
Influence of pH Regulation Mode in Glucose Fermentation on Product Selection and Process Stability

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Methods
2.1. Inoculum
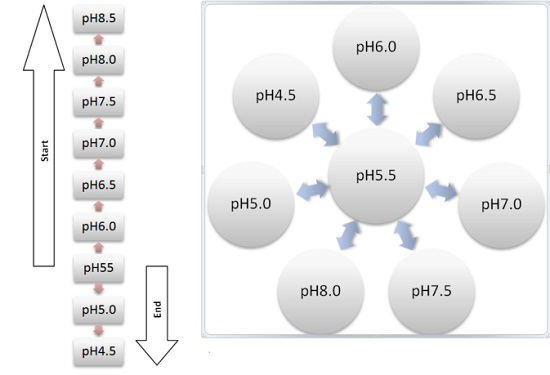
2.2. Reactor Setup

2.3. Reactor Operation
2.4. Chemical Analysis
2.5. Data Processing and Statistical Analysis
2.6. Microbial Analysis
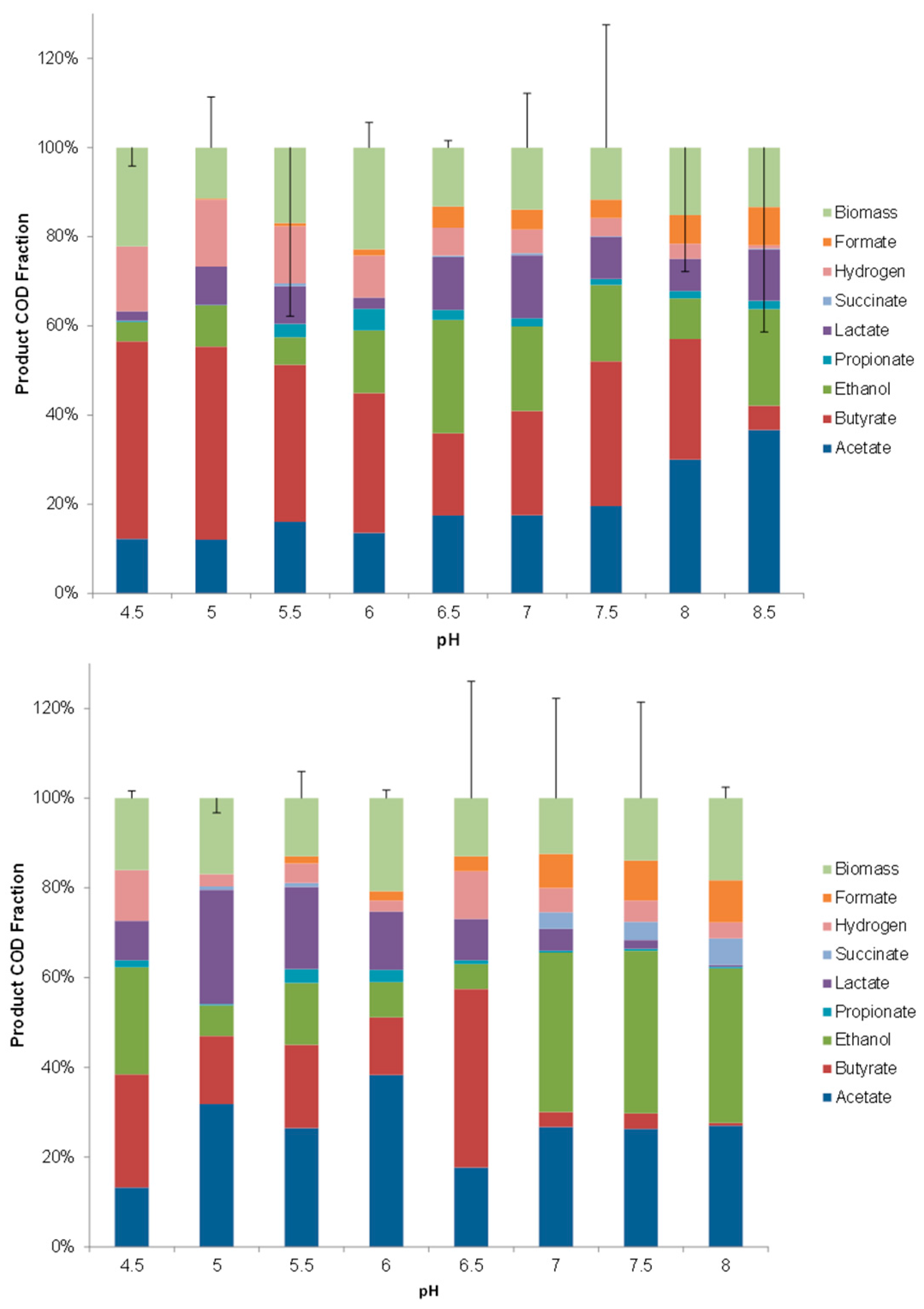
3. Results
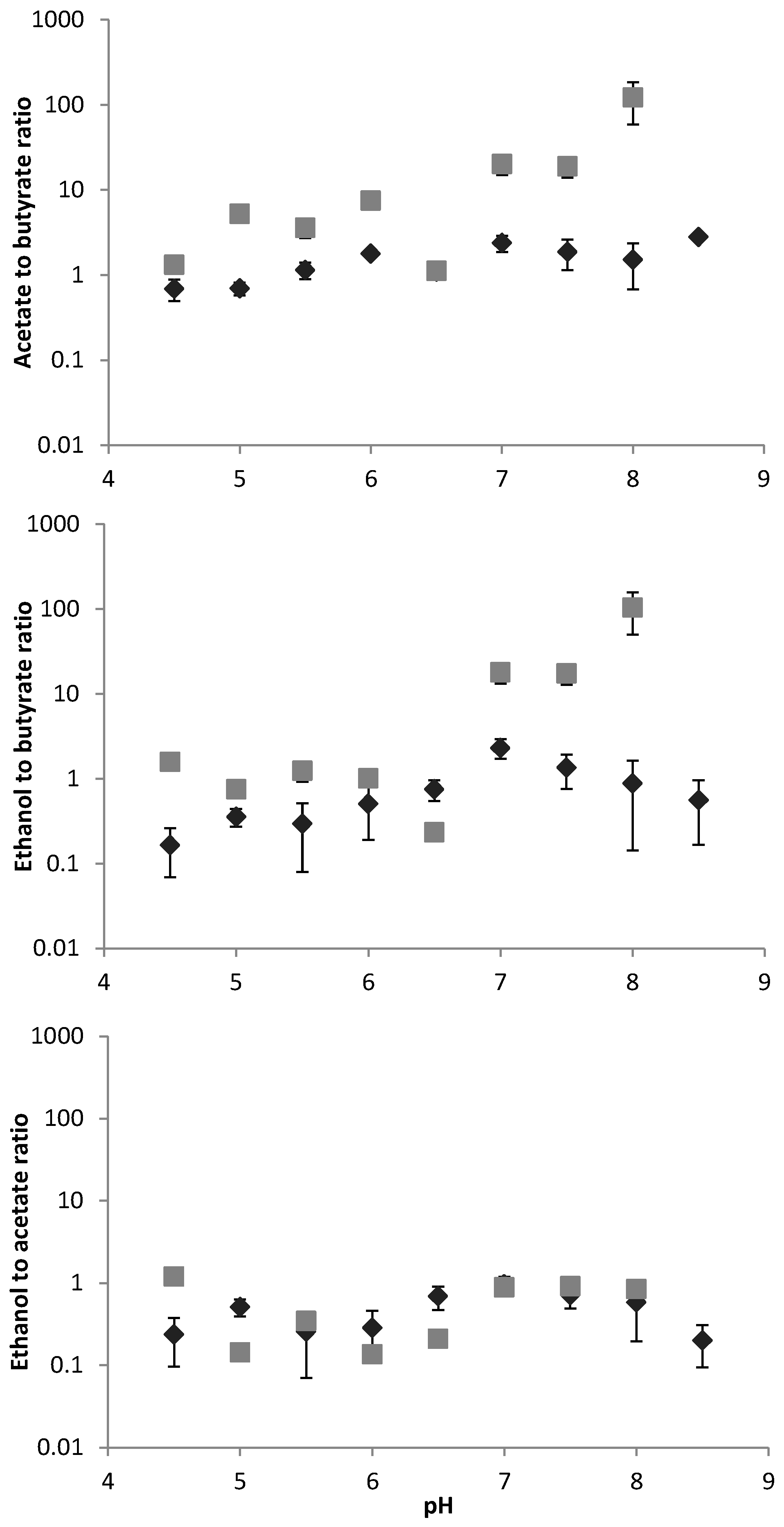
3.1. Major VFA Products


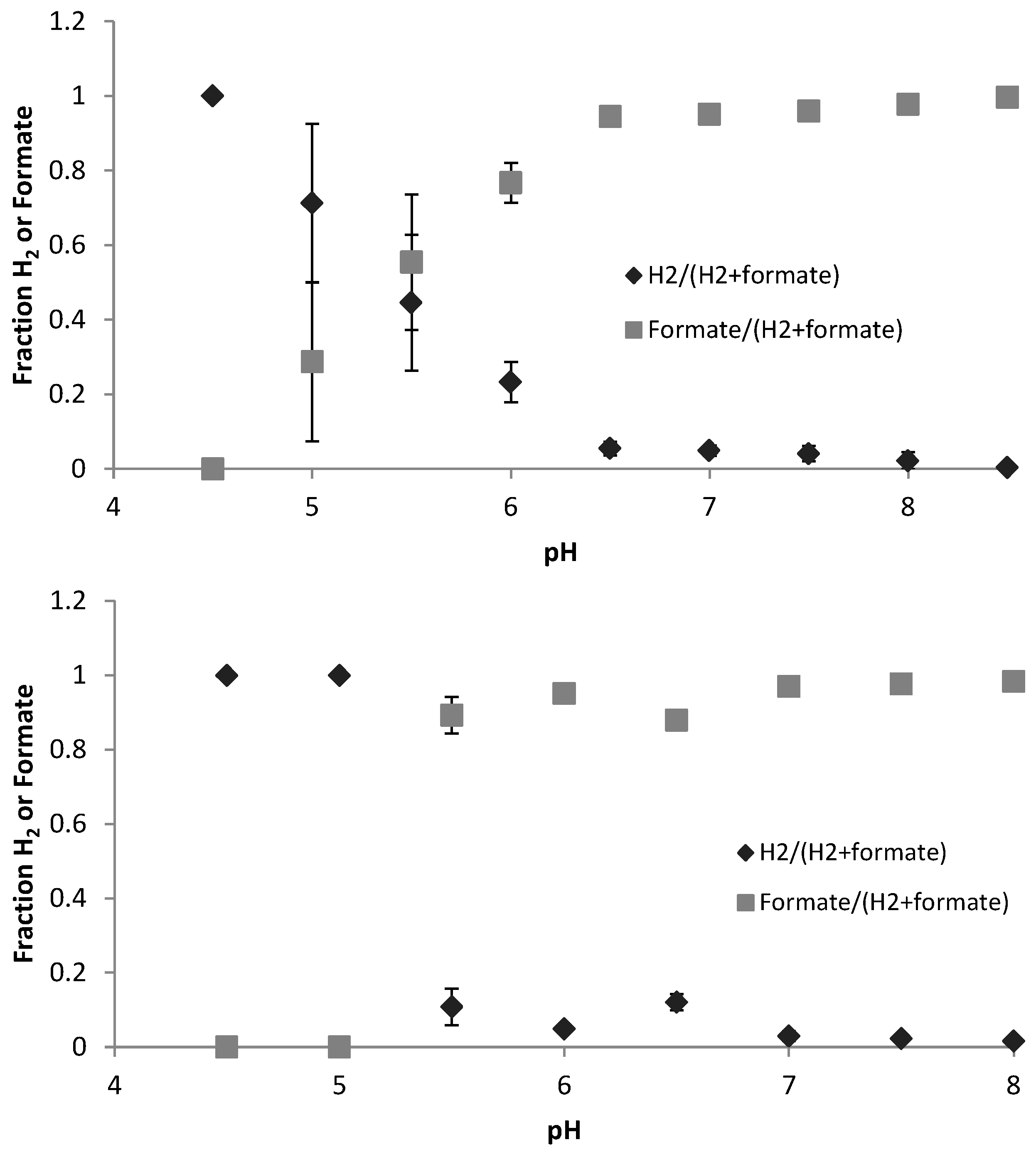
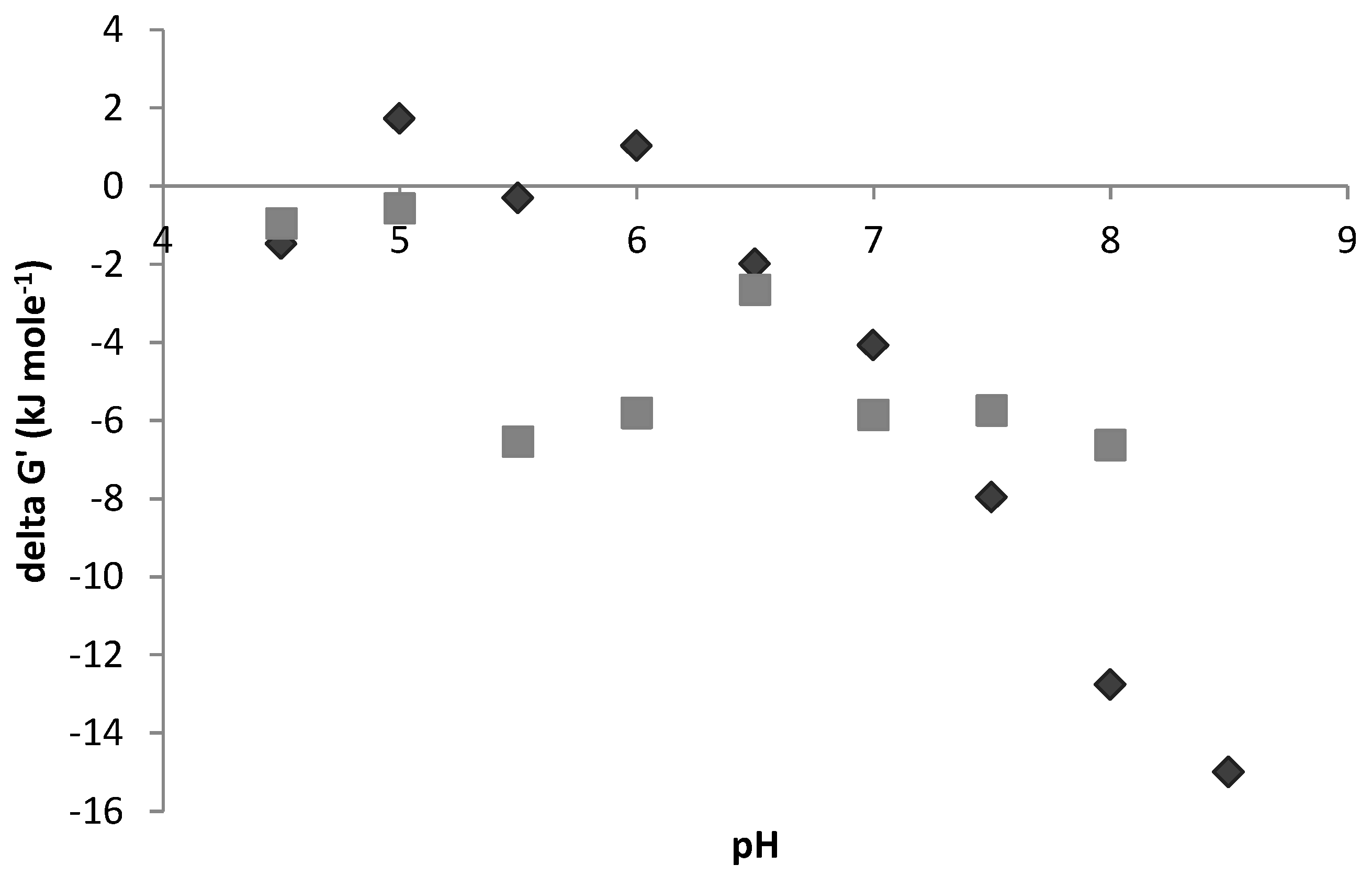
3.2. Hydrogen and Formate (Electron Sinks)
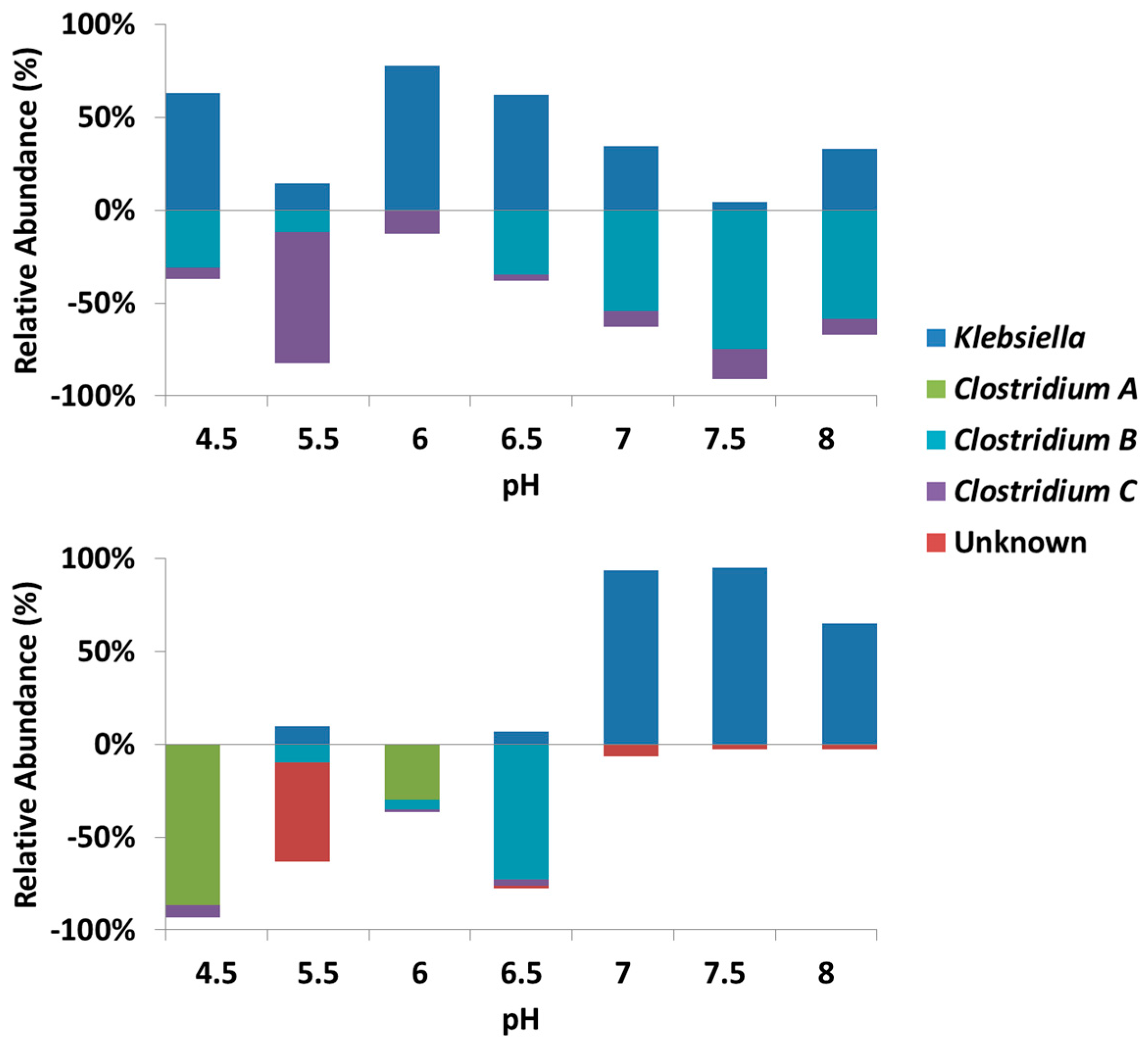
3.3. Microbial Community


4. Discussion

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Batstone, D.J.; Virdis, B. The role of anaerobic digestion in the emerging energy economy. Curr. Opin. Biotechnol. 2014, 27, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, R.; van Loosdrecht, M.C.M. Mixed culture biotechnology for bioenergy production. Curr. Opin. Biotechnol. 