] |
| Actinobacteria | ||||
| Corynebacterium glutamicum | 10/10/TGTTNCACGTGAAACA [72] | non-essential | 43.8% in MMI medium, 11.6% in LB (wt 0%) [73] | Reduced growth rate in MMI medium, growth not affected in an LB medium, altered cell morphology (almost “coccoid” cells and elongated anucleate cells) [73], impaired SMC loading (lack of visible foci) [72] |
| Mycobacterium smegmatis | 3/3/GTTTCACGTGAAAC [74] | non-essential | 10.3% (wt 0.8%) [74] | Elongated cells, overreplication, disturbed septa formation, origin positioning, and chromosomal topology [75] |
| Streptomyces coelicolor | 21/21/tGTTTCACCTGAAACa [76] | non-essential | 13%–17.4% anucleate spores (wt 1%–2%) [77,78] | Disturbed sporulation, reduced growth rate, elongated cells, premature and irregular Z-ring formation [79] |
| Gram-negative: | ||||
| Alphaproteobacteria | ||||
| Caulobacter crescentus | 5/5/t/cGTTt/cCACGTGAAAca [80,81] | essential | Indispensable, severe chromosome segregation defects, ParB depletion results in defective Z-ring formation and cell division, formation of long polyploid cells [82] | |
| Hyphomonas neptunium | 2/2/TGTTTCACGTGAAACA [83] | essential | anucleate buds [83] | parB mutants could not be obtained, depletion of ParA blocks cell division [83] |
| Betaproteobacteria | ||||
| Burkholderia cenocepacia | chrI: 2/2/tGTTNCACGTGAAACa chrII: 6/6/gTTTATGCGCATAAAc [84,85,86,87] | non-essential | 1–14% (depending on mutated system) [85] | Reduced growth rate, reduction in cell size, compromised viability, defects in ori positioning [85] |
| Deltaproteobacteria | ||||
| Myxococcus xanthus | 22/22/TGTTCCACGTGGAACG [88] | essential | ParB depletion: 1% after 24 h, 10.1–21.6% after 36–48 h [88] | ParB depletion: aberrant cell morphology, anomalies in DNA segregation and cell death [88] |
| Gammaproteobacteria | ||||
| Pseudomonas aeruginosa | 9/4/TGTTCCACGtGGAACa half-parSs GTTCCAC or GTTTCAC [89,90,91] | non-essential | 2–4% in LB medium, to 7% in an M9 medium (wt < 0.01%) [92] | Reduced growth rate, 10–15% increase in cell size and 10% longer generation time, altered colony morphology, affected motility; decreased ParA stability [92] |
| Pseudomonas putida | ?*/3 TGTTCCACGTGGAACA [63] | non-essential | 5–10% in minimal medium during the transition from exponential to stationary phase [93] | Defects in chromosome partitioning, abnormal cell morphologies during the deceleration phase of growth independent of the medium used [63,93] |
| Vibrio cholerae | chrI: 3/3/NGTTNCACGTGAAACN chrII: 10/9/NTTTACANTGTAAAN [94] | non-essential chr1 essential chr2 | no change in parB1 mutant [95] | Increased frequency of replication initiation, disturbed ori positioning in cell poles [95], no segregation defect for V. cholerae chrI [94] |
| Deinococci | ||||
| Deinococcus radiodurans | chrI: 3/1/NGTTTcgcGtgaAACN [96] | non-essential | 8%—13% for ∆parB1, (wt >1%) [96] | Reduced growth rate for ∆parB1 [96] |
| Thermus thermophilus | 1/1/TGTTTCCCGTGAAACA [97] | non-essential | 3% for ∆parAB (wt 1.2%) [97] | No apparent growth defects for ∆parAB [97,98] |
| Species | ParBs (or ParB–parS complexes) Protein Partners | |||
|---|---|---|---|---|
| ParA dependent re-locations of ParB–parS complexes | Chromosome ori domain modelling | Localization/anchoring Cell division control | Replication initiation regulation | |
| Bacillus subtilis | + [70] | SMC; Interplay between SMC and ParB–parS complexes shown by ChIP-seq, FM, and mutational analysis (deletion of parB or parS sites affect SMC foci formation) [5,6] | DivIVA; Direct interaction confirmed by Co-IP, Spo0J may participate in switching between vegetative growth and sporulation [164] | #DnaA; Spo0J induces NTPase activity of Soj (ParA), the DnaA regulator [160,161] |
| Myxococcus xanthus | ND | #PadC; PadC mediates the binding of ParA to BacNOP cytoskeletal proteins (BLI); ParB may interact with BacNOP through a yet unidentified protein (pull-down) [141,165] | ||
| Mycobacterium smegmatis | + [166] | SMC; Interplay between SMC and ParB–parS complexes shown by FM and mutational analysis (deletion of parB affects SMC foci formation) [75] | #DivIVA (Wag31); ParA interacts with DivIVA and mediates nucleoid anchoring at the cell poles, co-localisation confirmed by the FM of labelled proteins, interactions proven by BACTH and pull-down [166,167] | |
| Streptococcus pneumoniae | lack of ParA; CpsD, Walker-type ATPase, is involved in ParB–parS movements [168] | SMC; Interplay between SMC and ParB–parS complexes shown by FM and mutational analysis (deletion of parB affects SMC foci formation) [71] | DivIVA; Direct interactions confirmed by Co-IP and BACTH [169] RocS; Co-localisation confirmed by FM of labelled proteins, direct interactions confirmed by Co-IP and MST [170], RocS and ParB participate in DNA segregation; CpsD*; Co-localisation confirmed by FM of fusion proteins, direct interaction (CpsD phosphorylation dependent) confirmed by Co-IP and MST, CpsD and ParB cooperate in coordination with DNA segregation, cell division, and capsule formation [168] | |
