
Fed-Batch Production of Bacterial Ghosts Using Dielectric Spectroscopy for Dynamic Process Control

 and
and
Abstract
:1. Introduction
2. Methods
2.1. Strain
2.2. Media
2.3. Plasmid Stability
2.4. Bioreactors
2.5. Fermentation
2.6. Off-Gas Analytics
2.7. Dielectric Spectroscopy
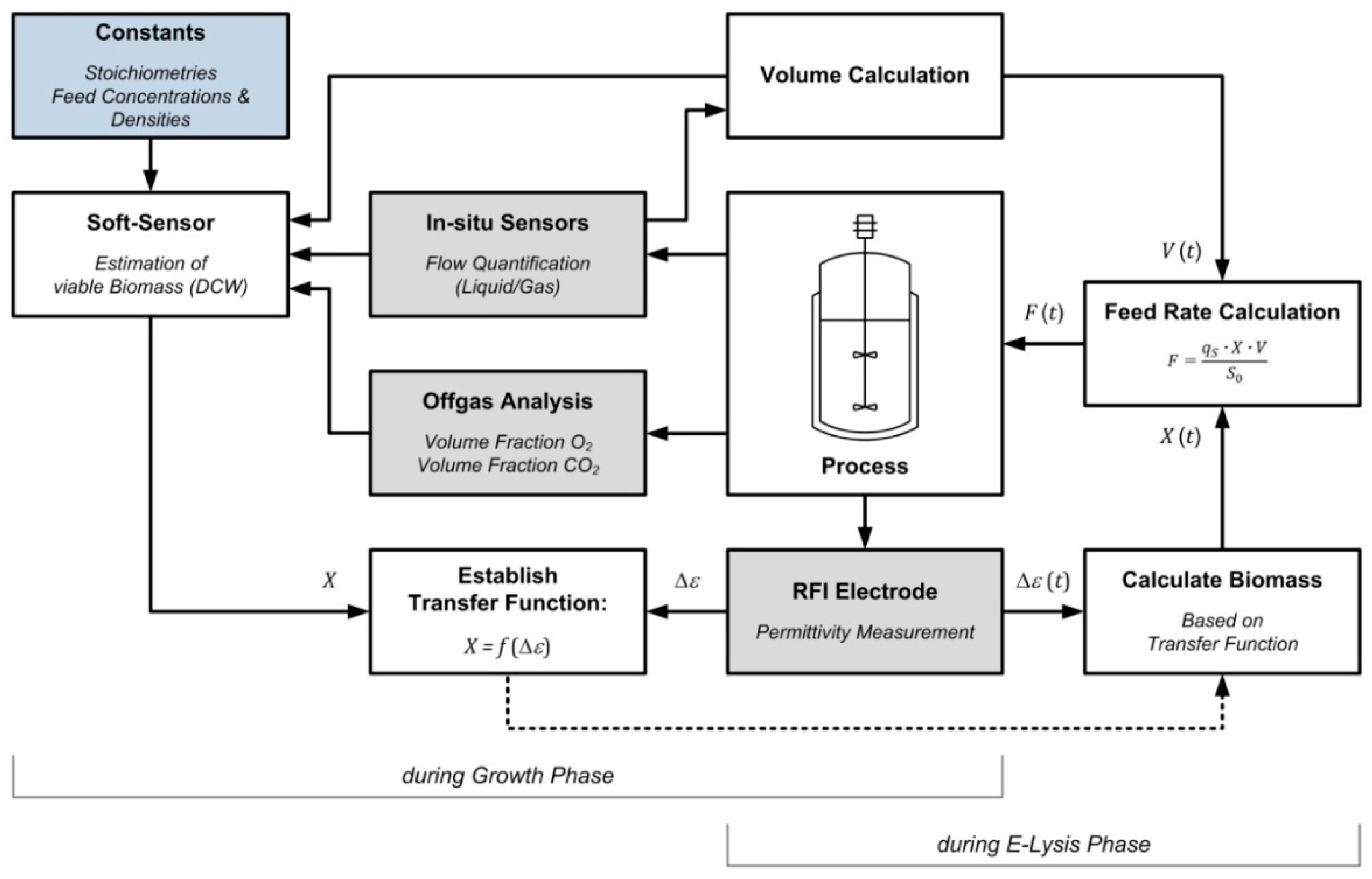
2.8. Process Management and Soft-Sensor
2.9. Biomass Concentrations
2.10. Flow Cytometry
2.11. E-Lysis Efficiency
3. Results
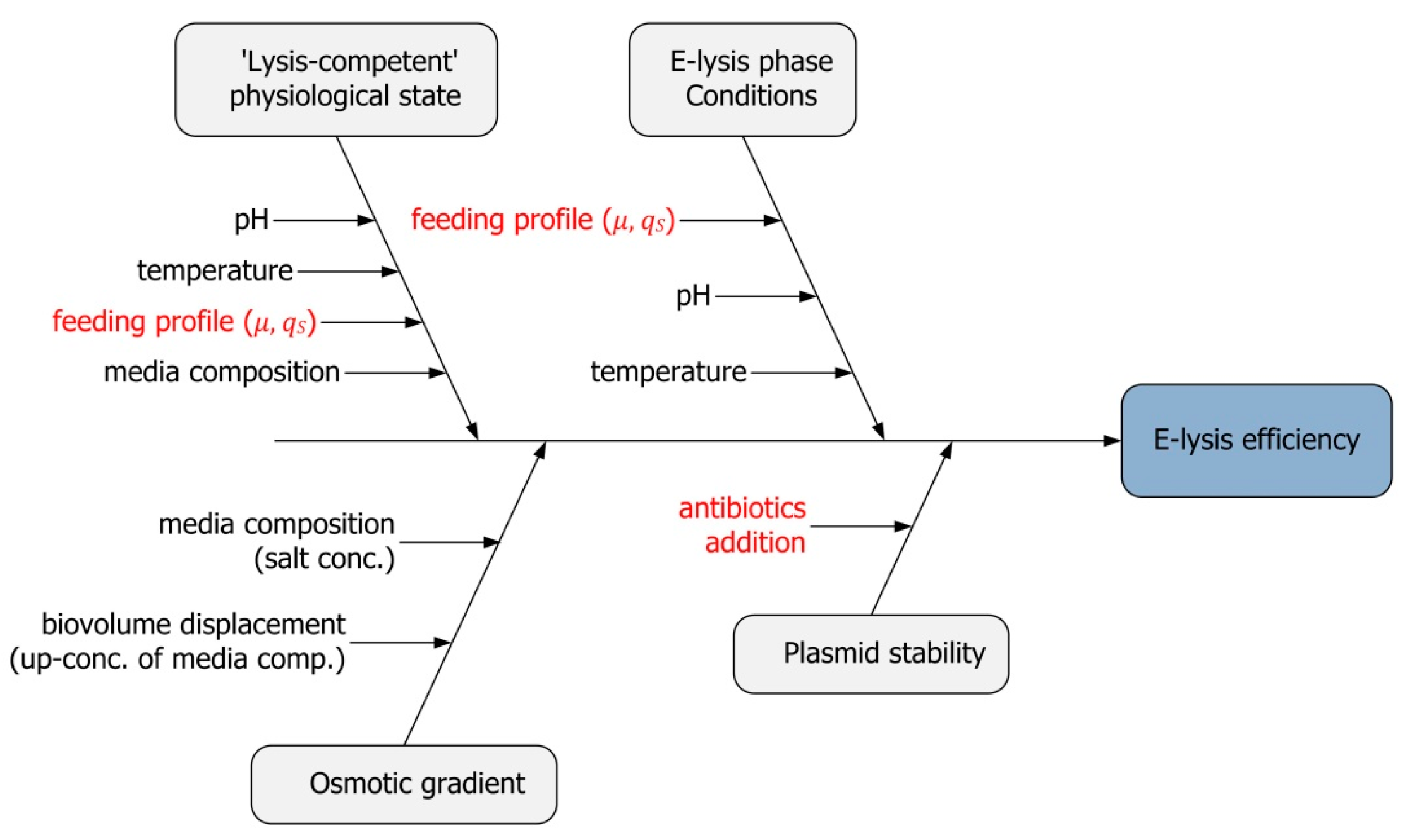
3.1. Identification of Critical Process Parameters
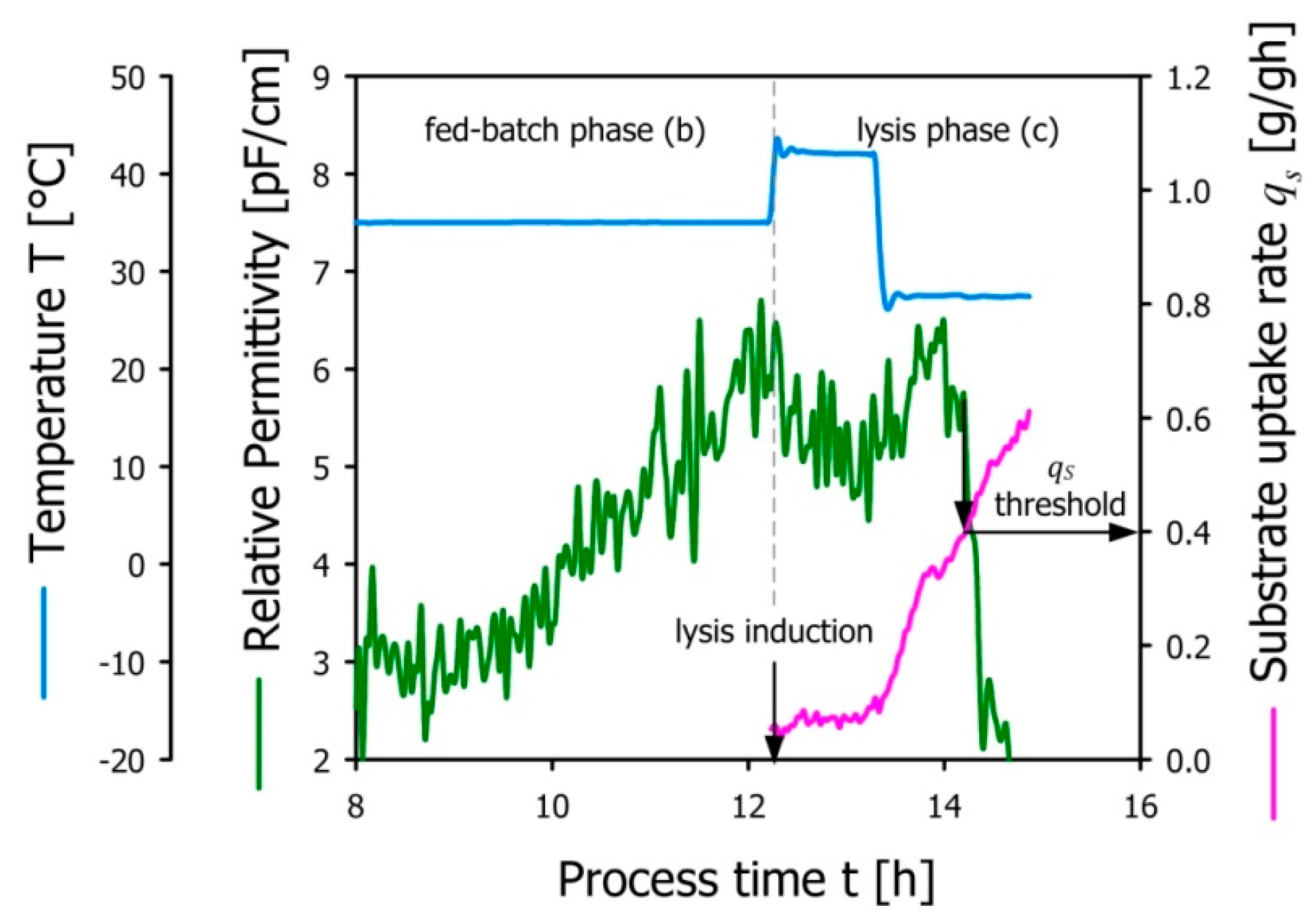
3.2. Effect of Specific Substrate Uptake Rate during Fed-Batch Phase on Lysis Efficiency
3.3. Determination of E-lysis Conditions Using Dielectric Spectroscopy
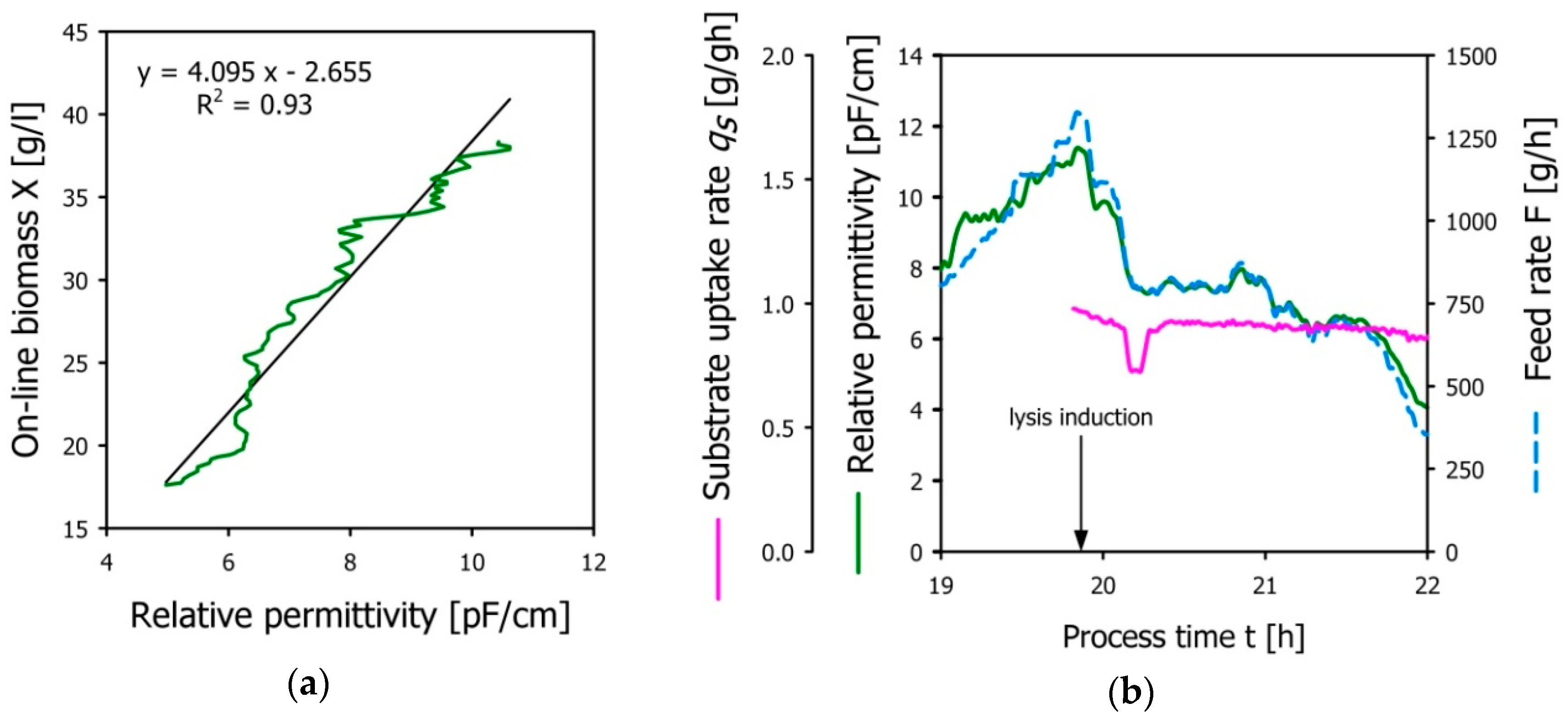
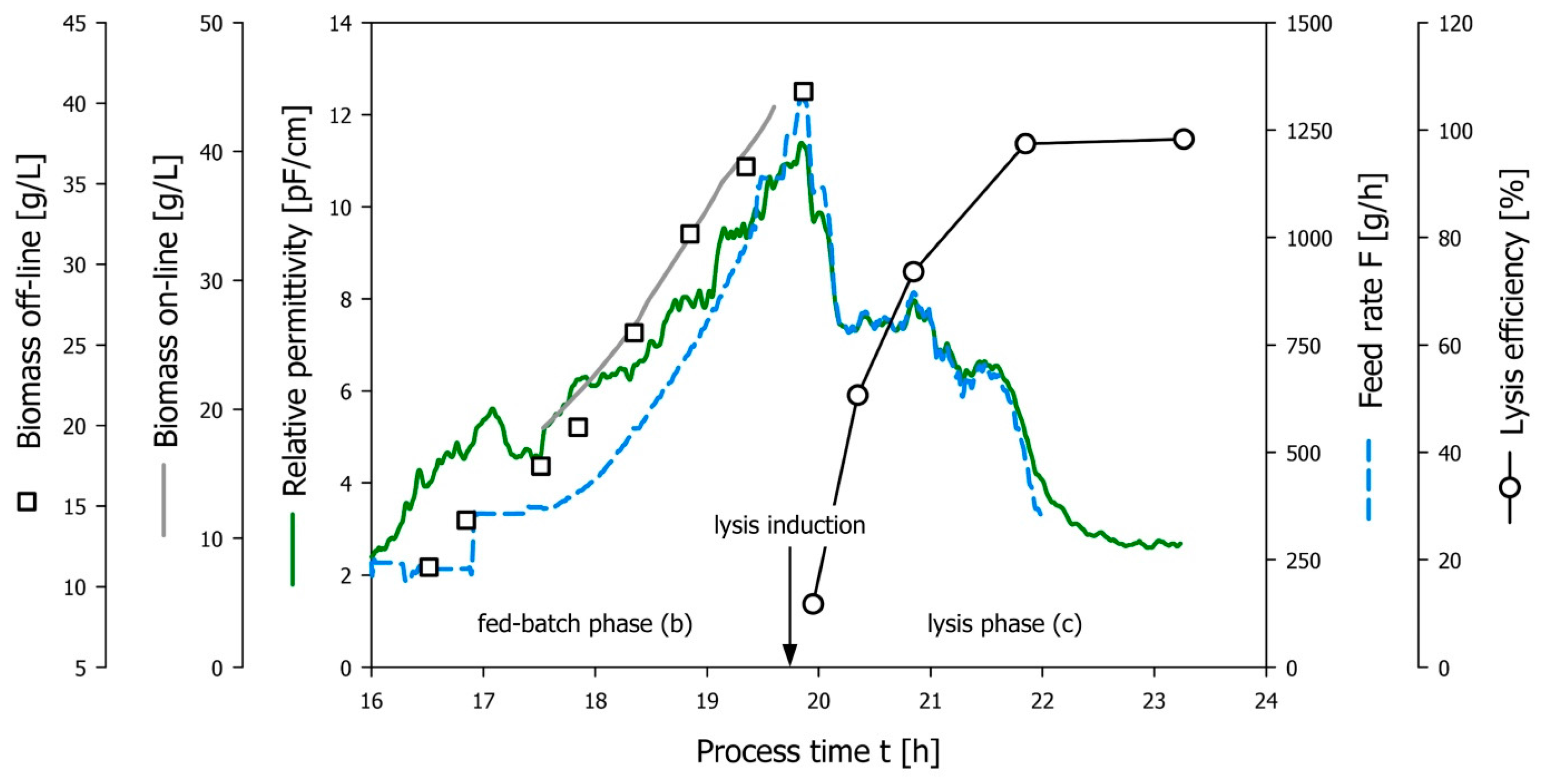
3.4. Demonstration of the Developed Permittivity Controlled Feeding Strategy
4. Discussion
5. Conclusions
- The biological process of E-lysis depends on a certain level of physiological activity; using the specific substrate uptake rate as a biomass specific measure of total metabolism, we were able to determine a threshold value qS > 0.4 g/gh required for successful E-lysis under substrate-limited conditions.
- We could show that quantifying the relative permittivity of the culture broth as a measure for membrane polarizability, one can follow the drop of viability in a culture affected by E-lysis.
- Viability information gained from permittivity measurements (Δε) could be aligned with soft-sensor-derived estimation of viable biomass concentrations (X) during bacterial growth.
- Using a transfer function X = f (Δε), we were able to control the specific substrate uptake rate qS throughout the entire lysis phase.
- The described fed-batch process increased the productivity of Bacterial Ghost production by a factor of 8–10 reaching lysis efficiencies of >98%.
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| µ | specific growth rate |
| µmax | maximum specific growth rate |
| Δε | relative permittivity |
| BGs | Bacterial Ghosts |
