
The Opportunity for High-Performance Biomaterials from Methane

 ,
,
Abstract
:1. Introduction


- (1)
- A potential approach for carbon sequestration and GHGs emission reduction;
- (2)
- Production of biodegradable polymers for replacing conventional fossil fuel-derived plastics;
- (3)
- Reducing the use of organic carbon sources such as sugars for PHB production [6].
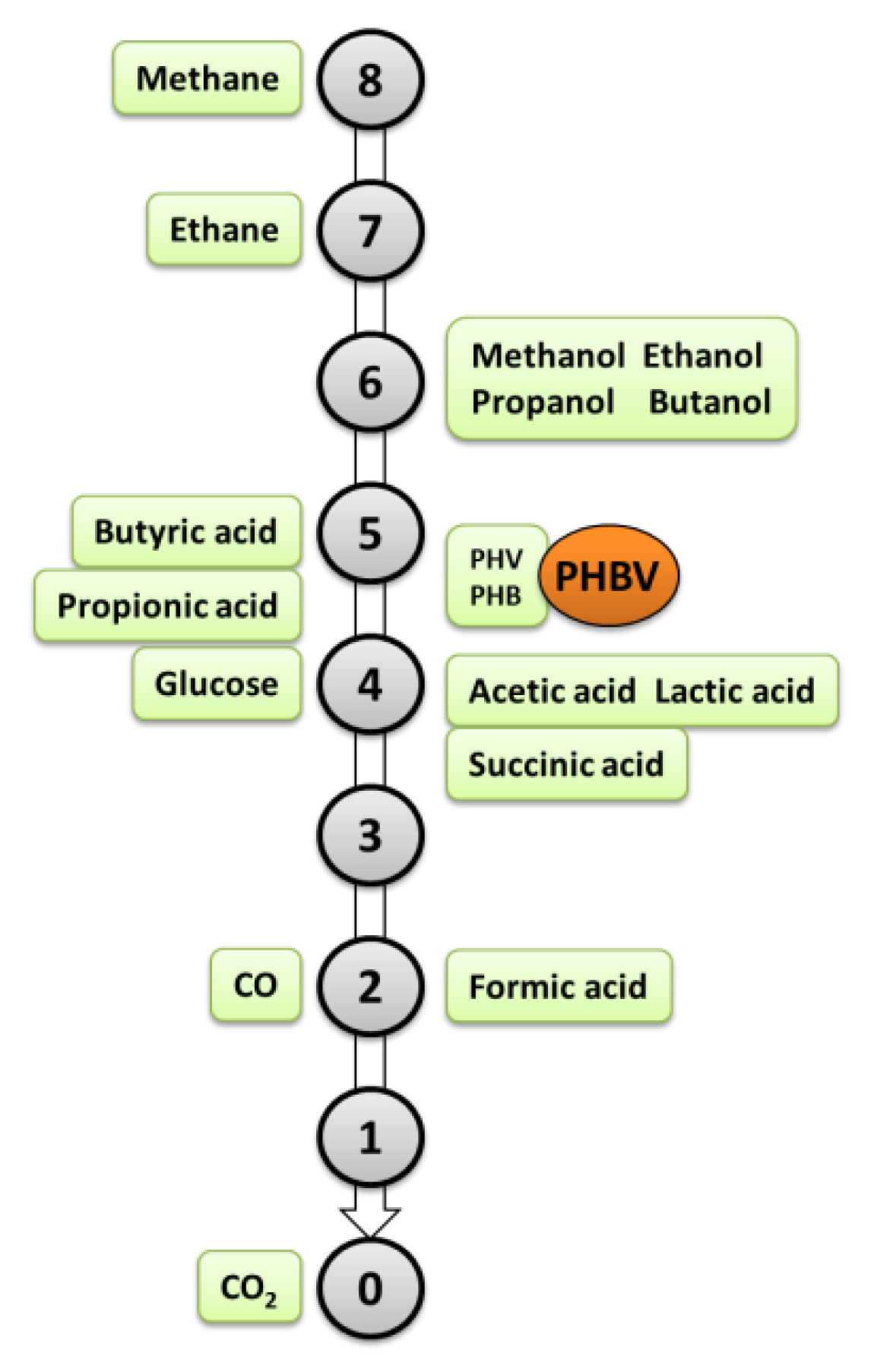
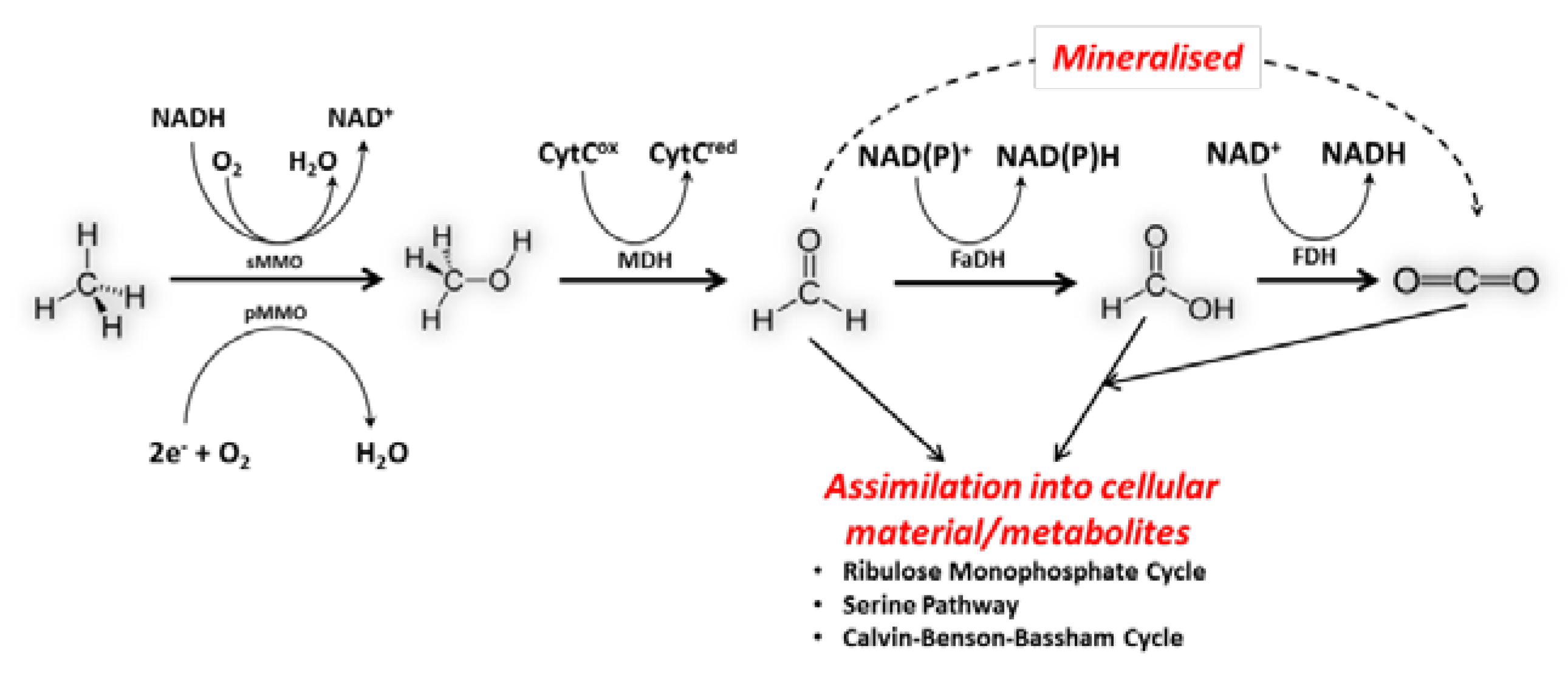
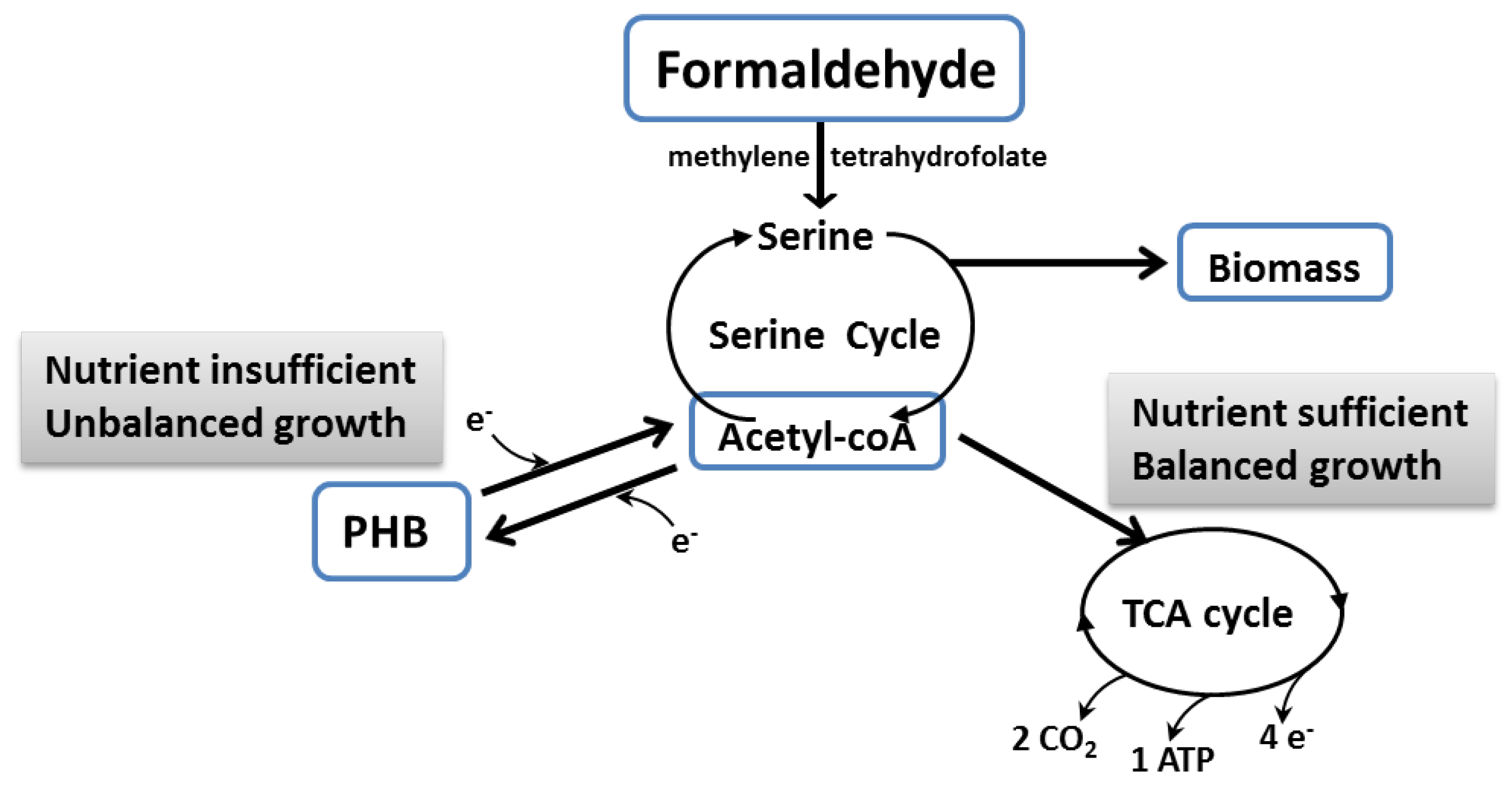
2. Metabolism of Methanotrophs and Biosynthesis of PHB from Methane


{kind=link}
{kind=link}
{kind=link}
| Microorganisms (% in Mixed Culture) | Carbon Source | PHB Content (% of Total Biomass) | Yield (g PHB/g Carbon Source) | References |
|---|---|---|---|---|
| Methlocystis sp. GB25 DSMZ 7674 (>90%) | Methane | 28.3–51.3 | 0.55 | [27,57,58] |
| Methlocystis sp. GB25 DSMZ 7674 (>86%) | Methane | 10.4–33.6 | 0.45 | [37] |
| Methylosinus/methylocystis (percent unknown) + Type I genera | Methane | 7–46 | n/a | [63] |
| Mixed consortium including Methylobacterium organophilum (percent unknown) | Methane | 34 | 0.80* | [59] |
| Methylosinus and Methylocystis spp. dominant in mixed consortium (percent unknown) | Methane | 17–26 | n/a | [67] |
| Methylocystis (~77%) | Methane | 39 | 0.64 * | [68] |
| Methylocystis (> 76%) | Methane | 40 | 0.49 * | [69] |
| Methylomicrobium (96%) | Methane | 23 | n/a | [70] |
| Methylocystis sp. (71.6%) | Methane | 2.5–8.5 | n/a | [71] |
| Methylosinus trichosporium | Methane | 20–25 | n/a | [72] |
| Methylocystis parvus OBBP | Methane | 68 | n/a | [38] |
| Methylosinus trichosporium OB3b | Methane | 30–50 | n/a | [73] |
| Type II methanotrophic strain MTS | Methane | 3 | n/a | [35] |
| Methylosinus trichosporium OB3b | Methane | 30 | n/a | [74] |
| Microorganisms (Pure Culture) | Carbon Source | PHB Content (% of Total Biomass) | Yield (g PHB/g Carbon Source) | References |
| Methylosinus trichosporium OB3b Methylobacterium organophilum strains | Methane Methane | 25 38–57 | n/a 0.48–0.59 * | [59] |
| Methylocystis spp. Methylosinus spp. | Methane Methane | 7–36 9–38 | n/a n/a | [63] |
| Methylocystis hirsute | Methane | 51.6 | n/a | [75] |
| Methylocystis parvus OBBP Methylosinus trichosporium OB3b | Methane | 60 29 | 0.88 1.13 | [76] |
| Methylocystis parvus OBBP | Methane | 49.4 | n/a | [77] |
| Methylosinus trichosporium IMV3011 | Methane + methanol | 40 | n/a | [36] |
| Methylosinus trichosporium IMV 3011 | Methane + Methanol | 46 | n/a | [78] |
| Pseudomonas sp. K | Methanol | 66 | 0.18 | [79] |
| Methylobacterium rhodesianum | Methanol | 45–55 | n/a | [65] |
| Methylobacterium extorquens K Paracoccus denitrificans | Methanol + n-amyl alcohol | 44 57 | 0.11 (0.97 on n-amyl alcohol) | [80] |
| Pseudomonas 135 | Methanol | 55 | n/a | [81] |
| Methylobacterium extorquens NCIMB 9133 | Methanol | 7–21 | [82] | |
| Methylobacterium extorquens ATCC 55366 | Methanol | 40–46 | 0.09–0.12 | [55] |
| Methylobacterium organophilum | Methanol | 52–56 | 0.19 | [83] |
| Methylobacterium sp V49 | Methanol | 11 | n/a | [84] |
| Methylobacterium extorquens AM1 | Methanol | 34–42 | n/a | [85] |
| Methylobacterium extorquens AM1 | Methanol | 22–25 | n/a | [56] |
| Methylobacterium sp. GW2 | Methanol | 40 | n/a | [86] |
| Methylobacterium extorquens DSMZ 1340 | Methanol | 35 | 0.3 | [87] |
| Methylosinus trichosporium IMV 3011 | Methanol | 32 | n/a | [88] |
| Methylobacterium extorquens AM1 | Methanol | 27 | n/a | [89] |
| Methylobacteria extorquens G10 Methyloligella halotolerans C2 | Methanol | 40 17 | n/a | [90] |
3. Process Conditions for PHB Production from Methane
4. High-Performance Biomaterials from Methane: PHA Co-Polymers
| Polymer | Melting Temperature Tm (◦C) | Glass-Transition Temperature Tg (◦C) | Young’s Modulus (GPa) | Tensile Strength (MPa) | Elongation to Break (%) |
|---|---|---|---|---|---|
| PHB | 180 | 4 | 3.5 | 40 | 5 |
| P(3HB-co-20 mol%3HV) | 145 | −1 | 0.8 | 20 | 50 |
| P(3HB-co-6 mol%3HA) * | 133 | −8 | 0.2 | 17 | 680 |
| Polypropylene | 176 | −10 | 1.7 | 38 | 400 |
| Low-density polyethylene | 130 | −30 | 0.2 | 10 | 620 |
5. Strategies for Producing PHA Co-Polymers from Methane
5.1. Direct Methanotrophic PHA Synthesis by Adding External Precursors
5.2. Indirect Biological PHA Production: Synthesis in Consortia
5.3. Downstream Processing of PHA—Including Precursor Production from Methane
5.4. Synopsis
