Cellulases from Thermophiles Found by Metagenomics


 ,
, 
 and
and 
Abstract
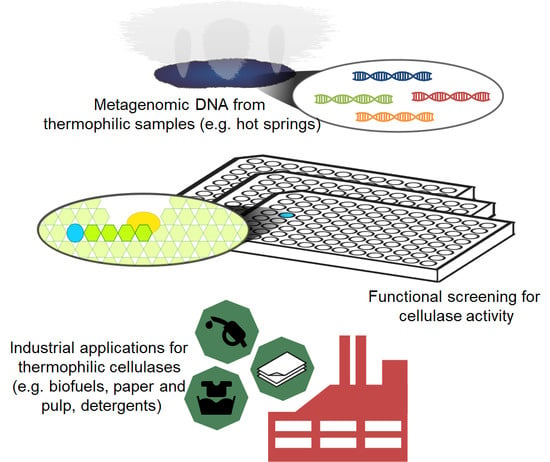
:
1. Introduction
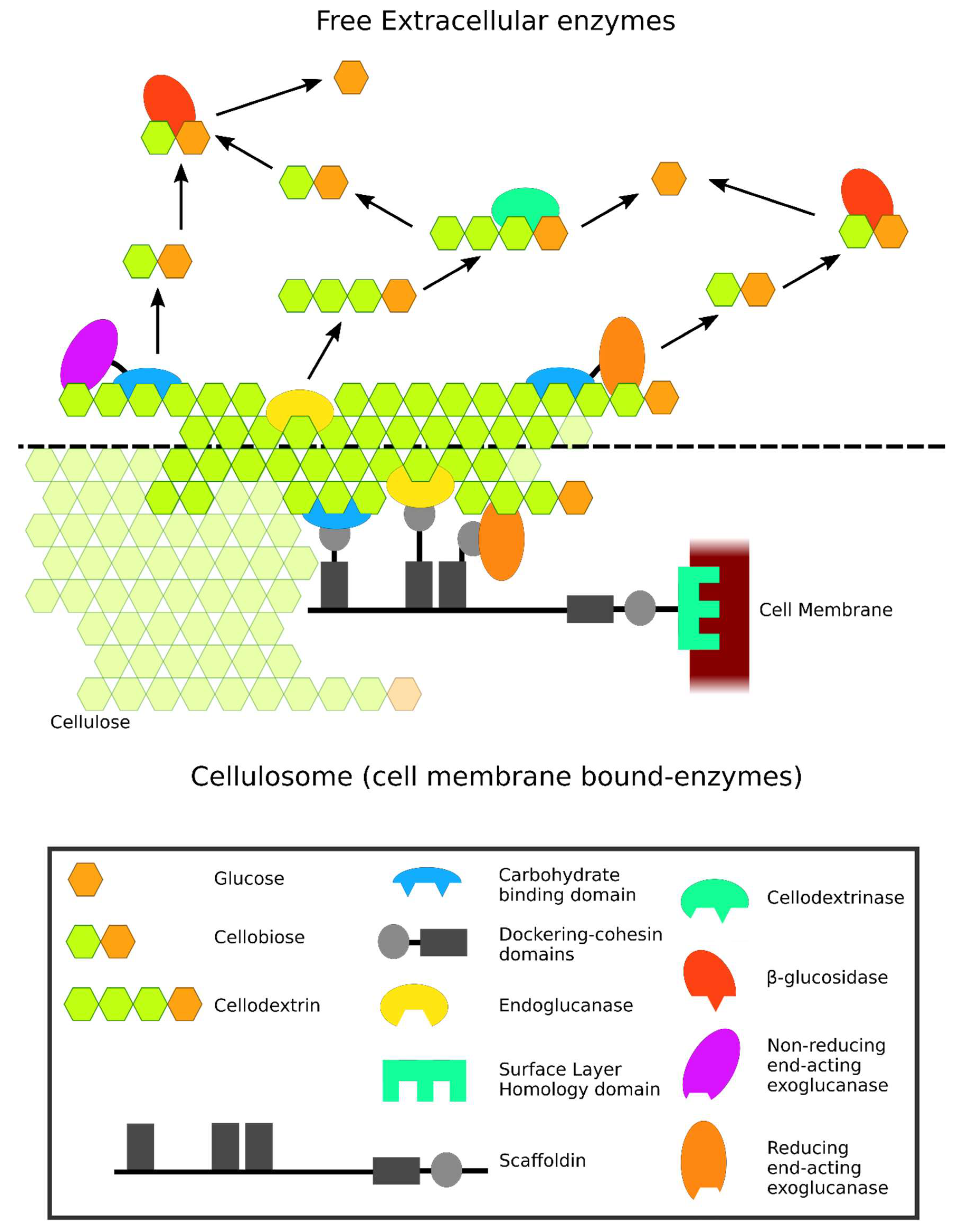
2. Modular Structure of Cellulases and their Classification
3. Factors Influencing Thermostability of Thermophile Cellulases
4. Biotechnological Applications by Thermophile Cellulases
4.1. Endoglucanase-Specific Industrial Applications
4.2. Exoglucanase-Specific Industrial Applications
4.3. β-glucosidase-Specific Industrial Applications
5. Metagenomics for the Search of Novel Cellulases
6. Thermophile Cellulases Characterized
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sandgren, M.; Ståhlberg, J.; Mitchinson, C. Structural and biochemical studies of GH family 12 cellulases: Improved thermal stability, and ligand complexes. Prog. Biophys. Mol. Biol. 2005, 89, 246–291. [Google Scholar] [CrossRef] [PubMed]
- Blumer-Schuette, S.E.; Kataeva, I.; Westpheling, J.; Adams, M.W.; Kelly, R.M. Extremely thermophilic microorganisms for biomass conversion: Status and prospects. Curr. Opin. Biotechnol. 2008, 19, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.-J.; Feng, J.-X. Mining metagenomes for novel cellulase genes. Biotechnol. Lett. 2010, 32, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Tewari, R.; Rana, S.S.; Soni, R.; Soni, S.K. Cellulases: Classification, Methods of Determination and Industrial Applications. Appl. Biochem. Biotechnol. 2016, 179, 1346–1380. [Google Scholar] [CrossRef] [PubMed]
- Couturier, M.; Feliu, J.; Haon, M.; Navarro, D.; Lesage-Meessen, L.; Coutinho, P.M.; Berrin, J.-G. A thermostable GH45 endoglucanase from yeast: Impact of its atypical multimodularity on activity. Microb. Cell Fact. 2011, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- López-Mondéjar, R.; Zühlke, D.; Becher, D.; Riedel, K.; Baldrian, P. Cellulose and hemicellulose decomposition by forest soil bacteria proceeds by the action of structurally variable enzymatic systems. Sci. Rep. 2016, 6, 25279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, B.; Chadha, B.S. Approaches for Bioprospecting Cellulases. In Extremophilic Enzymatic Processing of Lignocellulosic Feedstocks to Bioenergy; Sani, R.K., Krishnaraj, R.N., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 53–71. ISBN 978-3-319-54683-4. [Google Scholar]
- Bok, J.D.; Yernool, D.A.; Eveleigh, D.E. Purification, characterization, and molecular analysis of thermostable cellulases CelA and CelB from Thermotoga neapolitana. Appl. Environ. Microbiol. 1998, 64, 4774–4781. [Google Scholar] [PubMed]
- Elleuche, S.; Schäfers, C.; Blank, S.; Schröder, C.; Antranikian, G. Exploration of extremophiles for high temperature biotechnological processes. Curr. Opin. Microbiol. 2015, 25, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.; Nain, L.; Labrou, N.E.; Shukla, P. Bioprospecting of functional cellulases from metagenome for second generation biofuel production: A review. Crit. Rev. Microbiol. 2018, 44, 244–257. [Google Scholar] [CrossRef] [PubMed]
- DeCastro, M.-E.; Rodríguez-Belmonte, E.; González-Siso, M.-I. Metagenomics of Thermophiles with a Focus on Discovery of Novel Thermozymes. Front. Microbiol. 2016, 7, 1521. [Google Scholar] [CrossRef] [PubMed]
- Wagner, I.D.; Wiegel, J. Diversity of thermophilic anaerobes. Ann. N. Y. Acad. Sci. 2008, 1125, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, J.; Liu, Q.; Zhang, C.; Ma, Q. Molecular cloning of novel cellulase genes cel9A and cel12A from Bacillus licheniformis GXN151 and synergism of their encoded polypeptides. Curr. Microbiol. 2004, 49, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Crennell, S.J.; Hreggvidsson, G.O.; Nordberg Karlsson, E. The structure of Rhodothermus marinus Cel12A, a highly thermostable family 12 endoglucanase, at 1.8 A resolution. J. Mol. Biol. 2002, 320, 883–897. [Google Scholar] [CrossRef]
- Chhabra, S.R.; Kelly, R.M. Biochemical characterization of Thermotoga maritima endoglucanase Cel74 with and without a carbohydrate binding module (CBM). FEBS Lett. 2002, 531, 375–380. [Google Scholar] [CrossRef]
- Peer, A.; Smith, S.P.; Bayer, E.A.; Lamed, R.; Borovok, I. Noncellulosomal cohesin- and dockerin-like modules in the three domains of life. FEMS Microbiol. Lett. 2009, 291, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, E.A.; Belaich, J.-P.; Shoham, Y.; Lamed, R. The cellulosomes: Multienzyme machines for degradation of plant cell wall polysaccharides. Annu. Rev. Microbiol. 2004, 58, 521–554. [Google Scholar] [CrossRef] [PubMed]
- Sathya, T.A.; Khan, M. Diversity of glycosyl hydrolase enzymes from metagenome and their application in food industry. J. Food Sci. 2014, 79, R2149–R2156. [Google Scholar] [CrossRef] [PubMed]
- Poidevin, L.; Feliu, J.; Doan, A.; Berrin, J.-G.; Bey, M.; Coutinho, P.M.; Henrissat, B.; Record, E.; Heiss-Blanquet, S. Insights into exo- and endoglucanase activities of family 6 glycoside hydrolases from Podospora anserina. Appl. Environ. Microbiol. 2013, 79, 4220–4229. [Google Scholar] [CrossRef] [PubMed]
- Henrissat, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1991, 280, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Lee, M.J.; Cho, H.-Y.; Lee, J.S.; Lee, M.-H.; Chung, C.W.; Shin, D.-H.; Rhee, Y.H.; Son, K.-H.; Park, H.-Y. Genetic and functional characterization of an extracellular modular GH6 endo-β-1,4-glucanase from an earthworm symbiont, Cellulosimicrobium funkei HY-13. Antonie Van Leeuwenhoek 2016, 109, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Halldórsdóttir, S.; Thórólfsdóttir, E.T.; Spilliaert, R.; Johansson, M.; Thorbjarnardóttir, S.H.; Palsdottir, A.; Hreggvidsson, G.O.; Kristjánsson, J.K.; Holst, O.; Eggertsson, G. Cloning, sequencing and overexpression of a Rhodothermus marinus gene encoding a thermostable cellulase of glycosyl hydrolase family 12. Appl. Microbiol. Biotechnol. 1998, 49, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Ausili, A.; Cobucci-Ponzano, B.; Di Lauro, B.; D’Avino, R.; Perugino, G.; Bertoli, E.; Scirè, A.; Rossi, M.; Tanfani, F.; Moracci, M. A comparative infrared spectroscopic study of glycoside hydrolases from extremophilic archaea revealed different molecular mechanisms of adaptation to high temperatures. Proteins 2007, 67, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.F.; Sanderson, I.; Moracci, M.; Ciaramella, M.; Nucci, R.; Rossi, M.; Pearl, L.H. Crystal structure of the β-glycosidase from the hyperthermophilic archeon Sulfolobus solfataricus: Resilience as a key factor in thermostability. J. Mol. Biol. 1997, 271, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Wu, I.; Arnold, F.H. Engineered thermostable fungal Cel6A and Cel7A cellobiohydrolases hydrolyze cellulose efficiently at elevated temperatures. Biotechnol. Bioeng. 2013, 110, 1874–1883. [Google Scholar] [CrossRef] [PubMed]
- Rawat, R.; Kumar, S.; Chadha, B.S.; Kumar, D.; Oberoi, H.S. An acidothermophilic functionally active novel GH12 family endoglucanase from Aspergillus niger HO: Purification, characterization and molecular interaction studies. Antonie Van Leeuwenhoek 2015, 107, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Voutilainen, S.P.; Puranen, T.; Siika-Aho, M.; Lappalainen, A.; Alapuranen, M.; Kallio, J.; Hooman, S.; Viikari, L.; Vehmaanperä, J.; Koivula, A. Cloning, expression, and characterization of novel thermostable family 7 cellobiohydrolases. Biotechnol. Bioeng. 2008, 101, 515–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voutilainen, S.P.; Murray, P.G.; Tuohy, M.G.; Koivula, A. Expression of Talaromyces emersonii cellobiohydrolase Cel7A in Saccharomyces cerevisiae and rational mutagenesis to improve its thermostability and activity. Protein Eng. Des. Sel. 2010, 23, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-L.; Li, H.; Li, A.-N.; Li, D.-C. Cloning of a gene encoding thermostable cellobiohydrolase from the thermophilic fungus Chaetomium thermophilum and its expression in Pichia pastoris. J. Appl. Microbiol. 2009, 106, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Karnaouri, A.C.; Topakas, E.; Christakopoulos, P. Cloning, expression, and characterization of a thermostable GH7 endoglucanase from Myceliophthora thermophila capable of high-consistency enzymatic liquefaction. Appl. Microbiol. Biotechnol. 2014, 98, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Hua, C.; Yang, S.; Li, Y.; Jiang, Z. High level expression of extracellular secretion of a β-glucosidase gene (PtBglu3) from Paecilomyces thermophila in Pichia pastoris. Protein Expr. Purif. 2012, 84, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dun, B.; Shi, P.; Ma, R.; Luo, H.; Bai, Y.; Xie, X.; Yao, B. A Novel GH7 Endo-β-1,4-Glucanase from Neosartorya fischeri P1 with Good Thermostability, Broad Substrate Specificity and Potential Application in the Brewing Industry. PLoS ONE 2015, 10, e0137485. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, M.J.; D’haeseleer, P.; Hazen, T.C.; Simmons, B.A.; Adams, P.D.; Hadi, M.Z. Glycoside hydrolases from a targeted compost metagenome, activity-screening and functional characterization. BMC Biotechnol. 2012, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, G.; Li, Y.; Yang, L.; Liang, Y.; Jin, H.; Han, W.; Feng, Y.; Zhang, Z. Cloning, Expression, and Characterization of a Thermophilic Endoglucanase, AcCel12B from Acidothermus cellulolyticus 11B. Int. J. Mol. Sci. 2015, 16, 25080–25095. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Bao, L.; Chang, L.; Liu, Z.; You, C.; Lu, H. Beta-xylosidase activity of a GH3 glucosidase/xylosidase from yak rumen metagenome promotes the enzymatic degradation of hemicellulosic xylans. Lett. Appl. Microbiol. 2012, 54, 79–87. [Google Scholar] [CrossRef] [PubMed]