2007, 18, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Fang, H.H.P. Fermentative hydrogen production from wastewater and solid wastes by mixed culture. Crit. Rev. Environ. Sci. Technol. 2007, 37, 1–39. [Google Scholar] [CrossRef]
- Bastidas-Oyanedel, J.R.; Bonk, F.; Thomsen, M.H.; Schmidt, J.E. Dark fermentation biorefinery in the present and future (bio)chemical industry. Rev. Environ. Sci. Biotechnol. 2015, 14, 473–498. [Google Scholar] [CrossRef]
- Temudo, M.F.; Kleerebezem, R.; van Loosdrecht, M.C.M. Influence of the pH on (open) mixed culture fermentation of glucose: A chemostat study. Biotechnol. Bioeng. 2007, 98, 69–79. [Google Scholar] [CrossRef] [PubMed]
- González-Cabaleiro, R.; Lema, J.M.; Rodríguez, J. Metabolic energy-based modelling explains product yielding in anaerobic mixed culture fermentations. PLoS ONE 2015, 10, e0126739. [Google Scholar] [CrossRef] [PubMed]
- Hoelzle, R.D.; Virdis, B.; Batstone, D.J. Regulation mechanisms in mixed and pure culture microbial fermentation. Biotechnol. Bioeng. 2014, 111, 2139–2154. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, J.I.; Shimizu, T.; Tada, K.; Kanno, T.; Kobayashi, M. Selective production of organic acids in anaerobic acid reactor by pH control. Bioresour. Technol. 2002, 82, 209–213. [Google Scholar] [CrossRef]