| DCW | dry cell weight |
| DoE | design of experiments |
| DoR | Degree of Reduction |
| EcN | Escherichia coli Nissle 1917 |
| E-lysis | protein E-mediated lysis |
| F | feed flow rate |
| FCM | flow cytometry |
| FDA | Food and Drug Administration |
| LE | lysis efficiency |
| LI | E-lysis induction |
| MFC | mass flow controller |
| PAT | process analytical technology |
| PCS | process control system |
| PID controller | proportional-integral-derivative controller |
| qS | specific substrate uptake rate |
| qS,max | maximum specific substrate uptake rate |
| V | (fermentation) volume |
| X | viable biomass concentration |
| YX/S | biomass yield coefficient |
References
- Bläsi, U.; Henrich, B.; Lubitz, W. Lysis of Escherichia coli by cloned phi X174 gene E depends on its expression. J. Gen. Microbiol. 1985, 131, 1107–1114. [Google Scholar] [PubMed]
- Schön, P.; Schrot, G.; Wanner, G.; Lubitz, W.; Witte, A. Two-stage model for integration of the lysis protein E of phi X174 into the cell envelope of Escherichia coli. FEMS Microbiol. Rev. 1995, 17, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Witte, A.; Wanner, G.; Bläsi, U.; Halfmann, G.; Szostak, M.; Lubitz, W. Endogenous transmembrane tunnel formation mediated by phi X174 lysis protein E. J. Bacteriol. 1990, 172, 4109–4114. [Google Scholar] [PubMed]
- Witte, A.; Wanner, G.; Sulzner, M.; Lubitz, W. Dynamics of PhiX174 protein e-mediated lysis of Escherichia coli. Arch. Microbiol. 1992, 157, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, G.; Leduc, M.; Lubitz, W. Different sensitivity of autolytic deficient Escherichia coli mutants to the mode of induction. FEMS Microbiol. Lett. 1984, 24, 205–208. [Google Scholar] [CrossRef]
- Witte, A.; Lubitz, W. Biochemical characterization of Phi X174-protein-E-mediated lysis of Escherichia coli. Eur. J. Biochem. 1989, 180, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, G.; Lubitz, W. Differential induction of Escherichia coli autolysis by penicillin and the bacteriophage Phi X174 gene E product. J. Bacteriol. 1986, 166, 683–685. [Google Scholar] [PubMed]