- McAndrew, R.P.; Park, J.I.; Heins, R.A.; Reindl, W.; Friedland, G.D.; D’haeseleer, P.; Northen, T.; Sale, K.L.; Simmons, B.A.; Adams, P.D. From soil to structure, a novel dimeric β-glucosidase belonging to glycoside hydrolase family 3 isolated from compost using metagenomic analysis. J. Biol. Chem. 2013, 288, 14985–14992. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-M.; Lee, Y.-S.; Seo, S.-H.; Yoon, S.-H.; Kim, S.-J.; Hahn, B.-S.; Sim, J.-S.; Koo, B.-S. Screening and Characterization of a Novel Cellulase Gene from the Gut Microflora of Hermetia illucens Using Metagenomic Library. J. Microbiol. Biotechnol. 2014, 24, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Girfoglio, M.; Rossi, M.; Cannio, R. Cellulose degradation by Sulfolobus solfataricus requires a cell-anchored endo-β-1-4-glucanase. J. Bacteriol. 2012, 194, 5091–5100. [Google Scholar] [CrossRef] [PubMed]
- Biver, S.; Stroobants, A.; Portetelle, D.; Vandenbol, M. Two promising alkaline β-glucosidases isolated by functional metagenomics from agricultural soil, including one showing high tolerance towards harsh detergents, oxidants and glucose. J. Ind. Microbiol. Biotechnol. 2014, 41, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Konishi, T.; Takeda, T. Biochemical characterization of Magnaporthe oryzae β-glucosidases for efficient β-glucan hydrolysis. Appl. Microbiol. Biotechnol. 2011, 91, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Bansal, N.; Kumar, S.; Bischoff, K.M.; Sani, R.K. Improved lignocellulose conversion to biofuels with thermophilic bacteria and thermostable enzymes. Bioresour. Technol. 2013, 128, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Harnpicharnchai, P.; Champreda, V.; Sornlake, W.; Eurwilaichitr, L. A thermotolerant beta-glucosidase isolated from an endophytic fungi, Periconia sp., with a possible use for biomass conversion to sugars. Protein Expr. Purif. 2009, 67, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Fusco, F.A.; Fiorentino, G.; Pedone, E.; Contursi, P.; Bartolucci, S.; Limauro, D. Biochemical characterization of a novel thermostable β-glucosidase from Dictyoglomus turgidum. Int. J. Biol. Macromol. 2018, 113, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Boyce, A.; Walsh, G. Characterisation of a novel thermostable endoglucanase from Alicyclobacillus vulcanalis of potential application in bioethanol production. Appl. Microbiol. Biotechnol. 2015, 99, 7515–7525. [Google Scholar] [CrossRef] [PubMed]
- Ando, S.; Ishida, H.; Kosugi, Y.; Ishikawa, K. Hyperthermostable Endoglucanase from Pyrococcus horikoshii. Appl. Environ. Microbiol. 2002, 68, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wang, J.; Zhang, Z.; Shi, P.; Luo, H.; Huang, H.; Feng, Y.; Yao, B. Extremely acidic beta-1,4-glucanase, CelA4, from thermoacidophilic Alicyclobacillus sp. A4 with high protease resistance and potential as a pig feed additive. J. Agric. Food Chem. 2010, 58, 1970–1975. [Google Scholar] [CrossRef] [PubMed]
- Vuong, T.V.; Wilson, D.B. Processivity, synergism, and substrate specificity of Thermobifida fusca Cel6B. Appl. Environ. Microbiol. 2009, 75, 6655–6661. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Liu, S.; Jing, H.; Sun, R.; Liu, M.; Chen, H.; Wu, Q.; Han, X. Cloning and expression of A. oryzae β-glucosidase in Pichia pastoris. Mol. Biol. Rep. 2014, 41, 7567–7573. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yan, Q.; Yang, Y.; Yang, S.; Liu, Y.; Jiang, Z. Expression and characterization of a novel β-glucosidase, with transglycosylation and exo-β-1,3-glucanase activities, from Rhizomucor miehei. Food Chem. 2015, 175, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A.; Ramón, D.; Vallés, S.; Lluch, M.A.; MacCabe, A.P. Heterologous Expression in Aspergillus nidulans of a Trichoderma longibrachiatum Endoglucanase of Enological Relevance. J. Agric. Food Chem. 2000, 48, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Lee, C.-M.; Kim, M.-Y.; Yeo, Y.-S.; Yoon, S.-H.; Kang, H.-C.; Koo, B.-S. Screening and characterization of an enzyme with beta-glucosidase activity from environmental DNA. J. Microbiol. Biotechnol. 2007, 17, 905–912. [Google Scholar] [PubMed]
- Lee, G.-W.; Yoo, M.-H.; Shin, K.-C.; Kim, K.-R.; Kim, Y.-S.; Lee, K.-W.; Oh, D.-K. β-glucosidase from Penicillium aculeatum hydrolyzes exo-, 3-O-, and 6-O-β-glucosides but not 20-O-β-glucoside and other glycosides of ginsenosides. Appl. Microbiol. Biotechnol. 2013, 97, 6315–6324. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Yeom, S.-J.; Oh, D.-K. Characterization of a GH3 family β-glucosidase from Dictyoglomus turgidum and its application to the hydrolysis of isoflavone glycosides in spent coffee grounds. J. Agric. Food Chem. 2011, 59, 11812–11818. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Ryan, B.; Henehan, G.T.M. β-Glucosidase from Streptomyces griseus: Nanoparticle immobilisation and application to alkyl glucoside synthesis. Protein Expr. Purif. 2017, 132, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Placido, A.; Hai, T.; Ferrer, M.; Chernikova, T.N.; Distaso, M.; Armstrong, D.; Yakunin, A.F.; Toshchakov, S.V.; Yakimov, M.M.; Kublanov, I.V.; et al. Diversity of hydrolases from hydrothermal vent sediments of the Levante Bay, Vulcano Island (Aeolian archipelago) identified by activity-based metagenomics and biochemical characterization of new esterases and an arabinopyranosidase. Appl. Microbiol. Biotechnol. 2015, 99, 10031–10046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leis, B.; Heinze, S.; Angelov, A.; Pham, V.T.T.; Thürmer, A.; Jebbar, M.; Golyshin, P.N.; Streit, W.R.; Daniel, R.; Liebl, W. Functional Screening of Hydrolytic Activities Reveals an Extremely Thermostable Cellulase from a Deep-Sea Archaeon. Front. Bioeng. Biotechnol. 2015, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Clark, M.E.; Nadler, D.C.; Huffer, S.; Chokhawala, H.A.; Rowland, S.E.; Blanch, H.W.; Clark, D.S.; Robb, F.T. Identification and characterization of a multidomain hyperthermophilic cellulase from an archaeal enrichment. Nat. Commun. 2011, 2, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröder, C.; Elleuche, S.; Blank, S.; Antranikian, G. Characterization of a heat-active archaeal β-glucosidase from a hydrothermal spring metagenome. Enzyme Microb. Technol. 2014, 57, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Khan, H.; Azam, S.S.; Telke, A.; Kim, S.W.; Chung, Y.R. Cloning and functional characterization of endo-β-1,4-glucanase gene from metagenomic library of vermicompost. J. Microbiol. 2013, 51, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.J.; Jeong, Y.S.; Kim, Y.H.; Kim, S.K.; Na, H.B.; Kim, J.; Yun, H.D.; Kim, H. Construction of a Metagenomic Library from Compost and Screening of Cellulase- and Xylanase-positive Clones. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 702–708. [Google Scholar] [CrossRef]
- Okano, H.; Ozaki, M.; Kanaya, E.; Kim, J.-J.; Angkawidjaja, C.; Koga, Y.; Kanaya, S. Structure and stability of metagenome-derived glycoside hydrolase family 12 cellulase (LC-CelA) a homolog of Cel12A from Rhodothermus marinus. FEBS Open Bio 2014, 4, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Geng, A.; Zhang, J.; Wei, Y.; Zhang, L.; Qian, C.; Wang, Q.; Wang, S.; Zhou, Z. Discovery of (hemi-)cellulase genes in a metagenomic library from a biogas digester using 454 pyrosequencing. Appl. Microbiol. Biotechnol. 2013, 97, 8173–8182. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, T.M.; Paiva, J.H.; Ruiz, D.M.; Cairo, J.P.L.F.; Pereira, I.O.; Paixão, D.A.A.; de Almeida, R.F.; Tonoli, C.C.C.; Ruller, R.; Santos, C.R.; et al. Structure and function of a novel cellulase 5 from sugarcane soil metagenome. PLoS ONE 2013, 8, e83635. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, W.-D.; Zhao, X.-L.; Shen, W.-J.; Cao, H.; Cui, Z.-L. Cloning and functional characterization of a novel endo-β-1,4-glucanase gene from a soil-derived metagenomic library. Appl. Microbiol. Biotechnol. 2011, 89, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Pottkämper, J.; Barthen, P.; Ilmberger, N.; Schwaneberg, U.; Schenk, A.; Schulte, M.; Ignatiev, N.; Streit, W.R. Applying metagenomics for the identification of bacterial cellulases that are stable in ionic liquids. Green Chem. 2009, 11, 957. [Google Scholar] [CrossRef]