| Process | Pros | Cons | Unknowns |
|---|---|---|---|
| Direct production of PHB from CH4 | Has been achieved | Average to poor mechanical properties | Processes for maximising yields and rates |
| Methanotroph PHBV production through the co-addition of a fatty acid with odd carbon numbers | Has been achieved | The cost of the Codd feedstock offsets the savings of CH4; may produce blends of polymer product if there is variation in uptake rates amongst the different members of the community | Yields, homogeneity and maximum co-monomer content |
| Methanotrophs facilitating growth of a co-culture capable of PHBV production (direct or co-substrate addition) | Potentially achievable using Ralstonia sp. | The cost of the Codd feedstock offsets the savings of CH4; may produce blends of polymer product if there is variation in uptake rates amongst the different members of the community | Yields, homogeneity and maximum co-monomer content |
| Generate alternative copolymers through supply of appropriate feed. | Delivers broader range of mechanical properties | Cost of adding the monomers | Yields, homogeneity and maximum co-monomer content |
| Use a co-feeding strategy of timed pulses of methane and alternative feeds to tailor copolymer compositional distribution | Tailored, e.g., block copolymers already produced in the literature using alternating feeding strategy | Not proven in methanotrophs. May produce blends | Pulses of gas feed alternating with soluble carbon feed may prove difficult for cells to adapt to |
| Generate monomers biologically and polymerise ex situ | Can achieve desired copolymer composition | Monomer concentration and purification | Currently being commercialised |
| Downstream polymer modification/functionalisation/ depolymerisation into oligomers and use as building block | Processes are well established | Costly, intensive additional processes and can be time-consuming | Potential for development of unique material properties for niche applications. |
6. PHA Production from Other Gas Streams
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Laycock, B.; Halley, P.; Pratt, S.; Werker, A.; Lant, P. The chemomechanical properties of microbial polyhydroxyalkanoates. Prog. Polym. Sci. 2013, 38, 536–583. [Google Scholar] [CrossRef]
- Chen, G. Plastics completely synthesized by bacteria: Polyhydroxyalkanoates. In Plastics from Bacteria: Natural Functions and Applications; Chen, G., Ed.; Springer-Verlag: Berlin Heidelberg, 2010; Volume 14, pp. 17–37. [Google Scholar]
- Anderson, A.J.; Dawes, E.A. Occurrence, metabolism, metabolic role, and industrial uses of bacterial polyhydroxyalkanoates. Microbiol. Rev. 1990, 54, 450–472. [Google Scholar] [PubMed]
- Lee, S.Y. Bacterial polyhydroxyalkanoates. Biotechnol. Bioeng. 1996, 49, 1–14. [Google Scholar] [CrossRef]
- Jendrossek, D.; Knoke, I.; Habibian, R.; Steinbüchel, A.; Schlegel, H. Degradation of poly(3-hydroxybutyrate), PHB, by bacteria and purification of a novel PHB depolymerase from Comamonas sp. J. Environ. Polym. Degr. 1993, 1, 53–63. [Google Scholar] [CrossRef]
- Karthikeyan, O.; Karthigeyan, C.; Cirés, S.; Heimann, K. Review of sustainable methane mitigation and biopolymer production. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1579–1610. [Google Scholar] [CrossRef]
- Tempest, D.W.; Wouters, J.T.M. Properties and performance of microorganisms in chemostat culture. Enzyme Microb. Technol. 1981, 3, 283–290. [Google Scholar] [CrossRef]
- Gurieff, N.; Lant, P. Comparative life cycle assessment and financial analysis of mixed culture polyhydroxyalkanoate production. Bioresour. Technol. 2007, 98, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Halami, P.M. Production of polyhydroxyalkanoate from starch by the native isolate Bacillus cereus CFR06. World J. Microb. Biotechnol. 2008, 24, 805–812. [Google Scholar] [CrossRef]
- Choi, J.I.; Lee, S.Y. High-level production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by fed-batch culture of recombinant Escherichia coli. Appl. Environ. Microbiol. 1999, 65, 4363–4368. [Google Scholar] [PubMed]
- Yamane, T. Yield of poly-d(−)-3-hydroxybutyrate from various carbon-sources—A theoretical study. Biotechnol. Bioeng. 1993, 41, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Nikodinovic-Runic, J.; Guzik, M.; Kenny, S.T.; Babu, R.; Werker, A.; O Connor, K.E. Carbon-rich wastes as feedstocks for biodegradable polymer (polyhydroxyalkanoate) production using bacteria. Adv. Appl. Microbiol. 2013, 84, 139–200. [Google Scholar] [PubMed]