- Lubitz, W.; Pugsley, A.P. Changes in host cell phospholipid composition of øX174 gene E product. FEMS Microbiol. Lett. 1985, 30, 171–175. [Google Scholar] [CrossRef]
- Langemann, T.; Koller Verena, J.; Muhammad, A.; Kudela, P.; Mayr Ulrike, B.; Lubitz, W. The bacterial ghost platform system: Production and applications. Bioeng. Bugs 2010, 1, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Langemann, T.; Mayr, U.; Meitz, A.; Lubitz, W.; Herwig, C. Multi-parameter flow cytometry as a process analytical technology (PAT) approach for the assessment of bacterial ghost production. Appl. Microbiol. Biotechnol. 2016, 100, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Cannizzaro, C.; Gugerli, R.; Marison, I.; von Stockar, U. On-line biomass monitoring of CHO perfusion culture with scanning dielectric spectroscopy. Biotechnol. Bioeng. 2003, 84, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Carvell, J.P.; Dowd, J.E. On-line measurements and control of viable cell density in cell culture manufacturing processes using radio-frequency impedance. Cytotechnology 2006, 50, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Parta, L.; Zalai, D.; Borbely, S.; Putics, A. Application of dielectric spectroscopy for monitoring high cell density in monoclonal antibody producing CHO cell cultivations. Bioprocess Biosyst. Eng. 2014, 37, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Noll, T.; Biselli, M. Dielectric spectroscopy in the cultivation of suspended and immobilized hybridoma cells. J. Biotechnol. 1998, 63, 187–198. [Google Scholar] [CrossRef]
- Xiong, Z.-Q.; Guo, M.-J.; Guo, Y.-X.; Chu, J.; Zhuang, Y.-P.; Zhang, S.-L. Real-time viable-cell mass monitoring in high-cell-density fed-batch glutathione fermentation by saccharomyces cerevisiae T65 in industrial complex medium. J. Biosci. Bioeng. 2008, 105, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.P.; Vieira, L.M.; Cardoso, J.P.; Menezes, J.C. Evaluation of a new annular capacitance probe for biomass monitoring in industrial pilot-scale fermentations. J. Biotechnol. 