- Feng, Y.; Duan, C.-J.; Pang, H.; Mo, X.-C.; Wu, C.-F.; Yu, Y.; Hu, Y.-L.; Wei, J.; Tang, J.-L.; Feng, J.-X. Cloning and identification of novel cellulase genes from uncultured microorganisms in rabbit cecum and characterization of the expressed cellulases. Appl. Microbiol. Biotechnol. 2007, 75, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, M.V.; Fernández-Arrojo, L.; Gil-Martínez, J.; Montesinos, A.; Chernikova, T.N.; Nechitaylo, T.Y.; Waliszek, A.; Tortajada, M.; Rojas, A.; Huws, S.A.; et al. Microbial β-glucosidases from cow rumen metagenome enhance the saccharification of lignocellulose in combination with commercial cellulase cocktail. Biotechnol. Biofuels 2012, 5, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Escudero, M.; Del Pozo, M.V.; Marín-Navarro, J.; González, B.; Golyshin, P.N.; Polaina, J.; Ferrer, M.; Sanz-Aparicio, J. Structural and Functional Characterization of a Ruminal β-Glycosidase Defines a Novel Subfamily of Glycoside Hydrolase Family 3 with Permuted Domain Topology. J. Biol. Chem. 2016, 291, 24200–24214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beloqui, A.; Nechitaylo, T.Y.; López-Cortés, N.; Ghazi, A.; Guazzaroni, M.-E.; Polaina, J.; Strittmatter, A.W.; Reva, O.; Waliczek, A.; Yakimov, M.M.; et al. Diversity of glycosyl hydrolases from cellulose-depleting communities enriched from casts of two earthworm species. Appl. Environ. Microbiol. 2010, 76, 5934–5946. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Qian, C.; Zhang, X.-Z.; Liu, N.; Liu, N.; Yan, X.; Zhou, Z. Characterization of a novel thermostable β-glucosidase from a metagenomic library of termite gut. Enzyme Microb. Technol. 2012, 51, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, N.; Qian, C.; Wang, Q.; Wang, Q.; Long, Y.; Huang, Y.; Zhou, Z.; Yan, X. Phylogenetic and functional analysis of gut microbiota of a fungus-growing higher termite: Bacteroidetes from higher termites are a rich source of β-glucosidase genes. Microb. Ecol. 2014, 68, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Himmel, M.E.; Adney, W.S.; Tucker, M.P.; Grohmann, K. Thermostable Purified Endoglucanas from Acidothermus cellulolyticus ATCC 43068. U.S. Patent 5,275,944, 4 January 1994. [Google Scholar]
- Nurachman, Z.; Kurniasih, S.D.; Puspitawati, F.; Hadi, S.; Radjasa, O.K.; Natalia, D. Cloning of the Endoglucanase Gene from a Bacillus amyloliquefaciens PSM 3.1 in Escherichia coli Revealed Catalytic Triad Residues Thr-His-Glu. Am. J. Biochem. Biotechnol. 2010, 6, 268–274. [Google Scholar] [CrossRef]
- Bischoff, K.M.; Rooney, A.P.; Li, X.-L.; Liu, S.; Hughes, S.R. Purification and characterization of a family 5 endoglucanase from a moderately thermophilic strain of Bacillus licheniformis. Biotechnol. Lett. 2006, 28, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-J.; Lee, Y.-S.; Park, I.-H.; Chandra, M.S.; Kim, K.-K.; Choi, Y.-L. Molecular cloning, purification and characterization of thermostable beta-1,3-1,4 glucanase from Bacillus subtilis A8-8. Indian J. Biochem. Biophys. 2010, 47, 203–210. [Google Scholar] [PubMed]
- Chhabra, S.R.; Shockley, K.R.; Ward, D.E.; Kelly, R.M. Regulation of endo-acting glycosyl hydrolases in the hyperthermophilic bacterium Thermotoga maritima grown on glucan- and mannan-based polysaccharides. Appl. Environ. Microbiol. 2002, 68, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Vasu, P.; Persson, S.; Mort, A.J.; Somerville, C.R. Development and application of a suite of polysaccharide-degrading enzymes for analyzing plant cell walls. Proc. Natl. Acad. Sci. USA 2006, 103, 11417–11422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calza, R.E.; Irwin, D.C.; Wilson, D.B. Purification and characterization of two β-1,4-endoglucanases from Thermomonospora fusca. Biochemistry 1985, 24, 7797–7804. [Google Scholar] [CrossRef]
- Yin, Y.-R.; Zhang, F.; Hu, Q.-W.; Xian, W.-D.; Hozzein, W.N.; Zhou, E.-M.; Ming, H.; Nie, G.-X.; Li, W.-J. Heterologous expression and characterization of a novel halotolerant, thermostable, and alkali-stable GH6 endoglucanase from Thermobifida halotolerans. Biotechnol. Lett. 2015, 37, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Cazemier, A.E.; Verdoes, J.C.; Op den Camp, H.J.M.; Hackstein, J.H.P.; van Ooyen, A.J. A beta-1,4-endoglucanase-encoding gene from Cellulomonas pachnodae. Appl. Microbiol. Biotechnol. 1999, 52, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, J.; Zhang, W.; Huang, H.; Shi, P.; Luo, H.; Liu, B.; Zhang, Y.; Zhang, Z.; Fan, Y.; et al. A Neutral Thermostable β-1,4-Glucanase from Humicola insolens Y1 with Potential for Applications in Various Industries. PLoS ONE 2015, 10, e0124925. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Chen, H.; Ljungdahl, L.G. Two cellulases, CelA and CelC, from the polycentric anaerobic fungus Orpinomyces strain PC-2 contain N-terminal docking domains for a cellulase-hemicellulase complex. Appl. Environ. Microbiol. 1997, 63, 4721–4728. [Google Scholar] [PubMed]
- Takashima, S.; Nakamura, A.; Hidaka, M.; Masaki, H.; Uozumi, T. Cloning, sequencing, and expression of the cellulase genes of Humicola grisea var. thermoidea. J. Biotechnol. 1996, 50, 137–147. [Google Scholar] [CrossRef]
- Wei, X.-M.; Qin, Y.; Qu, Y. Molecular Cloning and Characterization of Two Major Endoglucanases from Penicillium decumbens. J. Microbiol. Biotechnol. 2010, 20, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Miettinen-Oinonen, A.; Londesborough, J.; Joutsjoki, V.; Lantto, R.; Vehmaanperä, J. Three cellulases from Melanocarpus albomyces for textile treatment at neutral pH. Enzyme Microb. Technol. 2004, 34, 332–341. [Google Scholar] [CrossRef]
- Yoo, J.-S.; Jung, Y.-J.; Chung, S.-Y.; Lee, Y.-C.; Choi, Y.-L. Molecular cloning and characterization of CMCase gene (celC) from Salmonella typhimurium UR. J. Microbiol. 2004, 42, 205–210. [Google Scholar] [PubMed]
- Kim, J.O.; Park, S.R.; Lim, W.J.; Ryu, S.K.; Kim, M.K.; An, C.L.; Cho, S.J.; Park, Y.W.; Kim, J.H.; Yun, H.D. Cloning and characterization of thermostable endoglucanase (Cel8Y) from the hyperthermophilic Aquifex aeolicus VF5. Biochem. Biophys. Res. Commun. 2000, 279, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Hakamada, Y.; Endo, K.; Takizawa, S.; Kobayashi, T.; Shirai, T.; Yamane, T.; Ito, S. Enzymatic properties, crystallization, and deduced amino acid sequence of an alkaline endoglucanase from Bacillus circulans. Biochim. Biophys. Acta 2002, 1570, 174–180. [Google Scholar] [CrossRef]
- Ul Haq, I.; Akram, F.; Khan, M.A.; Hussain, Z.; Nawaz, A.; Iqbal, K.; Shah, A.J. CenC, a multidomain thermostable GH9 processive endoglucanase from Clostridium thermocellum: Cloning, characterization and saccharification studies. World J. Microbiol. Biotechnol. 2015, 31, 1699–1710. [Google Scholar] [CrossRef] [PubMed]
- Zverlov, V.; Mahr, S.; Riedel, K.; Bronnenmeier, K. Properties and gene structure of a bifunctional cellulolytic enzyme (CelA) from the extreme thermophile “Anaerocellum thermophilum” with separate glycosyl hydrolase family 9 and 48 catalytic domains. Microbiology 1998, 144, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Z.; Sathitsuksanoh, N.; Zhang, Y.-H.P. Glycoside hydrolase family 9 processive endoglucanase from Clostridium phytofermentans: Heterologous expression, characterization, and synergy with family 48 cellobiohydrolase. Bioresour. Technol. 2010, 101, 5534–5538. [Google Scholar] [CrossRef] [PubMed]
- Liebl, W.; Ruile, P.; Bronnenmeier, K.; Riedel, K.; Lottspeich, F.; Greif, I. Analysis of a Thermotoga maritima DNA fragment encoding two similar thermostable cellulases, CelA and CelB, and characterization of the recombinant enzymes. Microbiology 1996, 142, 2533–2542. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.W.; Driskill, L.E.; Callen, W.; Snead, M.A.; Mathur, E.J.; Kelly, R.M. An Endoglucanase, EglA, from the Hyperthermophilic Archaeon Pyrococcus Furiosus Hydrolyzes β-1,4 Bonds in Mixed-Linkage (1→3),(1→4)-β-D-Glucans and Cellulose. J. Bacteriol. 1999, 181, 284–290. [Google Scholar] [PubMed]
- Huang, Y.; Krauss, G.; Cottaz, S.; Driguez, H.; Lipps, G. A highly acid-stable and thermostable endo-beta-glucanase from the thermoacidophilic archaeon Sulfolobus solfataricus. Biochem. J. 2005, 385, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Irdani, T.; Perito, B.; Mastromei, G. Characterization of a Streptomyces rochei endoglucanase. Ann. N. Y. Acad. Sci. 1996, 782, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Siika-aho, M.; Tenkanen, M.; Tjerneld, F. Enzymatic properties of the low molecular mass endoglucanases Cel12A (EG III) and Cel45A (EG V) of Trichoderma reesei. J. Biotechnol. 2002, 99, 63–78. [Google Scholar] [CrossRef]