- Koller, M.; Salerno, A.; Muhr, A.; Reiterer, A.; Braunegg, G. Polyhydroxyalkanoates: Biodegradable polymers and plastics from renewable resources. Mater. Tehnol. 2013, 47, 5–12. [Google Scholar]
- Wendlandt, K.D.; Stottmeister, U.; Helm, J.; Soltmann, B.; Jechorek, M.; Beck, M. The potential of methane-oxidizing bacteria for applications in environmental biotechnology. Eng. Life Sci. 2010, 10, 87–102. [Google Scholar] [CrossRef]
- Rehm, B.H.A. Polyester synthases: Natural catalysts for plastics. Biochem. J. 2003, 376, 15–33. [Google Scholar] [CrossRef] [PubMed]
- US Environmental Protection Agency. Methane and Nitrous Oxide Emissions from Natural Sources. EPA 430-R-10–001. Available online: http://nepis.epa.gov (accessed on 20 November 2015).
- Intergovernmental Panel on Climate Change (IPCC). IPCC Fourth Assessment Report (AR4). Climate Change 2007: The Physical Science Basis. Available online: http://www.ipcc.ch/publications_and_data/publications_ipcc_fourth_assessment_report_wg1_report_the_physical_science_basis.htm (accessed on 20 November 2015).
- Abbasi, T.; Tauseef, S.M.; Abbasi, S.A. Anaerobic digestion for global warming control and energy generation—An overview. Renew Sust. Energ. Rev. 2012, 16, 3228–3242. [Google Scholar] [CrossRef]
- Batstone, D.J.; Jensen, P.D. 4.17—Anaerobic processes. In Treatise on Water Science; Wilderer, P., Ed.; Elsevier: Oxford, UK, 2011; pp. 615–639. [Google Scholar]
- Rittmann, B.E. Opportunities for renewable bioenergy using microorganisms. Biotechnol. Bioeng. 2008, 100, 203–212. [Google Scholar] [CrossRef] [PubMed]
- WorldBank. Zero Routine Flaring by 2030. Available online: http://www.worldbank.org/en/programs/zero-routine-flaring-by-2030 (accessed on 20 November 2015).
- Tijm, P.J.A.; Waller, F.J.; Brown, D.M. Methanol technology developments for the new millennium. Appl. Catal. Gen. 2001, 221, 275–282. [Google Scholar] [CrossRef]
- Kuo, J.W. Engineering evaluation of direct methane conversion processes. In Methane Conversion by Oxidative Processes; Wolf, E.E., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 1992; pp. 483–526. [Google Scholar]
- Dorin, H.; Demmin, P.; Gabriel, D.L. Chemistry—The Study of Matter, 4th Ed. ed; Prentice Hall: Needham, MA, USA, 1987. [Google Scholar]
- Rostkowski, K.H. Understanding Methanotrophic Polyhydroxybutyrate (PHB) Production Across Scale: Life Cycle Assessment, Pure Culture Experimentation, and Pathway/Genome Database Development. Ph.D. Thesis, Stanford University, Stanford, CA, USA, 2012. [Google Scholar]
- U.S. Energy Information Administration, U.S. Price of Natural Gas Sold to Commercial Consumers, Monthly. Available online: http://www.eia.gov/opendata/qb.cfm?sdid=NG.N3020US3.M (accessed on 20 November 2015).
- Wendlandt, K.D.; Jechorek, M.; Helm, J.; Stottmeister, U. Producing poly-3-hydroxybutyrate with a high molecular mass from methane. J. Biotechnol. 2001, 86, 127–133. [Google Scholar] [CrossRef]
- Levett, I.; Birkett, G.; Davies, N.; Bell, A.; Langford, A.; Laycock, B.; Lant, P.; Pratt, S. Techno-economic assessment of poly-3-hydroxybutyrate (PHB) production from methane. Eng. Res. Design 2016. under review. [Google Scholar]
- Kracke, F.; Kromer, J.O. Identifying target processes for microbial electrosynthesis by elementary mode analysis. BMC Bioinform. 2014. [Google Scholar] [CrossRef] [PubMed]
- Pratt, S. The Development of the TOGA Sensor for the Study of Biological Wastewater Treatment Systems. Ph.D. Thesis, The University of Queensland, Queensland, Australia, 2003. [Google Scholar]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [PubMed]
- Kalyuzhnaya, M.G.; Puri, A.W.; Lidstrom, M.E. Metabolic engineering in methanotrophic bacteria. Metab. Eng. 2015, 29, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Strong, P.J.; Xie, S.; Clarke, W.P. Methane as a resource: Can the methanotrophs add value? Environ. Sci. Technol. 2015, 49, 4001–4018. [Google Scholar] [CrossRef] [PubMed]
- Whittenbury, R.; Phillips, K.C.; Wilkinson, J.F. Enrichment, isolation and some properties of methane-utilizing bacteria. J. Gen. Microbiol. 1970, 61, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Vecherskaya, M.; Dijkema, C.; Stams, A.J.M. Intracellular PHB conversion in a Type II methanotroph studied by 13c NMR. J. Ind. Microbiol. Biotechnol. 2001, 26, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Xin, J.Y.; Chen, L.L.; Song, H.; Xia, C.U. Biosynthesis of poly-3-hydroxybutyrate with a high molecular weight by methanotroph from methane and methanol. J. Nat. Gas. Chem. 2008, 17, 103–109. [Google Scholar] [CrossRef]