- Madigan, M.T.; Martinko, J.M.; Parker, J.; Brock, T.D. Biology of Microorganisms, 13th ed.; Pearson Education, Inc.: New York, NY, USA, 2012. [Google Scholar]
- Ren, N.; Wang, B.; Huang, J.C. Ethanol-type fermentation from carbohydrate in high rate acidogenic reactor. Biotechnol. Bioeng. 1997, 54, 428–433. [Google Scholar] [CrossRef]
- Fang, H.H.; Liu, H. Effect of pH on hydrogen production from glucose by a mixed culture. Bioresour. Technol. 2002, 82, 87–93. [Google Scholar] [CrossRef]
- Temudo, M.F.; Muyzer, G.; Kleerebezem, R.; van Loosdrecht, M.C. Diversity of microbial communities in open mixed culture fermentations: Impacts of the pH and carbon source. Appl. Microbiol. Biotechnol. 2008, 80, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Heyndrickx, M.; de Vos, P.; de Ley, J. Fermentation characteristics of Clostridium pasteurianum LMG 3285 grown on glucose and mannitol. J. Appl. Bacteriol. 1991, 70, 52–58. [Google Scholar] [CrossRef]
- Montville, T.J.; Parris, N.; Conway, L.K. Influence of pH on organic acid production by Clostridium sporogenes in test tube and fermenter cultures. Appl. Environ. Microbiol. 1985, 49, 733–736. [Google Scholar] [PubMed]
- Stinson, E.E.; Naftulin, K.A. Effect of pH on organic acid production by Clostridium propionicum in test tube and fermenter cultures. J. Ind. Microbiol. 1991, 8, 59–64. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, S.T. Effect of pH on metabolic pathway shift in fermentation of xylose by Clostridium tyrobutyricum. J. Biotechnol. 2004, 110, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Temudo, M.F.; Poldermans, R.; Kleerebezem, R.; van Loosdrecht, M. Glycerol fermentation by (open) mixed cultures: A chemostat study. Biotechnol. Bioeng. 2008, 100, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; van Gemert, J.M.; Zoetemeyer, R.J.; Breure, A.M. Main characteristics and stoichiometric spects of acidogenesis of soluble carbonhydrate containing wastewater. Process Biochem. 1984, 19, 228–232. [Google Scholar]