- Warner, C.D.; Hoy, J.A.; Shilling, T.C.; Linnen, M.J.; Ginder, N.D.; Ford, C.F.; Honzatko, R.B.; Reilly, P.J. Tertiary structure and characterization of a glycoside hydrolase family 44 endoglucanase from Clostridium acetobutylicum. Appl. Environ. Microbiol. 2010, 76, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.K.; Diderichsen, B.; Jørgensen, P.L. celA from Bacillus lautus PL236 encodes a novel cellulose-binding endo-beta-1,4-glucanase. J. Bacteriol. 1992, 174, 3522–3531. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, M.M.; Matsumoto, M.; Karita, S.; Kimura, T.; Sakka, K.; Ohmiya, K. Purification and Characterization of the Family J Catalytic Domain Derived from the Clostridium thermocellum Endoglucanase CelJ. Biosci. Biotechnol. Biochem. 1997, 61, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Koga, J.; Baba, Y.; Shimonaka, A.; Nishimura, T.; Hanamura, S.; Kono, T. Purification and characterization of a new family 45 endoglucanase, STCE1, from Staphylotrichum coccosporum and its overproduction in Humicola insolens. Appl. Environ. Microbiol. 2008, 74, 4210–4217. [Google Scholar] [CrossRef] [PubMed]
- Wonganu, B.; Pootanakit, K.; Boonyapakron, K.; Champreda, V.; Tanapongpipat, S.; Eurwilaichitr, L. Cloning, expression and characterization of a thermotolerant endoglucanase from Syncephalastrum racemosum (BCC18080) in Pichia pastoris. Protein Expr. Purif. 2008, 58, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.; Shimonaka, A.; Koga, J.; Murashima, K.; Kubota, H.; Kono, T. Purification and characterization of a new endo-1,4-beta-D-glucanase from Beltraniella portoricensis. Biosci. Biotechnol. Biochem. 2005, 69, 1198–1201. [Google Scholar] [CrossRef] [PubMed]
- Eckert, K.; Schneider, E. A thermoacidophilic endoglucanase (CelB) from Alicyclobacillus acidocaldarius displays high sequence similarity to arabinofuranosidases belonging to family 51 of glycoside hydrolases. Eur. J. Biochem. 2003, 270, 3593–3602. [Google Scholar] [CrossRef] [PubMed]
- Brás, J.L.A.; Cartmell, A.; Carvalho, A.L.M.; Verzé, G.; Bayer, E.A.; Vazana, Y.; Correia, M.A.S.; Prates, J.A.M.; Ratnaparkhe, S.; Boraston, A.B.; et al. Structural insights into a unique cellulase fold and mechanism of cellulose hydrolysis. Proc. Natl. Acad. Sci. USA 2011, 108, 5237–5242. [Google Scholar] [CrossRef] [PubMed]
- Park, C.-S.; Kawaguchi, T.; Sumitani, J.-I.; Takada, G.; Izumori, K.; Arai, M. Cloning and sequencing of an exoglucanase gene from Streptomyces sp. M 23, and its expression in Streptomyces lividans TK-24. J. Biosci. Bioeng. 2005, 99, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lao, G.; Wilson, D.B. Characterization of a Thermomonospora fusca exocellulase. Biochemistry 1995, 34, 3386–3395. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-A.; Li, D.-C.; Er, S.-J.; Zhang, Y. Cloning and expressing of cellulase gene (cbh2) from thermophilic fungi Chaetomium thermophilum CT2. Sheng Wu Gong Cheng Xue Bao 2005, 21, 892–899. [Google Scholar] [PubMed]
- Bukhtojarov, F.E.; Ustinov, B.B.; Salanovich, T.N.; Antonov, A.I.; Gusakov, A.V.; Okunev, O.N.; Sinitsyn, A.P. Cellulase complex of the fungus Chrysosporium lucknowense: Isolation and characterization of endoglucanases and cellobiohydrolases. Biochemistry. 2004, 69, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Toda, H.; Nagahata, N.; Amano, Y.; Nozaki, K.; Kanda, T.; Okazaki, M.; Shimosaka, M. Gene cloning of cellobiohydrolase II from the white rot fungus Irpex lacteus MC-2 and its expression in Pichia pastoris. Biosci. Biotechnol. Biochem. 2008, 72, 3142–3147. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, F.; Gao, F.; Wang, L.; Zhao, J.; Qu, Y. Purification and characterization of a novel cellobiohydrolase (PdCel6A) from Penicillium decumbens JU-A10 for bioethanol production. Bioresour. Technol. 2011, 102, 8339–8342. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, B.; Liu, Z.; Yang, Q. Cloning of two cellobiohydrolase genes from Trichoderma viride and heterogenous expression in yeast Saccharomyces cerevisiae. Mol. Biol. Rep. 2010, 37, 2135–2140. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.N.; Akimenko, V.K. Isolation of a Cellobiohydrolase of Clostridium thermocellum Capable of Degrading Natural Crystalline Substrates. Biochem. Biophys. Res. Commun. 1993, 192, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Kataeva, I.; Li, X.-L.; Chen, H.; Choi, S.-K.; Ljungdahl, L.G. Cloning and sequence analysis of a new cellulase gene encoding CelK, a major cellulosome component of Clostridium thermocellum: Evidence for gene duplication and recombination. J. Bacteriol. 1999, 181, 5288–5295. [Google Scholar] [PubMed]
- Zverlov, V.V.; Velikodvorskaya, G.A.; Schwarz, W.H. A newly described cellulosomal cellobiohydrolase, CelO, from Clostridium thermocellum: Investigation of the exo-mode of hydrolysis, and binding capacity to crystalline cellulose. Microbiology 2002, 148, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Takada, G.; Kawaguchi, T.; Sumitani, J.; Arai, M. Expression of Aspergillus aculeatus No. F-50 cellobiohydrolase I (cbhI) and beta-glucosidase 1 (bgl1) genes by Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 1998, 62, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Moroz, O.V.; Maranta, M.; Shaghasi, T.; Harris, P.V.; Wilson, K.S.; Davies, G.J. The three-dimensional structure of the cellobiohydrolase Cel7A from Aspergillus fumigatus at 1.5 Å resolution. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, D.; Teng, F. Purification and characterization of a cellobiohydrolase from the thermophilic fungus Chaetomium thermophilus CT2. Wei Sheng Wu Xue Bao 2006, 46, 143–146. [Google Scholar] [PubMed]
- Hobdey, S.E.; Knott, B.C.; Haddad Momeni, M.; Taylor, L.E.; Borisova, A.S.; Podkaminer, K.K.; VanderWall, T.A.; Himmel, M.E.; Decker, S.R.; Beckham, G.T.; et al. Biochemical and Structural Characterizations of Two Dictyostelium Cellobiohydrolases from the Amoebozoa Kingdom Reveal a High Level of Conservation between Distant Phylogenetic Trees of Life. Appl. Environ. Microbiol. 2016, 82, 3395–3409. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Iikura, H.; Nakamura, A.; Hidaka, M.; Masaki, H.; Uozumi, T. Isolation of the gene and characterization of the enzymatic properties of a major exoglucanase of Humicola grisea without a cellulose-binding domain. J. Biochem. 1998, 124, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Voutilainen, S.P.; Boer, H.; Linder, M.B.; Puranen, T.; Rouvinen, J.; Vehmaanperä, J.; Koivula, A. Heterologous expression of Melanocarpus albomyces cellobiohydrolase Cel7B, and random mutagenesis to improve its thermostability. Enzyme Microb. Technol. 2007, 41, 234–243. [Google Scholar] [CrossRef]
- Texier, H.; Dumon, C.; Neugnot-Roux, V.; Maestracci, M.; O’Donohue, M.J. Redefining XynA from Penicillium funiculosum IMI 378536 as a GH7 cellobiohydrolase. J. Ind. Microbiol. Biotechnol. 2012, 39, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Colussi, F.; Serpa, V.; Delabona, P.D.S.; Manzine, L.R.; Voltatodio, M.L.; Alves, R.; Mello, B.L.; Pereira, N.; Farinas, C.S.; Golubev, A.M.; et al. Purification, and biochemical and biophysical characterization of cellobiohydrolase I from Trichoderma harzianum IOC 3844. J. Microbiol. Biotechnol. 2011, 21, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Bronnenmeier, K.; Rücknagel, K.P.; Staudenbauer, W.L. Purification and properties of a novel type of exo-1,4-beta-glucanase (avicelase II) from the cellulolytic thermophile Clostridium stercorarium. Eur. J. Biochem. 1991, 200, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Z.; Zhang, Z.; Zhu, Z.; Sathitsuksanoh, N.; Yang, Y.; Zhang, Y.-H.P. The noncellulosomal family 48 cellobiohydrolase from Clostridium phytofermentans ISDg: Heterologous expression, characterization, and processivity. Appl. Microbiol. Biotechnol. 2010, 86, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Kruus, K.; Wang, W.K.; Ching, J.; Wu, J.H.D. Exoglucanase activities of the recombinant Clostridium thermocellum CelS, a major cellulosome component. J. Bacteriol. 1995, 177, 1641–1644. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Bevan, D.R.; Zhang, Y.-H.P. The family 1 glycoside hydrolase from Clostridium cellulolyticum H10 is a cellodextrin glucohydrolase. Appl. Biochem. Biotechnol. 2010, 161, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Yernool, D.A.; McCarthy, J.K.; Eveleigh, D.E.; Bok, J.D. Cloning and characterization of the glucooligosaccharide catabolic pathway β-glucan glucohydrolase and cellobiose phosphorylase in the marine hyperthermophile Thermotoga neapolitana. J. Bacteriol. 2000, 182, 5172–5179. [Google Scholar] [CrossRef] [PubMed]