- Helm, J.; Wendlandt, K.D.; Jechorek, M.; Stottmeister, U. Potassium deficiency results in accumulation of ultra-high molecular weight poly-beta-hydroxybutyrate in a methane-utilizing mixed culture. J. Appl. Microbiol. 2008, 105, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Asenjo, J.A.; Suk, J.S. Microbial conversion of methane into poly-β-hydroxybutyrate (PHB): Growth and intracellular product accumulation in a type-II methanotroph. J. Ferment. Bioeng. 1986, 64, 271–278. [Google Scholar] [CrossRef]
- Fei, Q.; Guarnieri, M.T.; Tao, L.; Laurens, L.M.L.; Dowe, N.; Pienkos, P.T. Bioconversion of natural gas to liquid fuel: Opportunities and challenges. Biotechnol. Adv. 2014, 32, 596–614. [Google Scholar] [CrossRef] [PubMed]
- Shiemke, A.K.; Cook, S.A.; Miley, T.; Singleton, P. Detergent solubilization of membrane-bound methane monooxygenase requires plastoquinol analogs as electron donors. Arch. Biochem. Biophys. 1995, 321, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Anthony, C. The quinoprotein dehydrogenases for methanol and glucose. Arch. Biochem. Biophys. 2004, 428, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Culpepper, M.A.; Rosenzweig, A.C. Structure and protein-protein interactions of methanol dehydrogenase from Methylococcus capsulatus (Bath). Biochemistry 2014, 53, 6211–6219. [Google Scholar] [CrossRef] [PubMed]
- Fassel, T.A.; Buchholz, L.A.; Collins, M.L.; Remsen, C.C. Localization of methanol dehydrogenase in two strains of methylotrophic bacteria detected by immunogold labeling. Appl. Environ. Microbiol. 1992, 58, 2302–2307. [Google Scholar] [PubMed]
- Kitmitto, A.; Myronova, N.; Basu, P.; Dalton, H. Characterization and structural analysis of an active particulate methane monooxygenase trimer from Methylococcus capsulatus (Bath). Biochemistry-US 2005, 44, 10954–10965. [Google Scholar] [CrossRef] [PubMed]
- Myronova, N.; Kitmitto, A.; Collins, R.F.; Miyaji, A.; Dalton, H. Three-dimensional structure determination of a protein supercomplex that oxidizes methane to formaldehyde in Methylococcus capsulatus (Bath). Biochemistry 2006, 45, 11905–11914. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M.G.; Yang, S.; Rozova, O.N.; Smalley, N.E.; Clubb, J.; Lamb, A.; Gowda, G.A.N.; Raftery, D.; Fu, Y.; Bringel, F.; et al. Highly efficient methane biocatalysis revealed in a methanotrophic bacterium. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Rahalkar, M. Aerobic Methanotrophic Bacterial Communities in Sediments of Lake Constance. Ph.D. Thesis, Universitat Konstanz, Konstanz, Germany, 2006. [Google Scholar]
- Anthony, C. How half a century of research was required to understand bacterial growth on C1 and C2 compounds; the story of the serine cycle and the ethylmalonyl-CoA pathway. Sci. Prog. 2011, 94, 109–137. [Google Scholar] [CrossRef] [PubMed]
- Vorholt, J.A.; Marx, C.J.; Lidstrom, M.E.; Thauer, R.K. Novel formaldehyde-activating enzyme in Methylobacterium extorquens AM1 required for growth on methanol. J. Bacteriol. 2000, 182, 6645–6650. [Google Scholar] [CrossRef] [PubMed]
- Vincenzini, M.; de Philippis, R. Polyhydroxyalkanoates. In Chemicals from Microalgae; Cohen, Z., Ed.; Taylor and Francis: London, UK, 1999; pp. 290–312. [Google Scholar]
- Sheu, D.S.; Wang, Y.T.; Lee, C.Y. Rapid detection of polyhydroxyalkanoate-accumulating bacteria isolated from the environment by colony PCR. Microbiol. UK 2000, 146, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Madison, L.L.; Huisman, G.W. Metabolic engineering of poly(3-hydroxyalkanoates): From DNA to plastic. Microbiol. Mol. Biol. R 1999, 63, 21–53. [Google Scholar]
- York, G.M.; Stubbe, J.; Sinskey, A.J. The Ralstonia eutropha PhaR protein couples synthesis of the PhaP phasin to the presence of polyhydroxybutyrate in cells and promotes polyhydroxybutyrate production. J. Bacteriol. 2002, 184, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Pieja, A.J.; Sundstrom, E.R.; Criddle, C.S. Poly-3-hydroxybutyrate metabolism in the type II methanotroph Methylocystis parvus OBBP. Appl. Environ. Microbiol. 2011, 77, 6012–6019. [Google Scholar] [CrossRef] [PubMed]