- Ren, N.Q.; Chua, H.; Chan, S.Y.; Tsang, Y.F.; Wang, Y.J.; Sin, N. Assessing optimal fermentation type for bio-hydrogen production in continous-flow acidogenic reators. Bioresour. Technol. 2007, 98, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.J.; Yu, H.Q. Roles of pH in biologic production of hydrogen and volatile fatty acids from glucose by enriched anaerobic cultures. Appl. Biochem. Biotechnol. 2004, 112, 79–90. [Google Scholar] [CrossRef]
- Van Ginkel, S.; Logan, B. Inhibition of biohydrogen by undissociated acetic and butyric acids. Environ. Sci. Technol. 2005, 39, 9351–9356. [Google Scholar] [CrossRef] [PubMed]
- Bastidas-Oyanedel, J.R.; Mohd-Zaki, Z.; Pratt, S.; Steyer, J.P.; Batstone, D.J. Development of membrane inlet mass spectrometry for examination of fermentation processes. Talanta 2010, 83, 482–492. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Batstone, D.J. Teaching uncertainty propagation as a core component in process engineering statistics. Educ. Chem. Eng. 2013, 8, e132–e139. [Google Scholar] [CrossRef]
- Lu, Y.; Slater, F.R.; Mohd-Zaki, Z.; Pratt, S.; Batstone, D.J. Impact of operating history on mixed culture fermentation microbial ecology and product mixture. Water Sci. Technol. 2011, 64, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [PubMed]
- Lynd, L.R.; Grethlein, H.E.; Wplkin, R.H. Fermentation of cellulosic substrates in batch and continuous culture by Clostridium thermocellum. Appl. Environ. Microbiol. 1989, 55, 3131–3139. [Google Scholar] [PubMed]
- Ramachandran, U.; Wrana, N.; Cicek, N.; Sparling, R.; Levin, D.B. Isolation and characterization of a hydrogen- and ethanol-producing Clostridium sp. strain URNW. Can. J. Mictobiol. 2011, 57, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Brisse, S.; Grimont, F.; Grimont, P.D. The genus klebsiella. In The Prokaryotes; Springer: New York, NY, USA, 2006; pp. 159–196. [Google Scholar]
- Teixera de Mattos, M.J.; Tempest, D.W. Metabolic and energetic aspects of the growth of Klebsiella aerogenes NCTC 418 on glucose in anaerobic chemostat culture. Arch. Microbiol. 1983, 134, 80–85. [Google Scholar] [CrossRef]