- Kengen, S.W.M.; Luesink, E.J.; Stams, A.J.M.; Zehnder, A.J.B. Purification and characterization of an extremely thermostable β-glucosidase from the hyperthermophilic archaeon Pyrococcus furiosus. Eur. J. Biochem. 1993, 213, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, J.E.; Lee, H.S.; Kwon, K.K.; Kang, S.G.; Lee, J. Novel substrate specificity of a thermostable β-glucosidase from the hyperthermophilic archaeon, Thermococcus pacificus P-4. Korean J. Microbiol. 2015, 51, 68–74. [Google Scholar] [CrossRef]
- Matsui, I.; Sakai, Y.; Matsui, E.; Kikuchi, H.; Kawarabayasi, Y.; Honda, K. Novel substrate specificity of a membrane-bound beta-glycosidase from the hyperthermophilic archaeon Pyrococcus horikoshii. FEBS Lett. 2000, 467, 195–200. [Google Scholar] [CrossRef]
- Wu, Y.; Yuan, S.; Chen, S.; Wu, D.; Chen, J.; Wu, J. Enhancing the production of galacto-oligosaccharides by mutagenesis of Sulfolobus solfataricus β-galactosidase. Food Chem. 2013, 138, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.K.; Datta, S. β-Glucosidase from the hyperthermophilic archaeon Thermococcus sp. is a salt-tolerant enzyme that is stabilized by its reaction product glucose. Appl. Microbiol. Biotechnol. 2016, 100, 8399–8409. [Google Scholar] [CrossRef] [PubMed]
- Di Lauro, B.; Rossi, M.; Moracci, M. Characterization of a beta-glycosidase from the thermoacidophilic bacterium Alicyclobacillus acidocaldarius. Extremophiles 2006, 10, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, R.; Wang, J.; Zhang, X.; Jia, R.; Gao, Y.; Peng, H. Increased enzymatic hydrolysis of sugarcane bagasse by a novel glucose- and xylose-stimulated β-glucosidase from Anoxybacillus flavithermus subsp. yunnanensis E13T. BMC Biochem. 2017, 18, 4. [Google Scholar] [CrossRef] [PubMed]
- Paavilainen, S.; Hellman, J.; Korpela, T. Purification, characterization, gene cloning, and sequencing of a new β-glucosidase from Bacillus circulans subsp. alkalophilus. Appl. Environ. Microbiol. 1993, 59, 927–932. [Google Scholar] [PubMed]
- Xu, H.; Xiong, A.-S.; Zhao, W.; Tian, Y.-S.; Peng, R.-H.; Chen, J.-M.; Yao, Q.-H. Characterization of a glucose-, xylose-, sucrose-, and D-galactose-stimulated β-glucosidase from the alkalophilic bacterium Bacillus halodurans C-125. Curr. Microbiol. 2011, 62, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Love, D.R.; Fisher, R.; Bergquist, P.L. Sequence structure and expression of a cloned β-glucosidase gene from an extreme thermophile. MGG Mol. Gen. Genet. 1988, 213, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, A.; Arai, T.; Doi, R.H. Degradation of cellulosome-produced cello-oligosaccharides by an extracellular non-cellulosomal beta-glucan glucohydrolase, BglA, from Clostridium cellulovorans. Biochem. Biophys. Res. Commun. 2006, 349, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.-Z.; Yu, H.-L.; Li, C.-X.; Zhou, X.-W.; Hayashi, C.; Sun, J.; Liu, B.-H.; Imanaka, T.; Xu, J.-H. A new thermostable β-glucosidase mined from Dictyoglomus thermophilum: Properties and performance in octyl glucoside synthesis at high temperatures. Bioresour. Technol. 2012, 118, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, D.; Klippel, B.; Antranikian, G. A novel thermostable and glucose-tolerant β-glucosidase from Fervidobacterium islandicum. Appl. Microbiol. Biotechnol. 2012, 93, 1947–1956. [Google Scholar] [CrossRef] [PubMed]
- Gefen, G.; Anbar, M.; Morag, E.; Lamed, R.; Bayer, E.A. Enhanced cellulose degradation by targeted integration of a cohesin-fused β-glucosidase into the Clostridium thermocellum cellulosome. Proc. Natl. Acad. Sci. USA 2012, 109, 10298–10303. [Google Scholar] [CrossRef] [PubMed]
- Brognaro, H.; Almeida, V.M.; de Araujo, E.A.; Piyadov, V.; Santos, M.A.M.; Marana, S.R.; Polikarpov, I. Biochemical Characterization and Low-Resolution SAXS Molecular Envelope of GH1 β-Glycosidase from Saccharophagus degradans. Mol. Biotechnol. 2016, 58, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.R.; Coutinho, P.M.; Videira, P.; Fialho, A.M.; Sá-Correia, I. Sphingomonas paucimobilis β-glucosidase Bgl1: A member of a new bacterial subfamily in glycoside hydrolase family 1. Biochem. J. 2003, 370, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pons, J.A.; Cayetano, A.; Rebordosa, X.; Lloberas, J.; Guasch, A.; Querol, E. A beta-glucosidase gene (bgl3) from Streptomyces sp. strain QM-B814. Molecular cloning, nucleotide sequence, purification and characterization of the encoded enzyme, a new member of family 1 glycosyl hydrolases. Eur. J. Biochem. 1994, 223, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Breves, R.; Bronnenmeier, K.; Wild, N.; Lottspeich, F.; Staudenbauer, W.L.; Hofemeister, J. Genes encoding two different β-glucosidases of Thermoanaerobacter brockii are clustered in a common operon. Appl. Environ. Microbiol. 1997, 63, 3902–3910. [Google Scholar] [PubMed]
- Song, X.; Xue, Y.; Wang, Q.; Wu, X. Comparison of three thermostable β-glucosidases for application in the hydrolysis of soybean isoflavone glycosides. J. Agric. Food Chem. 2011, 59, 1954–1961. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Pang, Q.; Zhao, L.; Fan, S.; Shi, H. Thermoanaerobacterium thermosaccharolyticum β-glucosidase: A glucose-tolerant enzyme with high specific activity for cellobiose. Biotechnol. Biofuels 2012, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Spiridonov, N.A.; Wilson, D.B. Cloning and biochemical characterization of BglC, a beta-glucosidase from the cellulolytic actinomycete Thermobifida fusca. Curr. Microbiol. 2001, 42, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.M.; Yablonsky, M.D.; Shalita, Z.P.; Goyal, A.K.; Eveleigh, D.E. Cloning, characterization, and nucleotide sequence of a gene encoding Microbispora bispora BglB, a thermostable beta-glucosidase expressed in Escherichia coli. Appl. Environ. Microbiol. 1992, 58, 3455–3465. [Google Scholar] [PubMed]
- Haq, I.U.; Khan, M.A.; Muneer, B.; Hussain, Z.; Afzal, S.; Majeed, S.; Rashid, N.; Javed, M.M.; Ahmad, I. Cloning, characterization and molecular docking of a highly thermostable β-1,4-glucosidase from Thermotoga petrophila. Biotechnol. Lett. 2012, 34, 1703–1709. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.-J.; Lee, Y.-J.; Chol, J.J.; Seo, M.S.; Lee, M.S.; Kim, G.A.; Kwon, S.-T. Mutational analysis of Thermus caldophilus GK24 beta-glycosidase: Role of His119 in substrate binding and enzyme activity. J. Microbiol. Biotechnol. 2008, 18, 287–294. [Google Scholar] [PubMed]
- Xiangyuan, H.; Shuzheng, Z.; Shoujun, Y. Cloning and expression of thermostable beta-glycosidase gene from Thermus nonproteolyticus HG102 and characterization of recombinant enzyme. Appl. Biochem. Biotechnol. 2001, 94, 243–255. [Google Scholar] [CrossRef]
- Kang, S.K.; Cho, K.K.; Ahn, J.K.; Bok, J.D.; Kang, S.H.; Woo, J.H.; Lee, H.G.; You, S.K.; Choi, Y.J. Three forms of thermostable lactose-hydrolase from Thermus sp. IB-21: Cloning, expression, and enzyme characterization. J. Biotechnol. 2005, 116, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Nam, E.S.; Kim, M.S.; Lee, H.B.; Ahn, J.K. β-Glycosidase of Thermus thermophilus KNOUC202: Gene and biochemical properties of the enzyme expressed in Escherichia coli. Appl. Biochem. Microbiol. 2010, 46, 515–524. [Google Scholar] [CrossRef]
- Zhao, Z.; Ramachandran, P.; Kim, T.-S.; Chen, Z.; Jeya, M.; Lee, J.-K. Characterization of an acid-tolerant β-1,4-glucosidase from Fusarium oxysporum and its potential as an animal feed additive. Appl. Microbiol. Biotechnol. 2013, 97, 10003–10011. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Nakamura, A.; Hidaka, M.; Masaki, H.; Uozumi, T. Molecular Cloning and Expression of the Novel Fungal -Glucosidase Genes from Humicola grisea and Trichoderma reesei. J. Biochem. 1999, 125, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-L.; Ljungdahl, L.G.; Ximenes, E.A.; Chen, H.; Felix, C.R.; Cotta, M.A.; Dien, B.S. Properties of a recombinant beta-glucosidase from polycentric anaerobic fungus Orpinomyces PC-2 and its application for cellulose hydrolysis. Appl. Biochem. Biotechnol. 2004, 113, 233–250. [Google Scholar] [CrossRef]
- Li, X.; Zhao, J.; Shi, P.; Yang, P.; Wang, Y.; Luo, H.; Yao, B. Molecular cloning and expression of a novel β-glucosidase gene from Phialophora sp. G5. Appl. Biochem. Biotechnol. 2013, 169, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Schröder, C.; Blank, S.; Antranikian, G. First Glycoside Hydrolase Family 2 Enzymes from Thermus antranikianii and Thermus brockianus with β-Glucosidase Activity. Front. Bioeng. Biotechnol. 2015, 3, 76. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, X.; Dang, W.; Tran, P.L.; Park, S.-H.; Oh, B.-C.; Hong, W.-S.; Lee, J.-S.; Park, K.-H. Characterization and application of an acidophilic and thermostable β-glucosidase from Thermofilum pendens. J. Biosci. Bioeng. 2013, 115, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wei, P.; Chen, Q.; Chen, X.; Wang, S.; Li, J.; Gao, C. Functional and structural characterization of a β-glucosidase involved in saponin metabolism from intestinal bacteria. Biochem. Biophys. Res. Commun. 2018, 496, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.T.Y.; Wong, W.K.R. Purification and characterization of a major secretory cellobiase, Cba2, from Cellulomonas biazotea. Protein Expr. Purif. 2001, 23, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wakarchuk, W. Characterization of five β-glycoside hydrolases from Cellulomonas fimi ATCC 484. J. Bacteriol. 2014, 196, 4103–4110. [Google Scholar] [CrossRef] [PubMed]