- Bourque, D.; Pomerleau, Y.; Groleau, D. High cell density production of poly-b-hydroxybutyrate (PHB) from methanol by Methylobacterium extorquens: Production of high-molecular-mass PHB. Appl. Microbiol. Biotechnol. 1995, 44, 367–376. [Google Scholar] [CrossRef]
- Korotkova, N.; Chistoserdova, L.; Lidstrom, M.E. Poly-b-hydroxybutyrate biosynthesis in the facultative methylotroph Methylobacterium extorquens AM1: Identification and mutation of gap11, gap20, and phaR. J. Bacteriol. 2002, 184, 6174–6181. [Google Scholar] [CrossRef] [PubMed]
- Wendlandt, K.D.; Jechorek, M.; Helm, J.; Stottmeister, U. Production of PHB with a high molecular mass from methane. Polym. Degrad. Stabil. 1998, 59, 191–194. [Google Scholar] [CrossRef]
- Helm, J.; Wendlandt, K.D.; Rogge, G.; Kappelmeyer, U. Characterizing a stable methane-utilizing mixed culture used in the synthesis of a high-quality biopolymer in an open system. J. Appl. Microbiol. 2006, 101, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga, C.; Morales, M.; Le Borgne, S.; Revah, S. Production of poly-β-hydroxybutyrate (PHB) by Methylobacterium organophilum isolated from a methanotrophic consortium in a two-phase partition bioreactor. J. Hazard Mater 2011, 190, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J. The methanotrophs—The families methylococcaceae and methylocystaceae. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 266–289. [Google Scholar]
- Bowman, J.P.; Jiménez, L.; Rosario, I.; Hazen, T.C.; Sayler, G.S. Characterization of the methanotrophic bacterial community present in a trichloroethylene-contaminated subsurface groundwater site. Appl. Environ. Microbiol. 1993, 59, 2380–2387. [Google Scholar] [PubMed]
- Heyer, J.; Berger, U.; Hardt, M.; Dunfield, P.F. Methylohalobius crimeensis gen. nov., sp. nov., a moderately halophilic, methanotrophic bacterium isolated from hypersaline lakes of Crimea. Int. J. Syst. Evol. Microbiol. 2005, 55, 1817–1826. [Google Scholar] [CrossRef] [PubMed]
- Pieja, A.J.; Rostkowski, K.H.; Criddle, C.S. Distribution and selection of poly-3-hydroxybutyrate production capacity in methanotrophic proteobacteria. Microb. Ecol. 2011, 62, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Follner, C.G.; Babel, W.; Valentin, H.E.; Steinbuchel, A. Expression of polyhydroxyalkanoic-acid-biosynthesis genes in methylotrophic bacteria relying on the ribulose monophosphate pathway. Appl. Microbiol. Biotechnol. 1993, 40, 284–291. [Google Scholar] [CrossRef]
- Babel, W. Pecularities of methylotrophs concerning overflow metabolism, especially the synthesis of polyhydroxyalkanoates. Fems. Microbiol. Lett. 1992, 103, 141–148. [Google Scholar] [CrossRef]
- Khosravi-Darani, K.; Mokhtari, Z.B.; Amai, T.; Tanaka, K. Microbial production of poly (hydroxybutyrate) from C-1 carbon sources. Appl. Microbiol. Biotechnol. 2013, 97, 1407–1424. [Google Scholar] [CrossRef] [PubMed]
- Pfluger, A.R.; Wu, W.M.; Pieja, A.J.; Wan, J.; Rostkowski, K.H.; Criddle, C.S. Selection of Type I and Type II methanotrophic proteobacteria in a fluidized bed reactor under non-sterile conditions. Bioresour. Technol. 2011, 102, 9919–9926. [Google Scholar] [CrossRef] [PubMed]
- Myung, J.; Wang, Z.Y.; Yuan, T.; Zhang, P.; van Nostrand, J.D.; Zhou, J.Z.; Criddle, C.S. Production of nitrous oxide from nitrite in stable Type II methanotrophic enrichments. Environ. Sci. Technol. 2015, 49, 10969–10975. [Google Scholar] [CrossRef] [PubMed]
- Myung, J.; Galega, W.M.; van Nostrand, J.D.; Yuan, T.; Zhou, J.Z.; Criddle, C.S. Long-term cultivation of a stable Methylocystis-dominated methanotrophic enrichment enabling tailored production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate). Bioresour. Technol. 2015, 198, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Chidambarampadmavathy, K.; Karthikeyan, O.P.; Heimann, K. Biopolymers made from methane in bioreactors. Eng. Life Sci. 2015, 15, 689–699. [Google Scholar] [CrossRef]
- Karthikeyan, O.P.; Chidambarampadmavathy, K.; Nadarajan, S.; Lee, P.K.H.; Heimann, K. Effect of CH4/O2 ratio on fatty acid profile and polyhydroxybutyrate content in a heterotrophic-methanotrophic consortium. Chemosphere 2015, 141, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.; Brannan, J.; Higgins, I.J. The effect of growth conditions on intracytoplasmic membranes and methane mono-oxygenase activities in Methylosinus trichosporium OB3b. Microbiology 1981, 125, 63–72. [Google Scholar] [CrossRef]