2005, 116, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Maskow, T.; Rollich, A.; Fetzer, I.; Ackermann, J.U.; Harms, H. On-line monitoring of lipid storage in yeasts using impedance spectroscopy. J. Biotechnol. 2008, 135, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Arnoux, A.S.; Preziosi-Belloy, L.; Esteban, G.; Teissier, P.; Ghommidh, C. Lactic acid bacteria biomass monitoring in highly conductive media by permittivity measurements. Biotechnol. Lett. 2005, 27, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, C.; Carvell, J.P.; Luttmann, R. A sensitive, compact, in situ biomass measurement system controlling and monitoring microbial fermentations using radio-frequency impedance. Bioprocess Int. 2007, 5, 52–55. [Google Scholar]
- Harris, C.M.; Todd, R.W.; Bungard, S.J.; Lovitt, R.W.; Morris, J.G.; Kell, D.B. Dielectric permittivity of microbial suspensions at radio frequencies: A novel method for the real-time estimation of microbial biomass. Enzyme Microb. Technol. 1987, 9, 181–186. [Google Scholar] [CrossRef]
- Markx, G.H.; Davey, C.L. The dielectric properties of biological cells at radiofrequencies: Applications in biotechnology. Enzyme Microb. Technol. 1999, 25, 161–171. [Google Scholar] [CrossRef]
- Asami, K.; Yonezawa, T.; Wakamatsu, H.; Koyanagi, N. Dielectric spectroscopy of biological cells. Bioelectrochem. Bioenerg. 1996, 40, 141–145. [Google Scholar] [CrossRef]
- Davey, C.L.; Davey, H.M.; Kell, D.B.; Todd, R.W. Introduction to the dielectric estimation of cellular biomass in real time, with special emphasis on measurements at high volume fractions. Anal. Chim. Acta 1993, 279, 155–161. [Google Scholar] [CrossRef]
- Kiviharju, K.; Salonen, K.; Moilanen, U.; Eerikainen, T. Biomass measurement online: The performance of in situ measurements and software sensors. J. Ind. Microbiol. Biotechnol. 2008, 35, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Ehgartner, D.; Sagmeister, P.; Herwig, C.; Wechselberger, P. A novel real-time method to estimate volumetric mass biodensity based on the combination of dielectric spectroscopy and soft-sensors. J. Chem. Technol. Biotechnol. 2015, 90, 262–272. [Google Scholar] [CrossRef]