- Jarvis, G.N.; Moore, E.R.B.; Thiele, J.H. Formate and ethanol are the major products of glycerol fermentation produced by a Klebsiella planticola strain isolated from red deer. J. Appl. Microbiol. 1997, 83, 166–174. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd-Zaki, Z.; Bastidas-Oyanedel, J.R.; Lu, Y.; Hoelzle, R.; Pratt, S.; Slater, F.R.; Batstone, D.J. Influence of pH Regulation Mode in Glucose Fermentation on Product Selection and Process Stability. Microorganisms 2016, 4, 2. https://doi.org/10.3390/microorganisms4010002
Mohd-Zaki Z, Bastidas-Oyanedel JR, Lu Y, Hoelzle R, Pratt S, Slater FR, Batstone DJ. Influence of pH Regulation Mode in Glucose Fermentation on Product Selection and Process Stability. Microorganisms. 2016; 4(1):2. https://doi.org/10.3390/microorganisms4010002
Chicago/Turabian StyleMohd-Zaki, Zuhaida, Juan R. Bastidas-Oyanedel, Yang Lu, Robert Hoelzle, Steven Pratt, Fran R. Slater, and Damien J. Batstone. 2016. "Influence of pH Regulation Mode in Glucose Fermentation on Product Selection and Process Stability" Microorganisms 4, no. 1: 2. https://doi.org/10.3390/microorganisms4010002