- Shah, N.N.; Hanna, M.L.; Taylor, R.T. Batch cultivation of Methylosinus trichosporium OB3b: V. Characterization of poly-b-hydroxybutyrate production under methane-dependent growth conditions. Biotechnol. Bioeng. 1996, 49, 161–171. [Google Scholar] [CrossRef]
- Doronina, N.V.; Ezhov, V.A.; Trotsenko, Y.A. Growth of Methylosinus trichosporium OB3b on methane and poly-b-hydroxybutyrate biosynthesis. Appl. Biochem. Microbiol. 2008, 44, 182–185. [Google Scholar] [CrossRef]
- Rahnama, F.; Vasheghani-Farahani, E.; Yazdian, F.; Shojaosadati, S.A. PHB production by Methylocystis hirsuta from natural gas in a bubble column and a vertical loop bioreactor. Biochem. Eng. J. 2012, 65, 51–56. [Google Scholar] [CrossRef]
- Rostkowski, K.H.; Pfluger, A.R.; Criddle, C.S. Stoichiometry and kinetics of the PHB-producing Type II methanotrophs Methylosinus trichosporium OB3b and Methylocystis parvus OBBP. Bioresour. Technol. 2013, 132, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Sundstrom, E.R.; Criddle, C.S. Optimization of methanotrophic growth and production of poly(3-hydroxybutyrate) in a high-throughput microbioreactor system. Appl. Environ. Microbiol. 2015, 81, 4767–4773. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, Y.X.; Kong, W.B.; Xia, C.G. Activities of key enzymes in the biosynthesis of poly-3-hydroxybutyrate by Methylosinus trichosporium IMV3011. Chin. J. Catal. 2012, 33, 1754–1761. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamane, T.; Shimizu, S. Mass production of poly-β-hydroxybutyric acid by fully automatic fed-batch culture of methylotroph. Appl. Microbiol. Biotechnol. 1986, 23, 322–329. [Google Scholar] [CrossRef]
- Ueda, S.; Matsumoto, S.; Takagi, A.; Yamane, T. Synthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from methanol and n-amyl alcohol by the methylotrophic bacteria Paracoccus denitrificans and Methylobacterium extorquens. Appl. Environ. Microbiol. 1992, 58, 3574–3579. [Google Scholar] [PubMed]
- Daniel, M.; Choi, J.H.; Kim, J.H.; Lebeault, J.M. Effect of nutrient deficiency on accumulation and relative molecular weight of poly-β-hydroxybutyric acid by methylotrophic bacterium, Pseudomonas 135. Appl. Microbiol. Biotechnol. 1992, 37, 702–706. [Google Scholar] [CrossRef]
- Taidi, B.; Anderson, A.J.; Dawes, E.A.; Byrom, D. Effect of carbon source and concentration on the molecular-mass of poly(3-hydroxybutyrate) produced by Methylobacterium extorquens and Alcaligenes eutrophus. Appl. Microbiol. Biotechnol. 1994, 40, 786–790. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, P.; Lee, H.S.; Kim, J.H. High production of poly-b-hydroxybutyrate (PHB) from Methylobacterium organophilum under potassium limitation. Biotechnol. Lett. 1996, 18, 25–30. [Google Scholar] [CrossRef]
- Ghatnekar, M.S.; Pai, J.S.; Ganesh, M. Production and recovery of poly-3-hydroxybutyrate from Methylobacterium sp V49. J. Chem. Technol. Biotechnol. 2002, 77, 444–448. [Google Scholar] [CrossRef]
- Korotkova, N.; Lidstrom, M.E. Connection between poly-b-hydroxybutyrate biosynthesis and growth on C-1 and C-2 compounds in the methylotroph Methylobacterium extorquens AM1. J. Bacteriol. 2001, 183, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Yezza, A.; Fournier, D.; Halasz, A.; Hawari, J. Production of polyhydroxyalkanoates from methanol by a new methylotrophic bacterium Methylobacterium sp GW2. Appl. Microbiol. Biotechnol. 2006, 73, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari-Hosseini, Z.B.; Vasheghani-Farahani, E.; Shojaosadati, S.A.; Karimzadeh, R.; Heidarzadeh-Vazifekhoran, A. Effect of feed composition on PHB production from methanol by HCDC of Methylobacterium extorquens (DSMZ 1340). J. Chem. Technol. Biotechnol. 2009, 84, 1136–1139. [Google Scholar] [CrossRef]
- Xin, J.Y.; Zhang, Y.X.; Dong, J.; Song, H.; Xia, C.G. An experimental study on molecular weight of poly-3-hydroxybutyrate (PHB) accumulated in Methylosinus trichosporium IMV 3011. Afr. J. Biotechnol. 2011, 10, 7078–7087. [Google Scholar]
- Orita, I.; Nishikawa, K.; Nakamura, S.; Fukui, T. Biosynthesis of polyhydroxyalkanoate copolymers from methanol by Methylobacterium extorquens AM1 and the engineered strains under cobalt-deficient conditions. Appl. Microbiol. Biotechnol. 2014, 98, 3715–3725. [Google Scholar] [CrossRef] [PubMed]