- Bronnenmeier, K.; Staudenbauer, W.L. Purification and properties of an extracellular β-glucosidase from the cellulolytic thermophile Clostridium stercorarium. Appl. Microbiol. Biotechnol. 1988, 28, 380–386. [Google Scholar] [CrossRef]
- Li, Y.-K.; Lee, J.-A. Cloning and expression of β-glucosidase from Flavobacterium meningosepticum: A new member of family B β-glucosidase. Enzyme Microb. Technol. 1999, 24, 144–150. [Google Scholar] [CrossRef]
- Xie, J.; Zhao, D.; Zhao, L.; Pei, J.; Xiao, W.; Ding, G.; Wang, Z. Overexpression and characterization of a Ca2+ activated thermostable β-glucosidase with high ginsenoside Rb1 to ginsenoside 20(S)-Rg3 bioconversion productivity. J. Ind. Microbiol. Biotechnol. 2015, 42, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Colabardini, A.C.; Valkonen, M.; Huuskonen, A.; Siika-aho, M.; Koivula, A.; Goldman, G.H.; Saloheimo, M. Expression of Two Novel β-Glucosidases from Chaetomium atrobrunneum in Trichoderma reesei and Characterization of the Heterologous Protein Products. Mol. Biotechnol. 2016, 58, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, R.; Yang, X.; Zhang, Z.; Song, S.; Miao, Y.; Shen, Q. Characterization of a thermostable β-glucosidase from Aspergillus fumigatus Z5, and its functional expression in Pichia pastoris X33. Microb. Cell Fact. 2012, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Teng, F.; Zhang, C.; Li, D. Cloning of a Gene Encoding β-Glucosidase from Chaetomium thermophilum CT2 and Its Expression in Pichia pastoris. J. Mol. Microbiol. Biotechnol. 2011, 20, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Dotsenko, G.S.; Sinitsyna, O.A.; Hinz, S.W.A.; Wery, J.; Sinitsyn, A.P. Characterization of a GH family 3 β-glycoside hydrolase from Chrysosporium lucknowense and its application to the hydrolysis of β-glucan and xylan. Bioresour. Technol. 2012, 112, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Zhao, J.; Cai, P.; Sun, W.; Ren, J.; Wu, Q.; Zhang, S.; Tian, C. Heterologous expression of a GH3 β-glucosidase from Neurospora crassa in Pichia pastoris with high purity and its application in the hydrolysis of soybean isoflavone glycosides. Protein Expr. Purif. 2016, 119, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Nomura, T.; Ogita, S.; Takano, M.; Hoshino, K. Two new β-glucosidases from ethanol-fermenting fungus Mucor circinelloides NBRC 4572: Enzyme purification, functional characterization, and molecular cloning of the gene. Appl. Microbiol. Biotechnol. 2013, 97, 10045–10056. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ma, R.; Shi, P.; Huang, H.; Bai, Y.; Wang, Y.; Yang, P.; Fan, Y.; Yao, B. Molecular characterization of a highly-active thermophilic β-glucosidase from Neosartorya fischeri P1 and its application in the hydrolysis of soybean isoflavone glycosides. PLoS ONE 2014, 9, e106785. [Google Scholar] [CrossRef] [PubMed]
- Krogh, K.B.R.M.; Harris, P.V.; Olsen, C.L.; Johansen, K.S.; Hojer-Pedersen, J.; Borjesson, J.; Olsson, L. Characterization and kinetic analysis of a thermostable GH3 β-glucosidase from Penicillium brasilianum. Appl. Microbiol. Biotechnol. 2010, 86, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Qin, Y.; Liu, Z.; Liu, K.; Wang, F.; Qu, Y. Isolation and characterization of a β-glucosidase from Penicillium decumbens and improving hydrolysis of corncob residue by using it as cellulase supplementation. Enzyme Microb. Technol. 2010, 46, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Ohtsuki, I.; Fukui, S.; Yamashita, I. Nucleotide sequences of Saccharomycopsis fibuligera genes for extracellular β-glucosidases as expressed in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1988, 54, 3147–3155. [Google Scholar] [PubMed]
- Murray, P.; Aro, N.; Collins, C.; Grassick, A.; Penttilä, M.; Saloheimo, M.; Tuohy, M. Expression in Trichoderma reesei and characterisation of a thermostable family 3 beta-glucosidase from the moderately thermophilic fungus Talaromyces emersonii. Protein Expr. Purif. 2004, 38, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Xu, X.; Qian, L.; Shi, P.; Bai, Y.; Luo, H.; Ma, R.; Yao, B. Engineering a highly active thermophilic β-glucosidase to enhance its pH stability and saccharification performance. Biotechnol. Biofuels 2016, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Tamaki, H.; Kumagai, H. Cloning and functional expression of thermostable beta-glucosidase gene from Thermoascus aurantiacus. Appl. Microbiol. Biotechnol. 2007, 73, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Karnaouri, A.; Topakas, E.; Paschos, T.; Taouki, I.; Christakopoulos, P. Cloning, expression and characterization of an ethanol tolerant GH3 β-glucosidase from Myceliophthora thermophila. PeerJ 2013, 1, e46. [Google Scholar] [CrossRef] [PubMed]
- Brunner, F.; Wirtz, W.; Rose, J.K.C.; Darvill, A.G.; Govers, F.; Scheel, D.; Nürnberger, T. A β-glucosidase/xylosidase from the phytopathogenic oomycete, Phytophthora infestans. Phytochemistry 2002, 59, 689–696. [Google Scholar] [CrossRef]
- Ferrara, M.C.; Cobucci-Ponzano, B.; Carpentieri, A.; Henrissat, B.; Rossi, M.; Amoresano, A.; Moracci, M. The identification and molecular characterization of the first archaeal bifunctional exo-β-glucosidase/N-acetyl-β-glucosaminidase demonstrate that family GH116 is made of three functionally distinct subfamilies. Biochim. Biophys. Acta 2014, 1840, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Sansenya, S.; Mutoh, R.; Charoenwattanasatien, R.; Kurisu, G.; Ketudat Cairns, J.R. Expression and crystallization of a bacterial glycoside hydrolase family 116 β-glucosidase from Thermoanaerobacterium xylanolyticum. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 41–44. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Enzyme | GH Family Domains | Optimum Temperature | Optimum pH | Temperature Stability 1 | Source | Reference |
|---|---|---|---|---|---|---|
| EGPh | 5 | >97 °C | 5.4–6.0 | 80%; 97 °C; 3 h | Archaea (Pyrococcus horikoshii) | [46] |
| EG1 | 5 | 83 °C | 5.0 | 20%; 90 °C; 2 h | Bacteria (Acidothermus cellolyticus) | [73] |
| EglII | 5 | 50 °C | 6.0 | NM | Bacteria (Bacillus amyloliquefaciens) | [74] |
| EG | 5 | 65 °C | 6.0 | 72%; 55 °C; 42 h 50%; 65 °C; 12 min | Bacteria (Bacillus licheniformis) | [75] |
| CelA | 5 | 60 °C | 8.0 | 30%; 70 °C; 1 h | Bacteria (Bacillus subtilis) | [76] |
| TmCel5A | 5 | 80 °C | 6.0 | 50%; 80 °C; 18 h | Bacteria (Thermotoga maritima) | [77] |
| EglA | 5 | 57 °C | 4.0 | NM | Fungi (Aspergillus nidulans) | [78] |
| EglB | 5 | 52 °C | 4.0 | NM | Fungi (Aspergillus nidulans) | [78] |
| EBI-244 | 5 | 109 °C | 5.5 | 50%; 100 °C; 4.5 h 50%; 105 °C; 0.57 h 108 °C; 50%; 0.17 h | Uncultured Archaea (Continental geothermal pool enrichment) | [58] |
| CelE1 | 5 | 50 °C | 7.0 | NM | Uncultured organism (Sugarcane field soil metagenome) | [64] |
| CelA10 | 5 | 55 °C | 7.5 | NM | Uncultured organism (Aquatic community and soil sample) | [66] |
| CelA24 | 5 | 55 °C | 7.0 | NM | Uncultured organism (Aquatic community and soil sample) | [66] |
| cMGL504 | 5 | 50 °C | 5.5 | NM | Uncultured organism (Vermicompost sample) | [60] |
| Cel5G | 5 | 50 °C | 4.8 | >90%; 50 °C; 30 min | Uncultured organism (Soil metagenome) | [65] |
| En1 | 5 | 55 °C | 5.5 | 87%; 45 °C; 16 h 67%; 50 °C; 6 h 42%; 55 °C; 30 min | Uncultured organism (Biogas digester metagenome) | [63] |
| RC1 | 5 | 55 °C | 6.0–6.5 | >90%; 50 °C; 30 min | Uncultured organism (Rabbit cecum metagenome) | [67] |
| RC3 | 5 | 50 °C | 6.0–7.0 | NM | Uncultured organism (Rabbit cecum metagenome) | [67] |
| RC5 | 5 | 50 °C | 6.5–7.0 | NM | Uncultured organism (Rabbit cecum metagenome) | [67] |
| CelL | 6 | 50 °C | 5.0 | 50%; 50 °C; 12 min | Bacteria (Cellulosimicrobium funkei) | [22] |
| Cel6A | 6 | 58 °C | 6.5 | >80%; 56 °C; 18 h | Bacteria (Thermobifida fusca) | [79] |
| ThCel6A | 6 | 55 °C | 8.5 | 58%; 90 °C; 1 h | Bacteria (Thermobifida halotolerans) | [80] |
| Cel6A | 6 | 50–55 °C | 5.5 | NM | Bacteria (Xylanimicrobium pachnodae) | [81] |
| HiCel6C | 6 | 70 °C | 6.5 | >90%; 60 °C; 1 h | Fungi (Humicola insolens) | [82] |
| Cel6A | 6 | 50 °C | 4.8 | >90%; 45 °C; 24 h 92%; 50 °C; 5 h | Fungi (Orpinomyces sp.) | [83] |
| C1 | 6 | 50 °C | 6.0 | 100%; 60 °C; 30 min | Uncultured organism (Compost metagenome) | [61] |
| pre-LC-CelB | 6 | NM | NM | NM | Uncultured organism (Compost metagenome) | [62] |
| pre-LC-CelJ | 6 | NM | NM | NM | Uncultured organism (Compost metagenome) | [62] |