- Sührer, I.; Langemann, T.; Lubitz, W.; Weuster-Botz, D.; Castiglione, K. A novel one-step expression and immobilization method for the production of biocatalytic preparations. Microb. Cell Fact. 2015. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, B.; Locher, G.; Fiechter, A. Biomass determination. J. Biotechnol. 1992, 25, 5–22. [Google Scholar] [CrossRef]
- Luttmann, R.; Bracewell, D.G.; Cornelissen, G.; Gernaey, K.V.; Glassey, J.; Hass, V.C.; Kaiser, C.; Preusse, C.; Striedner, G.; Mandenius, C.-F. Soft sensors in bioprocessing: A status report and recommendations. Biotechnol. J. 2012, 7, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Gadkar, K.G.; Mehra, S.; Gomes, J. On-line adaptation of neural networks for bioprocess control. Comput. Chem. Eng. 2005, 29, 1047–1057. [Google Scholar] [CrossRef]
- Nayak, R.; Gomes, J. Sequential adaptive networks: An ensemble of neural networks for feed forward control of l-methionine production. Chem. Eng. Sci. 2009, 64, 2401–2412. [Google Scholar] [CrossRef]
- Van der Heijden, R.T.; Heijnen, J.J.; Hellinga, C.; Romein, B.; Luyben, K.C. Linear constraint relations in biochemical reaction systems: I. Classification of the calculability and the balanceability of conversion rates. Biotechnol. Bioeng. 1994, 43, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Wechselberger, P.; Sagmeister, P.; Herwig, C. Real-time estimation of biomass and specific growth rate in physiologically variable recombinant fed-batch processes. Bioprocess Biosyst. Eng. 2013, 36, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Sagmeister, P.; Kment, M.; Wechselberger, P.; Meitz, A.; Langemann, T.; Herwig, C. Soft-sensor assisted dynamic investigation of mixed feed bioprocesses. Process Biochem. 2013, 48, 1839–1847. [Google Scholar] [CrossRef]