- Poroshina, M.N.; Doronina, N.V.; Ezhov, V.A.; Trotsenko, Y.A. Comparative characteristics of biosynthesis of polyhydroxybutyrate from methanol by Methylobacteria extorquens G10 and Methyloligella halotolerans C2. Appl. Biochem. Microbiol. 2014, 50, 253–258. [Google Scholar] [CrossRef]
- Pieja, A.J.; Sundstrom, E.R.; Criddle, C.S. Cyclic, alternating methane and nitrogen limitation increases PHB production in a methanotrophic community. Bioresour. Technol. 2012, 107, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Listewnik, H.F.; Wendlandt, K.D.; Jechorek, M.; Mirschel, G. Process design for the microbial synthesis of poly-b-hydroxybutyrate (PHB) from natural gas. Eng. Life Sci. 2007, 7, 278–282. [Google Scholar] [CrossRef]
- Johnson, K.; van Loosdrecht, M.C.M.; Kleerebezem, R. Influence of ammonium on the accumulation of polyhydroxybutyrate (PHB) in aerobic open mixed cultures. J. Biotechnol. 2010, 147, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; de Roy, K.; Thas, O.; de Neve, J.; Hoefman, S.; Vandamme, P.; Heylen, K.; Boon, N. The more, the merrier: Heterotroph richness stimulates methanotrophic activity. ISME J. 2014, 8, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.C.; Quijano, G.; Perez, R.; Munoz, R. Assessing the influence of CH4 concentration during culture enrichment on the biodegradation kinetics and population structure. J. Environ. Manag. 2014, 146, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.G.; McArthur, J.V.; Shimkets, L.J. Methanotroph diversity in landfill soil: Isolation of novel type I and type II methanotrophs whose presence was suggested by culture-independent 16S ribosomal DNA analysis. Appl. Environ. Microbiol. 1999, 65, 4887–4897. [Google Scholar] [PubMed]
- Cebron, A.; Bodrossy, L.; Stralis-Pavese, N.; Singer, A.C.; Thompson, I.P.; Prosser, J.I.; Murrell, J.C. Nutrient amendments in soil DNA stable isotope probing experiments reduce the observed methanotroph diversity. Appl. Environ. Microbiol. 2007, 73, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Dedysh, S.N. Methanotrophic bacteria of acidic Sphagnum peat bogs. Microbiology 2002, 71, 638–650. [Google Scholar] [CrossRef]
- Henckel, T.; Roslev, P.; Conrad, R. Effects of O-2 and CH4 on presence and activity of the indigenous methanotrophic community in rice field soil. Environ. Microbiol. 2000, 2, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Hanna, M.L.; Taylor, R.T.; Droege, M.W. Batch cultivation of Methylosinus trichosporium OB3b.1. Production of soluble methane monooxygenase. Biotechnol. Bioeng. 1991, 38, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Kunz, R.C.; Boyd, E.S.; Semrau, J.D.; Antholine, W.E.; Han, J.I.; Zahn, J.A.; Boyd, J.M.; de la Mora, A.M.; DiSpirito, A.A. The membrane-associated methane monooxygenase (pMMO) and pMMO-NADH: Quinone oxidoreductase complex from Methylococcus capsulatus bath. J. Bacteriol. 2003, 185, 5755–5764. [Google Scholar] [CrossRef] [PubMed]
- Chidambarampadmavathy, K.; Obulisamy, P.K.; Heimann, K. Role of copper and iron in methane oxidation and bacterial biopolymer accumulation. Eng. Life Sci. 2015, 15, 387–399. [Google Scholar] [CrossRef]
- Gupta, G.; Bhardwaj, K.N.; Choudhary, C.; Chandna, P.; Jain, K.K.; Kaur, A.; Kumar, S.; Shrivastava, B.; Ninawe, S.; Singh, A.; Singh, A.; et al. Biofuels: The environment-friendly energy carriers. In Biotechnology for Environmental Management and Resource Recovery; Singh, A., Kuhad, R.C., Eds.; Springer India: New Delhi, India, 2013; pp. 125–148. [Google Scholar]
- Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiol. Rev. 2010, 34, 496–531. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga, C.; Morales, M.; Revah, S. Polyhydroxyalkanoates accumulation by Methylobacterium organophilum CZ-2 during methane degradation using citrate or propionate as cosubstrates. Bioresour. Technol. 2013, 129, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Siegel, M.H.; Robinson, C.W. Application of airlift gas-liquid-solid reactors in biotechnology. Chem. Eng. Sci. 1992, 47, 3215–3229. [Google Scholar] [CrossRef]
- Chisti, Y.; Kasper, M.; Moo-Young, M. Mass transfer in external-loop airlift bioreactors using static mixers. Can. J. Chem. Eng. 1990, 68, 45–50. [Google Scholar] [CrossRef]
- Newlight Technologies: From Greenhouse Gas to Plastic. Available online: http://newlight.com/ (accessed on 20 November 2015).
- Mango Materials. Available online: http://mangomaterials.com/ (accessed on 20 November 2015).