| EGI | 7 | 55–60 °C | 5.0 | >80%; 60 °C; 10 min | Fungi (Humicola grisea var. thermoidea) | [84] |
| Cel7B | 7 | 60 °C | 4.0 | >90%; 60 °C; 1 h | Fungi (Penicillium decumbens) | [85] |
| Cel7A | 7 | 60 °C | 5.0 | 100%; 60 °C; 1 h 16.1%; 70 °C; 1 h | Fungi (Neosartorya fischeri) | [33] |
| MtEG7 | 7 | 60 °C | 5.0 | 50%; 70 °C; 9.96 h 50%; 80 °C; 6.5 h | Fungi (Myceliophthora thermophila) | [31] |
| EGL1 | 7 | 62 °C | 4.8 | NM | Fungi (Trichoderma longibrachiatum) | [51] |
| MaCel7A | 7 | 65–70 °C | 6.0 | NM | Fungi (Melanocarpus albomyces) | [86] |
| CelC | 8 | 50 °C | 6.5 | NM | Bacteria (Salmonella typhimurium) | [87] |
| Cel8Y | 8 | 80 °C | 7.0 | 50%; 90 °C; 4 h 50%; 100 °C; 2 h | Bacteria (Aquifex geolicus) | [88] |
| Egl-257 | 8 | 55 °C | 8.5 | 100%; 50 °C; 15 min >50%; 60 °C; 15 min | Bacteria (Bacillus circulans) | [89] |
| CenC | 9 | 70 °C | 6.0 | 100%; 60 °C; 2 h 60%; 70 °C; 1 h | Bacteria (Clostridium thermocellum) | [90] |
| CelA | 9 (endoglucanase) and 48 (cellobiohydrolase) | 95 °C (endoglucanase) and 85 °C (cellobiohydrolase) | 5.0–6.0 | 50%; 95 °C; 40 min (endoglucanase) 100%; 85 °C; 4 h (cellobiohydrolase) | Bacteria (Caldicellulosiruptor bescii) | [91] |
| Cel9A | 9 | 65 °C | 6.5 | NM | Bacteria (Lachnoclostridium phytofermentans) | [92] |
| CelA20 | 9 | 55 °C | 5.0 | NM | Uncultured organism (Aquatic community and soil metagenome) | [66] |
| AcCel12B | 12 | 75 °C | 4.5 | 50%; 60 °C; 90 h 50%; 65 °C; 55 h 50%; 70 °C; 2 h | Bacteria (Acidothermus cellulolyticus) | [35] |
| CelA | 12 | 95 °C | 6.0 | NM | Bacteria (Thermotoga neapolitana) | [8] |
| CelB | 12 | 106 °C | 6.0–6.6 | 50%; 106 °C; 130 min 50%; 110 °C; 26 min 73%; 100 °C; 4 h | Bacteria (Thermotoga neapolitana) | [8] |
| TmCel12A | 12 | 90 °C | 7.0 | >40%; 85 °C; 48 h 50%; 90 °C; 3 h | Bacteria (Thermotoga maritima) | [93] |
| TmCel12B | 12 | 85 °C | 6.0 | 50%; 90 °C; 9 h | Bacteria (Thermotoga maritima) | [93] |
| CelA | 12 | >100 °C | 6.0–7.0 | 45%; 90 °C; 8 h | Bacteria (Rhodothermus marinus) | [23] |
| EglA | 12 | 100 °C | 6.0 | 50%; 95 °C; 40 h | Archaea (Pyrococcus furiosus) | [94] |
| SSO1949 | 12 | 80 °C | 1.8 | 50%; 80 °C; 8 h | Archeaea (Sulfolobus solfataricus) | [95] |
| SSO1354 | 12 | 90 °C | 4.0 | 50%; 90 °C; 180 min | Archaea (Sulfolobus solfataricus) | [39] |
| EglS | 12 | 65 °C | 6.0 | >40%; 60 °C; 30 min | Bacteria (Streptomyces rochei) | [96] |
| Cel12A | 12 | 50 °C | 5.0 | NM | Fungi (Trichoderma reseei) | [97] |
| EG | 12 | 70 °C | 3.5 | 50%; 70 °C; 3 h 50%; 80 °C; 1 h | Fungi (Aspergillus niger) | [27] |
| Pre-LC-CelA | 12 | 90 °C | 5.0–9.0 | 100%; 90 °C; 30 min | Uncultured organism (Compost metagenome) | [62] |
| Pre-LC-CelD | 12 | NM | NM | NM | Uncultured organism (Compost metagenome) | [62] |
| Pre-LC-CelE | 12 | NM | NM | NM | Uncultured organism (Compost metagenome) | [62] |
| Cel12E | 12 | 92 °C | 5.5 | >80%; 80 °C; 4.5 h | Uncharacterized archeon (deep sea vents metagenome enrichment) | [57] |
| GH44EG | 44 | 55 °C | 5.0 | NM | Bacteria (Clostridium acetobutylicum) | [98] |
| CelA | 44 | 60 °C | 5.0–8.5 | 50%; 60 °C; 70 min | Bacteria (Paenibacillus lautus) | [99] |
| CelJ | 44 | 70 °C | 6.5 | >90%; 80 °C; 10 min | Bacteria (Ruminiclostridium thermocellum) | [100] |
| pre-LC-CelH | 44 | NM | NM | NM | Uncultured organism (Compost metagenome) | [62] |
| Cel45A | 45 | 60 °C | 5.0 | NM | Fungi (Trichoderma reseei) | [97] |
| PpCel45A | 45 | 65 °C | 4.8 | 70%; 65 °C; 48 h 60%; 80 °C; 4 h | Fungi (Picchia pastoris) | [5] |
| STCE1 | 45 | 60 °C | 6.0 | NM | Fungi (Staphylotrichum coccosporum) | [101] |
| BCC18080 | 45 | 70 °C | 6.0 | >70%; 70 °C; 2 h >50%; 70 °C; 4 h | Fungi (Syncephalastrum racemosum) | [102] |
| BCE1 | 45 | 55 °C | 4.5 | NM | Fungi (Beltraniella portoricensis) | [103] |
| MaCel45A | 45 | 70 °C | 7.0 | NM | Fungi (Melanocarpus albomyces) | [86] |
| CelB | 51 | 80 °C | 4.0 | 60%; 80 °C; 1 h | Bacteria (Alicyclobacillus acidocaldarius) | [104] |
| CelA4 | 51 | 65 °C | 2.6 | >85%; 60 °C; 1 h | Bacteria (Alicyclobacillus sp. A4) | [47] |
| CelVA | 51 | 80 °C | 3.6–4.5 | 70%; 70 °C; 2 h | Bacteria (Alicyclobacillus vulcanalis) | [45] |
| pre-LC-CelC | 51 | NM | NM | NM | Uncultured organism (Compost metagenome) | [62] |
| TmCel74 | 74 | 90 °C | 6.0 | 50%; 90 °C; 5 h | Bacteria (Thermotoga maritima) | [15] |
| CtCel124 | 124 | NM | NM | NM | Bacteria (Ruminiclostridium thermocellum) | [105] |
| Enzyme | GH Family Domains | Optimum Temperature | Optimum pH | Temperature Stability 1 | Source | Reference |
|---|---|---|---|---|---|---|
| CBHII | 6 | 60 °C | 4.0 | 30%; 100 °C; 10 min | Bacteria (Streptomyces sp. M23) | [106] |
| Cel6B | 6 | NM | 7.0–8.0 | 100%; 55 °C; 16 h | Bacteria (Thermobifida fusca) | [107] |
| CBHII | 6 | 57 °C | 5.5 | NM | Fungi (Aspergillus nidulans) | [78] |
| Cel6A | 6 | 50 °C | 4.0 | 50%; 70 °C; 30 min | Fungi (Chaetomium thermophilum) | [108] |
| CBHII (Cel6A) | 6 | 60 °C | 5.0–5.5 | >90%; 50 °C; 5 h | Fungi (Chrysosporium lucknowense) | [109] |
| HiCel6A | 6 | 60–65 °C | NM | 50%; 75 °C; <25 min | Fungi (Humicola insolens) | [26] |
| Ex-4 | 6 | 50 °C | 5.0 | 80%; 60 °C; 60 min | Fungi (Irpex Lacteus) | [110] |
| PoCel6A | 6 | 50 °C | 5.0 | 90%; 50 °C; 2 h 80%; 60 °C; 4 h | Fungi (Penicillium oxalicum) | [111] |
| PaCel6A | 6 | 55 °C | 5.0–9.0 | 100%; 35 °C; 24 h >20%; 45 °C; 24 h | Fungi (Podospora anserina) | [19] |
| CBHII | 6 | 70 °C | 5.0 | NM | Fungi (Trichoderma viride) | [112] |
| G10-6 | 6 | 55 °C | 9.5 | NM | Uncultured organism (Eathworm casts metagenome) | [70] |
| Cbh9A | 9 | 60 °C | 6.5 | NM | Bacteria (Ruminiclostridium thermocellum) | [113] |
| Cel9K | 9 | 65 °C | 6.0 | 97%; 60 °C; 200 h | Bacteria (Ruminiclostridium thermocellum) | [114] |
| Enzyme | GH Family Domains | Optimum Temperature | Optimum pH | Temperature Stability 1 | Source | Reference |
|---|---|---|---|---|---|---|
| CelO | 5 | 65 °C | 6.6 | NM | Bacteria (Ruminiclostridium thermocellum) | [115] |
| AtCel7A | 7 | 60 °C | 5.0 | NM | Fungi (Acremonium thermophilum) | [28] |
| CBHI | 7 | 60 °C | 3.0 | NM | Fungi (Aspergillus aculeatus) | [116] |
| CBHI | 7 | 55 °C | NM | NM | Fungi (Aspergillus fumigatus) | [117] |
| CtCel7A | 7 | 65 °C | 4.0 | NM | Fungi (Chaetomium thermophilum) | [28] |
| CBH3 | 7 | 65 °C | 5.0 | 50%; 70 °C; 1 h 20%; 80 °C; 20 min | Fungi (Chaetomium thermophilum) | [118] |
| DpuCel7A | 7 | 55 °C | 5.0 | NM | Metazoa (Dictyostelium purpureum) | [119] |
| CBHI | 7 | 60 °C | 5.0 | >90%; 55 °C; 10 min | Fungi (Humicola grisea var. thermoidea) | [84] |
| EXO1 | 7 | 65 °C | 5.0 | >80%; 65 °C; 10 min | Fungi (Humicola grisea var. thermoidea) | [120] |
| MaCel7B | 7 | 55 °C | NM | NM | Fungi (Melanocarpus albomyces) | [121] |
| TeCel7A | 7 | 65 °C | 4.0–5.0 | 50%; 70 °C; 30 min | Fungi (Talaromyces emersonii) | [29] |
| Cel7A | 7 | 55 °C | 3.7–5.2 | 50%; 50 °C; 2.5 h | Fungi (Penicillium funiculosum) | [122] |
| TaCel7A | 7 | 65 °C | 5.0 | NM | Fungi (Thermoascus aurantiacus) | [28] |
| ThCBHI | 7 | 50 °C | 5.0 | NM | Fungi (Trichoderma harzianum) | [123] |
| CBHI | 7 | 60 °C | 5.8 | NM | Fungi (Trichoderma viride) | [112] |
| CelA | 9 (endoglucanase) and 48 (cellobiohydrolase) | 95 °C (endoglucanase) and 85 °C (cellobiohydrolase) | 5.0–6.0 | 50%; 95 °C; 40 min (endoglucanase) 100%; 85 °C; 4 h (cellobiohydrolase) | Bacteria (Caldicellulosiruptor bescii) | [91] |
| CelY | 48 | 70 °C | 5.0–6.0 | NM | Bacteria (Clostridium stercorarium) | [124] |
| CpCel48 | 48 | 55 °C | 5.0–6.0 | >70%; 50 °C; 30 min>20%; 55 °C; 30 min | Bacteria (Lachnoclostridium phytofermentans) | [125] |
| CelS | 48 | 70 °C | 5.5 | NM | Bacteria (Ruminiclostridium thermocellum) | [126] |
| Enzyme | GH family Domains | Optimum Temperature | Optimum pH | Temperature Stability 1 | Source | Reference |
|---|---|---|---|---|---|---|
| CcGH1 | 1 | 60 °C | 6.5 | 61%; 50 °C; 30 min | Bacteria (Clostridium Cellulolyticum) | [127] |
| GghA | 1 | 95 °C | 6.5 | 85%; 90 °C; 9 h 88%; 95 °C; 1 h | Bacteria (Thermotoga neapolitana) | [128] |
| Enzyme | GH family Domains | Optimum Temperature | Optimum pH | Temperature Stability 1 | Source | Reference |
|---|---|---|---|---|---|---|
| CelB | 1 | 102–105 °C | 5.0 | 50%; 100 °C; 85 h 50%; 110 °C; 13 h | Arquea (Pyrococcus furiosus) | [129] |
| Tpa-glu | 1 | 75 °C | 7.5 | 50%; 90 °C; 6 h | Arquea (Thermococcus pacificus) | [130] |
| BGPh | 1 | >100 °C | 6.0 | 50%; 90 °C; 15 h | Arquea (Pyrococcus horikoshii) | [131] |
| LacS | 1 (β-glucosidase and β-galactosidase) | 90 °C | 6.0 | 90%; 75 °C; 80 h | Arquea (Sulfolobus solfataricus) | [132] |
| O08324 | 1 | 78 °C | 5.0–6.8 | 50%; 78 °C; 860 min | Arquea (Thermococcus sp.) | [133] |
| Bgl1 | 1 | 90 °C | 6.5 | 67%; 90 °C; 1.5 h 78%; 50 °C; 24 h 68%; 60 °C; 24 h | Uncultured Arquea (hot spring metagenome) | [59] |
| GlyB | 1 (multiple substrates) | 85 °C | 5.5 | 8%; 80 °C; 10 min >70%; 65 °C; 3 h | Bacteria (Alicyclobacillus acidocaldarius) | [134] |
| Bglp | 1 | 60 °C | 7.0 | 50%; 60 °C; 10 h | Bacteria (Anoxybacillus flavithermus) | [135] |