| Streptomyces coelicolor | + [171] | TopA; Interactions with ParB–parS complexes proven by pull-down, Co-IP, ChIP-seq; these interactions may support chromosome resolution [158] | #ParJ; ParJ negatively regulates the ParA polymerization indispensable for chromosome segregation, direct interaction confirmed by BACTH and SPR [172] #Scy; The polarity determinant interacts with ParA, co-localisation confirmed by FM of labelled proteins, direct interactions confirmed by BACTH, co-purification and SPR, ParA and Scy coordinate growth and chromosome segregation [173] | |
| Caulobacter crescentus | + [174] | SMC; Interplay with ParB–parS complexes shown by Hi-C, ChIP-seq, and mutational analysis, ParB dependent loading also detected on differently positioned parS sites [151] | PopZ*; Co-localisation confirmed by the FM of labelled proteins, direct interactions confirmed by Co-IP and SPR, EMSA (PopZ with ParB–parS complexes), PopZ–ParB–parS complexes mediate chromosome anchoring to the cell poles [175] #PopZ*; PopZ interacts also with ParA, co-localisation confirmed by FM of labelled proteins, direct interaction confirmed by Co-IP and SPR, interactions between PopZ and ParA mediate chromosome movement towards the swarmer pole [176] MipZ*; Co-localisation confirmed by the FM of labelled proteins, direct interactions confirmed by SPR and EMSA (MipZ with ParB–parS complexes), ParB–MipZ complexes control Z-ring positioning [177] #TipN; The polarity determinant interacts with ParA, direct interactions proven by FM and SPR, ParA–TipN complexes anchor ParB–parS (ori domain) to the new cell pole [103,178] | |
| Corynebacterium glutamicum | + [73] | SMC; Interplay with ParB–parS complexes shown by FM, ChIP-seq, deletion of parB or parSs results in the loss of SMC loading and foci formation [72] | FtsZ; Direct interactions confirmed by BACTH [73] PldP*; Direct interactions confirmed by BACTH [73], putative new cell division control system | |
| Deinococcus radiodurans | + [179] | MinC; Interactions with ParB1 (chrI), ParB3 (megaplasmid), and ParB4 (plasmid) confirmed by BACTH [180]; DivIVA; Interactions with ParB1, ParB2, ParB3, and ParB4 confirmed by BACTH and Co-IP [181]; ParB1 interacting with DivIVA may mediate chromosome anchoring to the cell poles and through interactions with FtsZ inhibitor MinC may affect the cell division. Interactions of DivIVA and MinC with ParB2-4 coordinate the segregation of multipartite genome and cell division. | DnaA, DnaB; interactions with ParB1(chrI) ParB2 (chrII) and ParB3 (megaplasmid), confirmed by BACTH and Co-IP; these interactions probably regulate DNA replication initiation [163] | |
| Vibrio cholerae | + [107] | #HubP; ParA1 interacts with HubP to mediate nucleoid anchoring at the cell poles; interactions confirmed by the FM of fusion proteins and BACTH [182] | DnaA; Interactions with ParB1 confirmed by BACTH, a proposed role in the regulation of replication initiation [95] | |
| Hyphomonas neptunium | + [83] | PopZ*; Co-localisation with ParB–parS in the swarmer cells and in the bud compartments confirmed by the FM of tagged proteins [83] | ||
| Rhodobacter sphaeroides | ND | MipZ*; Co-localization demonstrated by the FM of fusion proteins; direct interactions confirmed by BACTH; the ParB–MipZ system supports the Z-ring assembly and its stability [183] | ||
| Magnetospirillum gryphiswaldense | ND | MipZ1*; Co-localization demonstrated by the FM of fusion proteins; direct interactions confirmed by BLI, ParB–MipZ1 system controls Z-ring positioning; MipZ2* may also interact with ParB [184] | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawalek, A.; Wawrzyniak, P.; Bartosik, A.A.; Jagura-Burdzy, G. Rules and Exceptions: The Role of Chromosomal ParB in DNA Segregation and Other Cellular Processes. Microorganisms 2020, 8, 105. https://doi.org/10.3390/microorganisms8010105
Kawalek A, Wawrzyniak P, Bartosik AA, Jagura-Burdzy G. Rules and Exceptions: The Role of Chromosomal ParB in DNA Segregation and Other Cellular Processes. Microorganisms. 2020; 8(1):105. https://doi.org/10.3390/microorganisms8010105
Chicago/Turabian StyleKawalek, Adam, Pawel Wawrzyniak, Aneta Agnieszka Bartosik, and Grazyna Jagura-Burdzy. 2020. "Rules and Exceptions: The Role of Chromosomal ParB in DNA Segregation and Other Cellular Processes" Microorganisms 8, no. 1: 105. https://doi.org/10.3390/microorganisms8010105