- Koller, M.; Niebelschutz, H.; Braunegg, G. Strategies for recovery and purification of poly[(R)-3-hydroxyalkanoates] (PHA) biopolyesters from surrounding biomass. Eng. Life Sci. 2013, 13, 549–562. [Google Scholar] [CrossRef]
- Chen, G.Q.; Patel, M.K. Plastics derived from biological sources: Present and future: A technical and environmental review. Chem. Rev. 2012, 112, 2082–2099. [Google Scholar] [CrossRef] [PubMed]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Myung, J. PHA copolymers from methane. In Proceedings of the International Conference and Exhibition on Biopolymers and Bioplastics, San Francisco, CA, USA, 10–12 August 2015.
- McChalicher, C.W.J.; Srienc, F. Investigating the structure-property relationship of bacterial PHA block copolymers. J. Biotechnol. 2007, 132, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M.G.; Lapidus, A.; Ivanova, N.; Copeland, A.C.; McHardy, A.C.; Szeto, E.; Salamov, A.; Grigoriev, I.V.; Suciu, D.; Levine, S.R.; et al. High-resolution metagenomics targets specific functional types in complex microbial communities. Nat. Biotechnol. 2008, 26, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Bothe, H.; Møller Jensen, K.; Mergel, A.; Larsen, J.; Jørgensen, C.; Bothe, H.; Jørgensen, L. Heterotrophic bacteria growing in association with Methylococcus capsulatus (Bath) in a single cell protein production process. Appl. Microbiol. Biotechnol. 2002, 59, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Slater, S.; Houmiel, K.L.; Tran, M.; Mitsky, T.A.; Taylor, N.B.; Padgette, S.R.; Gruys, K.J. Multiple b-ketothiolases mediate poly(b-hydroxyalkanoate) copolymer synthesis in Ralstonia eutropha. J. Bacteriol. 1998, 180, 1979–1987. [Google Scholar] [PubMed]
- Wang, Y.P.; Chen, R.H.; Cai, J.Y.; Liu, Z.G.; Zheng, Y.M.; Wang, H.T.; Li, Q.B.; He, N. Biosynthesis and thermal properties of PHBV produced from levulinic acid by Ralstonia eutropha. PLoS ONE 2013, 8, e60318. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.C.; Chen, J.Y.; Zhang, L.; Chen, G.Q. Polyhydroxyalkanoate copolyesters produced by Ralstonia eutropha PHB(−)4 harboring a low-substrate-specificity PHA synthase PhaC2(Ps) from Pseudomonas stutzeri 1317. Biochem. Eng. J. 2006, 32, 218–225. [Google Scholar] [CrossRef]
- Torres, M.G.; Talavera, J.R.R.; Munoz, S.V.; Carreon-Castro, M.D.; Munoz, E.M.R.; Padron, G.H.; Guerrero, F.Q. Crystalline and spectroscopic characterization of poly(2-aminoethyl methacrylate hydrochloride) chains grafted onto poly[(R)-3-hydroxybutyric acid]. Vib. Spectrosc. 2015, 76, 55–62. [Google Scholar] [CrossRef]
- Nguyen, S. Graft copolymers containing poly(3-hydroxyalkanoates)—A review on their synthesis, properties, and applications. Can. J. Chem. 2008, 86, 570–578. [Google Scholar] [CrossRef]
- Hazer, D.B.; Kilicay, E.; Hazer, B. Poly(3-hydroxyalkanoate)s: Diversification and biomedical applications, a state of the art review. Mat. Sci. Eng. 2012, 32, 637–647. [Google Scholar] [CrossRef]
- Pina, S.; Oliveira, J.M.; Reis, R.L. Natural-based nanocomposites for bone tissue engineering and regenerative medicine: A review. Adv. Mater. 2015, 27, 1143–1169. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Dean, K.; Li, L. Polymer blends and composites from renewable resources. Prog. Polym. Sci. 2006, 31, 576–602. [Google Scholar] [CrossRef]
- Orts, W.J.; Nobes, G.A.R.; Kawada, J.; Nguyen, S.; Yu, G.E.; Raveneile, F. Poly(hydroxyalkanoates): Biorefinery polymers with a whole range of applications. The work of Robert H. Marchessault. Can. J. Chem. 2008, 86, 628–640. [Google Scholar] [CrossRef]
- Nguyen, S.; Yu, G.E.; Marchessault, R.H. Thermal degradation of poly(3-hydroxyalkanoates): Preparation of well-defined oligomers. Biomacromolecules 2002, 3, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Saville, R.M.; Lee, S.; Regitsky, D.D.; Resnick, S.M.; Silverman, J. Compositions and Methods for Biological Production of Lactate From C1 Compounds Using Lactate Dehydrogenase Transformants. Patent WO 2014205146, 24 December 2014. [Google Scholar]
- Choi, D.; Chipman, D.C.; Bents, S.C.; Brown, R.C. A techno-economic analysis of polyhydroxyalkanoate and hydrogen production from syngas fermentation of gasified bimass. Appl. Biochem. Biotechnol. 2010, 160, 1032–1046. [Google Scholar] [CrossRef] [PubMed]
- Aragno, M.; Schlegel, H.G. The mesophilic hydrogen-oxidizing (knallgas) bacteria. In The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Balows, A.T., Truper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer-Verlag: Berlin, Germany, 1992; Volume 1, pp. 344–384. [Google Scholar]
- Sonnleitner, B.; Heinzle, E.; Braunegg, G.; Lafferty, R.M. Formal kinetics of poly-b-hydroxybutyric acid (PHB) production in Alcaligenes eutrophus H16 and Mycoplana rubra R14 with respect to the dissolved-oxygen tension in ammonium-limited batch cultures. Eur. J. Appl. Microbiol. 1979, 7, 1–10. [Google Scholar] [CrossRef]
- Panda, B.; Mallick, N. Enhanced poly-b-hydroxybutyrate accumulation in a unicellular cyanobacterium, Synechocystis sp PCC 6803. Lett. Appl. Microbiol. 2007, 44, 194–198. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strong, P.J.; Laycock, B.; Mahamud, S.N.S.; Jensen, P.D.; Lant, P.A.; Tyson, G.; Pratt, S. The Opportunity for High-Performance Biomaterials from Methane. Microorganisms 2016, 4, 11. https://doi.org/10.3390/microorganisms4010011
Strong PJ, Laycock B, Mahamud SNS, Jensen PD, Lant PA, Tyson G, Pratt S. The Opportunity for High-Performance Biomaterials from Methane. Microorganisms. 2016; 4(1):11. https://doi.org/10.3390/microorganisms4010011
Chicago/Turabian StyleStrong, Peter James, Bronwyn Laycock, Syarifah Nuraqmar Syed Mahamud, Paul Douglas Jensen, Paul Andrew Lant, Gene Tyson, and Steven Pratt. 2016. "The Opportunity for High-Performance Biomaterials from Methane" Microorganisms 4, no. 1: 11. https://doi.org/10.3390/microorganisms4010011