| BglA | 1 | 55 °C | 6.0–9.0 | 80%; 50 °C; 15 min 1%; 60 °C; 15 min | Bacteria (Bacillus circulans subsp. Alkalophilus) | [136] |
| BhbglA | 1 | 50 °C | 7.0 | 50%; 50 °C; 30 min | Bacteria (Bacillus halodurans) | [137] |
| BglA | 1 | 85 °C | 6.25 | 50%; 70 °C; 2280 min | Bacteria (Caldicellulosiruptor saccharolyticus) | [138] |
| BglA | 1 | 50 °C | 6.0 | NM | Bacteria (Clostridium cellulovorans) | [139] |
| DtGH | 1 | 90 °C | 7.0 | 50%; 70 °C; 533 h 50%; 80 °C; 44 h 50%; 90 °C; 5 h | Bacteria (Dictyoglomus thermophilum) | [140] |
| DturβGlu | 1 | 80 °C | 5.4 | 70%; 70 °C; 2 h | Bacteria (Dictyoglomus turgidum) | [44] |
| FiBgl1A | 1 | 90 °C | 6.0–7.0 | 50%; 90 °C 25 min 50%; 100 °C; 15 min | Bacteria (Fervidobacterium islandicum) | [141] |
| BglA | 1 | 60 °C | 6.5 | 91%; 60 °C; 3 h 34%; 60 °C; 43 h | Bacteria (Ruminiclostridium thermocellum) | [142] |
| SdBgl1B | 1 | 50 °C | 6.0–7.5 | NM | Bacteria (Saccharophagus degradans) | [143] |
| Bgl1 | 1 | 50 °C | 5.1–5.7 | 60%; 40 °C; 4 h | Bacteria (Sphingomonas paucimobilis) | [144] |
| SGR_2426 | 1 | 69 °C | 6.9 | 50%; 69 °C; 1.5 h | Bacteria (Streptomyces griseus) | [55] |
| Bgl3 | 1 | 50 °C | 6.5 | NM | Bacteria (Streptomyces sp. strain QM-B814) | [145] |
| CglT | 1 | 75 °C | 5.5 | 100%; 60 °C; 24 h | Bacteria (Thermoanaerobacter brockii) | [146] |
| TeBglA | 1 | 80 °C | 7.0 | 10%; 65 °C; 5 h | Bacteria (Thermoanaerobacter ethanolicus) | [147] |
| TmBglA | 1 | 90 °C | 6.2 | >80%; 65 °C; 5 h | Bacteria (Thermotoga maritima) | [147] |
| Bgl | 1 | 70 °C | 6.4 | 50%; 68 °C; 1 h >80%; 60 °C; 2 h | Bacteria (Thermoanaerobacterium thermosaccharolyticum) | [148] |
| BglC | 1 | 50 °C | 7.0 | NM | Bacteria (Thermobifida fusca) | [149] |
| BglB | 1 | 60 °C | 6.2 | 70%; 60 °C; 48 h | Bacteria (Thermobispora bispora) | [150] |
| BglA | 1 | 80–90 °C | 7.0–8.0 | 100%; 70 °C; 6 h | Bacteria (Thermotoga petrophila) | [151] |
| TcaBglA | 1 | 90 °C | 5.5–6.5 | >40%; 80 °C; 30 min >20%; 80 °C; 30 min | Bacteria (Thermus caldophilus) | [152] |
| TnGly | 1 | 90 °C | 5.6 | 50%; 90 °C; 2.5 h | Bacteria (Thermus nonproteolyticus) | [153] |
| BglA | 1 | 70 °C | 5.0–6.0 | 50%; 70 °C; 38 h 50%; 80 °C; <0.4 h 50%; 90 °C; <0.3 h | Bacteria (Thermus sp. IB-21) | [154] |
| BglB | 1 | 80 °C | 5.0–6.0 | 50%; 70 °C; 38 h 50%; 80 °C; 2.7 h 50%; 90 °C; 24 min | Bacteria (Thermus sp. IB-21) | [154] |
| Bgly | 1 | 90 °C | 5.4 | 100%; 80 °C; 2 h 50%; 90 °C; 1.5 h 50%; 95 °C; 20 min | Bacteria (Thermus thermophilus) | [155] |
| BglA | 1 | 55 °C | 6.5 | 82%; 50 °C; 60 min 20%; 55 °C; 60 min | Uncultured organism (soil metagenome) | [52] |
| AS-Esc10 | 1 | 60 °C | 8.0 | 100%; 50 °C; 1 h | Uncultured organism (agricultural soil metagenome) | [40] |
| Bgl-gs1 | 1 | 90 °C | 6.0 | 50%; 90 °C; 5 min 50%; 85 °C; 15 min 50%; 80 °C; 45 min | Uncultured organism (termite gut metagenome) | [71] |
| Bgl | 1 | 60 °C | 5.0 | 50%; 60 °C; 540 min | Fungi (Fusarium oxysporum) | [156] |
| Bgl4 | 1 | 55 °C | 6.0 | 80%; 50 °C; 10 min | Fungi (Humicola grisea var. thermoidea IFO9854) | [157] |
| Bgl1 | 1 | 55 °C | 5.5–7.5 | 100%; 50 °C; 8 h 50%; 55 °C; 8 h | Fungi (Orpinomyces sp. PC-2) | [158] |
| Bgl1G5 | 1 | 50 °C | 6.0 | 50%; 50 °C; 6 h | Fungi (Phialophora sp. G5) | [159] |
| TaGH2 | 2 | 95 °C | 6.5 | 100%; 90 °C; 3 h 50%; 70 °C; 22 h | Bacteria (Thermus antranikianii) | [160] |
| TbGH2 | 2 | 90 °C | 6.5 | 17%; 80 °C; 3 h 50%; 70 °C; 12 h | Bacteria (Thermus brockianus) | [160] |
| TbBgl | 3 | 90 °C | 3.5 | 50%; 95 °C; 60 min | Arquea (Thermofilum pendens) | [161] |
| BlBG3 | 3 | 50 °C | 6.0 | NM | Bacteria (Bifidobacterium longum) | [162] |
| Cba2 | 3 | 70 °C | 4.8 | NM | Bacteria (Cellulomonas biazotea) | [163] |
| CfBgl3A | 3 | 55 °C | 7.5 | NM | Bacteria (Cellulomonas fimi) | [164] |
| Bgl3Z | 3 | 65 °C | 5.5 | 50%; 60 °C; 5 h | Bacteria (Clostridium stercorarium) | [165] |
| Dtur_0219 | 3 | 85 °C | 5.0 | 50%; 70 °C; 1575 min 50%; 75 °C; 854 min 50%; 80 °C; 524 min 50%; 85 °C; 334 min 50%; 90 °C; 20 min | Bacteria (Dictyoglomus turgidum) | [54] |
| Bgl | 3 | 50 °C | 4.2–5.0 | NM | Bacteria (Elizabethkingia meningoseptica) | [166] |
| TmBglB | 3 | 80 °C | 4.2 | >80%; 65 °C; 5 h | Bacteria (Thermotoga maritima) | [147] |
| Tpebgl3 | 3 | 90 °C | 5.0 | >90%; 70 °C; 3 h >50%; 90 °C; 3 h | Bacteria (Thermotoga petrophila) | [167] |
| Cel3A | 3 | 50–60 °C | 5.0 | 98%; 60 °C; 6 h >50%; 60 °C; 24 h >50%; 70 °C; 24 h | Fungi (Amesia atrobrunnea) | [168] |
| Cel3B | 3 | 50–60 °C | 5.0 | 88%; 60 °C; 6 h >50%; 60 °C; 24 h >50%; 70 °C; 24 h | Fungi (Amesia atrobrunnea) | [168] |
| Bgl3 | 3 | 60 °C | 6.0 | >50%; 70 °C; 1 h | Fungi (Aspergillus fumigatus) | [169] |
| BglB | 3 | 52 °C | 5.5 | NM | Fungi (Aspergillus nidulans) | [78] |
| BglC | 3 | 52 °C | 6.0 | NM | Fungi (Aspergillus nidulans) | [78] |
| Bgl | 3 | 50 °C | 5.0 | 100%; 50 °C; 30 min 60%; 60 °C; 30 min | Fungi (Aspergillus oryzae) | [49] |
| Bgl | 3 | 60 °C | 5.0 | 67.7%; 60 °C; 1 h 50%; 65 °C; 55 min 29.7%; 70 °C; 10 min | Fungi (Chaetomium thermophilum) | [170] |
| Bxl5 | 3 | 75 °C | 4.6 | 50%; 65 °C; 5 h 50%; 70 °C; 20 min 50%; 75 °C; 5 min | Fungi (Chrysosporium lucknowense) | [171] |
| MoCel3A | 3 | 50 °C | 5.0–5.5 | NM | Fungi (Magnaporthe oryzae) | [41] |
| MoCel3B | 3 | 50 °C | 5.0–5.5 | NM | Fungi (Magnaporthe oryzae) | [41] |
| Bgl2 | 3 | 60 °C | 5.4 | >50%; 40 °C; 2 h >45%; 50 °C; 2 h 25%; 55 °C; 1 h | Fungi (Neurospora crassa) | [172] |
| Bgl1 | 3 | 50 °C | 3.5–5.0 | 100%; 45 °C; 30 min | Fungi (Mucor circinelloides) | [173] |
| Bgl2 | 3 | 55 °C | 3.5–5.5 | 100%; 55 °C; 30 min | Fungi (Mucor circinelloides) | [173] |
| NfBGL1 | 3 | 80 °C | 5.0 | >80%; 70 °C; 2 h | Fungi (Neosartorya fischeri) | [174] |
| PtBglu3 | 3 | 65 °C | 6.0 | >85%; 60 °C; 30 min | Fungi (Paecilomyces thermophila) | [32] |
| Bgl1 | 3 | 70 °C | 4.8 | 50%; 65 °C; 24 h | Fungi (Penicillium brasilianum) | [175] |
| pBGL1 | 3 | 65–70 °C | 4.5–5.50 | 96.3%; 50 °C; 12 h 50%; 70 °C; 4 h | Fungi (Penicillium decumbens) | [176] |
| Bgl1 | 3 | 70 °C | 5.0–6.0 | 60%; 70 °C; 1.5 h | Fungi (Periconia sp.) | [43] |
| RmBglu3B | 3 | 50 °C | 5.0 | 50%; 50 °C; 30 min | Fungi (Rhizomucor miehei) | [50] |
| Bgl1 | 3 | 50 °C | 5.0 | >70%; 50 °C; 30 min <10%; 60 °C; 30 min | Fungi (Saccharomycopsis fibuligera) | [177] |
| Bgl2 | 3 | 50 °C | 5.0 | >70%; 50 °C; 30 min <10%; 60 °C; 30 min | Fungi (Saccharomycopsis fibuligera) | [177] |
| β-glucosidase | 3 | 75 °C | 4.5 | 50%; 60 °C; 136 h 50%; 65 °C; 55 h 50%; 70 °C; 10 h 50%; 75 °C; 1 h | Fungi (Talaromyces aculeatus) | [53] |
| Cel3a | 3 | 71.5 °C | 4.02 | 50%; 65 °C; 62 min 50%; 75 °C; 18 min | Fungi (Talaromyces emersonii) | [178] |
| Bgl3A | 3 | 75 °C | 4.5 | >65%; 60 °C; 1 h | Fungi (Talaromyces leycettanus) | [179] |
| Bgl1 | 3 | 70 °C | 5.0 | >70%; 60 °C; 1 h | Fungi (Thermoascus auranticus) | [180] |
| Bgl3a | 3 | 70 °C | 5.0 | 50%; 60 °C; 143 min | Fungi (Myceliophthora thermophila) | [181] |
| RG3 | 3 | 50–55 °C | 5.5–6.0 | NM | Uncultured organism (Rabbit cecum metagenome) | [67] |
| RG14 | 3 | 50–55 °C | 5.5–7.0 | NM | Uncultured organism (Rabbit cecum metagenome) | [67] |
| BGL7 | 3 | 50 °C | 6.5 | NM | Uncultured organism (Termite gut metagenome) | [72] |
| LAB25g2 | 3 | 55 °C | 4.5 | 82%; 50 °C; 5 d | Uncultured organism (Cow rumen metagenome) | [68] |
| SRF2g14 | 3 | 55 °C | 5.0 | 50%; 50 °C; 18.06 h | Uncultured organism (Cow rumen metagenome) | [68] |
| SRF2g18 | 3 | 50 °C | 4.0 | 50%; 50 °C; 37.5 h | Uncultured organism (Cow rumen metagenome) | [68] |
| RuBGX1 | 3 | 50 °C | 6.0 | 62%; 50 °C; 10 min | Uncultured organism (Yak rumen metagenome) | [36] |
| JMB19063 | 3 | 50–55 °C | 6.5 | NM | Uncultured organism (Compost metagenome) | [37] |
| GlyA1 | 3 | 55 °C | 6.5 | NM | Uncultured organism (Cow rumen metagenome) | [69] |
| Bgx1 | 30 | 50 °C | 4.0–6.0 | NM | Oomycota (Phytophthora infestans) | [182] |
| SSO3039 | 116 | >70 °C | 4.0 | >70%; 65 °C; 48 h >50%; 85 °C; 8 h | Arquea (Sulfolobus solfataricus) | [183] |
| TxGH116 | 116 | 85 °C | 6.0 | NM | Bacteria (Thermoanaerobacterium xylanolyticum) | [184] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escuder-Rodríguez, J.-J.; DeCastro, M.-E.; Cerdán, M.-E.; Rodríguez-Belmonte, E.; Becerra, M.; González-Siso, M.-I. Cellulases from Thermophiles Found by Metagenomics. Microorganisms 2018, 6, 66. https://doi.org/10.3390/microorganisms6030066
Escuder-Rodríguez J-J, DeCastro M-E, Cerdán M-E, Rodríguez-Belmonte E, Becerra M, González-Siso M-I. Cellulases from Thermophiles Found by Metagenomics. Microorganisms. 2018; 6(3):66. https://doi.org/10.3390/microorganisms6030066
Chicago/Turabian StyleEscuder-Rodríguez, Juan-José, María-Eugenia DeCastro, María-Esperanza Cerdán, Esther Rodríguez-Belmonte, Manuel Becerra, and María-Isabel González-Siso. 2018. "Cellulases from Thermophiles Found by Metagenomics" Microorganisms 6, no. 3: 66. https://doi.org/10.3390/microorganisms6030066