- Sagmeister, P.; Langemann, T.; Wechselberger, P.; Meitz, A.; Herwig, C. A dynamic method for the investigation of induced state metabolic capacities as a function of temperature. Microb. Cell Fact. 2013, 12, 94–94. [Google Scholar] [CrossRef] [PubMed]
- Sagmeister, P.; Wechselberger, P.; Jazini, M.; Meitz, A.; Langemann, T.; Herwig, C. Soft sensor assisted dynamic bioprocess control: Efficient tools for bioprocess development. Chem. Eng. Sci. 2013, 96, 190–198. [Google Scholar] [CrossRef]
- ICH Guidelines—Pharmaceutical Development Q8 (R2). Available online: http://www.ich.org/fileadmin/Public_Web_Site/ICH_Products/Guidelines/Quality/Q8_R1/Step4/Q8_R2_Guideline.pdf (accessed on 7 March 2012).
- Mab: A Case Study in Bioprocess Development. Available online: http://c.ymcdn.com/sites/www.casss.org/resource/resmgr/imported/A-Mab_Case_Study_Version_2-1.pdf (accessed on 23 March 2012).
- Rathore, A.S. Roadmap for implementation of quality by design (QbD) for biotechnology products. Trends Biotechnol. 2009, 27, 546–553. [Google Scholar] [CrossRef] [PubMed]
- PAT-A Framework for Innovative Pharmaceutical Development, Manufacturing, and Quality Assurance. Available online: http://www.fda.gov/downloads/Drugs/Guidances/ucm070305.pdf (accessed on 13 July 2013).
- Mayr, U.B.; Koller, V.J.; Lubitz, P.; Lubitz, W. Bacterial Ghosts as Vaccine and Drug Delivery Platforms; Landes Bioscience: Austin, TX, USA, 2008; pp. 50–59. [Google Scholar]
- Kudela, P.; Koller, V.J.; Lubitz, W. Bacterial ghosts (BGs)—Advanced antigen and drug delivery system. Vaccine 2010, 28, 5760–5767. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, P.; Mayr, U.B.; Lubitz, W. Applications of bacterial ghosts in biomedicine. Adv. Exp. Med. Biol. 2009, 655, 159–170. [Google Scholar] [PubMed]
- Mayr, U.B.; Walcher, P.; Azimpour, C.; Riedmann, E.; Haller, C.; Lubitz, W. Bacterial ghosts as antigen delivery vehicles. Adv. Drug Deliv. Rev. 2005, 57, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Champeimont, J.; Mayr, U.B.; Lubitz, W.; Kudela, P. Bacterial ghosts as carriers of protein subunit and DNA-encoded antigens for vaccine applications. Expert Rev. Vaccines 2012, 11, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Riedmann, E.M.; Kyd, J.M.; Cripps, A.W.; Lubitz, W. Bacterial ghosts as adjuvant particles. Expert Rev. Vaccines 2007, 6, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Vilte, D.A.; Larzábal, M.; Mayr, U.B.; Garbaccio, S.; Gammella, M.; Rabinovitz, B.C.; Delgado, F.; Meikle, V.; Cantet, R.J.C.; Lubitz, P.; et al. A systemic vaccine based on Escherichia coli O157:H7 bacterial ghosts (BGs) reduces the excretion of E. coli O157:H7 in calves. Vet. Immunol. Immunopathol. 2012, 146, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kudela, P.; Koller, V.J.; Mayr, U.B.; Nepp, J.; Lubitz, W.; Barisani-Asenbauer, T. Bacterial ghosts as antigen and drug delivery system for ocular surface diseases: Effective internalization of bacterial ghosts by human conjunctival epithelial cells. J. Biotechnol. 2011, 153, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Abtin, A.; Kudela, P.; Mayr, U.B.; Koller, V.J.; Mildner, M.; Tschachler, E.; Lubitz, W. Escherichia coli ghosts promote innate immune responses in human keratinocytes. Biochem. Biophys. Res. Commun. 2010, 400, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Chen, A.Y.; Wang, H.; McAllister, L.J.; Höggerl, F.; Mayr, U.B.; Shewell, L.K.; Jennings, M.P.; Morona, R.; Lubitz, W.; et al. Protection against shiga-toxigenic Escherichia coli by non-genetically modified organism receptor mimic bacterial ghosts. Infect. Immun. 2015, 83, 3526–3533. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.; McKeithen, D.; Martinez, O.; Igietseme, J.; Eko, F.; Black, C.; He, Q.; Ananaba, G. Vibrio cholerae ghosts elicit the production of soluble immunostimulatory factors and prevent development of chlamydia infection (INM3P.408). J. Immunol. 2015, 194, 127–133. [Google Scholar]
- DeLisa, M.P.; Li, J.; Rao, G.; Weigand, W.A.; Bentley, W.E. Monitoring GFP-operon fusion protein expression during high cell density cultivation of Escherichia coli using an on-line optical sensor. Biotechnol. Bioeng. 1999, 65, 54–64. [Google Scholar] [CrossRef]
- Langemann, T. Process Development for Industrial Scale Bacterial Ghost Production. Ph.D. Thesis, University of Vienna, Vienna, Austria, 2011. [Google Scholar]
- Witte, A.; Lubitz, W.; Bakker, E.P. Proton-motive-force-dependent step in the pathway to lysis of Escherichia coli induced by bacteriophage phi X174 gene E product. J. Bacteriol. 1987, 169, 1750–1752. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process Parameter | Value | |
|---|---|---|
| General | growth temperature [°C] | 35 |
| pressure [bar] | 1.2/- * | |
| air flow rate [L/min] | 1 vvm | |
| stirring speed [rpm] | 1200/1500 * | |
| Batch (a) | glucose conc. (S) [g/L] | 20.0 |
| final biomass X0 [g/L] | 9.0 | |
| volume V0 [L] | 7.0/1.0 * | |
| Fed–Batch (b) | specific substrate uptake rate qS [g/gh] | 0.6/varying * |
| oxygen flow rate [L/min] | max. 0.1/1.0 vvm * | |
| glucose conc. S0 feed [g/L] | 400 | |
| target biomass concentration X [g/L] | 40 |
| qS (b) [g/gh] | R1 [counts/mL] | R2 [counts/mL] | R3 [counts/mL] | LE [%] |
|---|---|---|---|---|
| 0.4 | 1.04 × 109 | n.d. | 2.20 × 1010 | 95.5 |
| 0.7 | 2.53 × 109 | 1.20 × 106 | 2.82 × 1010 | 91.7 |
| 0.7 | 1.30 × 109 | 5.20 × 106 | 2.25 × 1010 | 94.5 |
| 1.0 | 3.82 × 108 | n.d. | 1.13 × 1010 | 96.7 |
© 2016 by the authors; Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meitz, A.; Sagmeister, P.; Lubitz, W.; Herwig, C.; Langemann, T. Fed-Batch Production of Bacterial Ghosts Using Dielectric Spectroscopy for Dynamic Process Control. Microorganisms 2016, 4, 18. https://doi.org/10.3390/microorganisms4020018
Meitz A, Sagmeister P, Lubitz W, Herwig C, Langemann T. Fed-Batch Production of Bacterial Ghosts Using Dielectric Spectroscopy for Dynamic Process Control. Microorganisms. 2016; 4(2):18. https://doi.org/10.3390/microorganisms4020018
Chicago/Turabian StyleMeitz, Andrea, Patrick Sagmeister, Werner Lubitz, Christoph Herwig, and Timo Langemann. 2016. "Fed-Batch Production of Bacterial Ghosts Using Dielectric Spectroscopy for Dynamic Process Control" Microorganisms 4, no. 2: 18. https://doi.org/10.3390/microorganisms4020018