
Recombinant Production of Pseudomonas aeruginosa Rhamnolipids in P. putida KT2440 on Acetobacterium woodii Cultures Grown Chemo-Autotrophically with Carbon Dioxide and Hydrogen

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media and Cultivation
2.2. Bacterial Strains and Plasmids
2.3. Optical Density (OD600) Measurement
2.4. Quantification of Mono-Rhamnolipids
2.5. Gas Chromatography (GC)
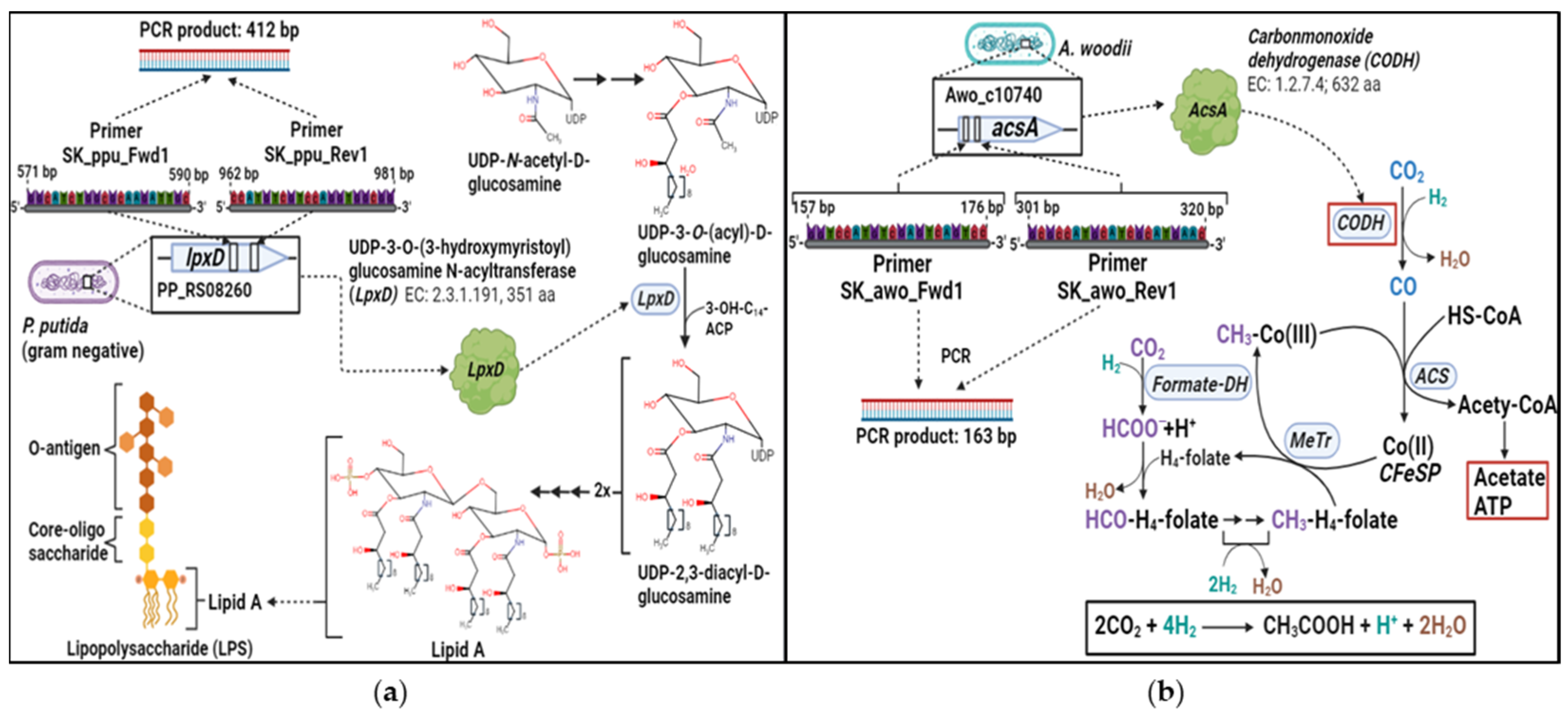
2.6. Primers and qPCR
2.7. Preservation of the Bacterial Strains
3. Results
3.1. Cultivation of the P. putida Strains Using Acetate as a Substrate
3.2. Production of Mono-Rhamnolipids by P. putida [pVLT31_rhlAB] in Cell-Free A. woodii [pMTL83251] Spent Medium
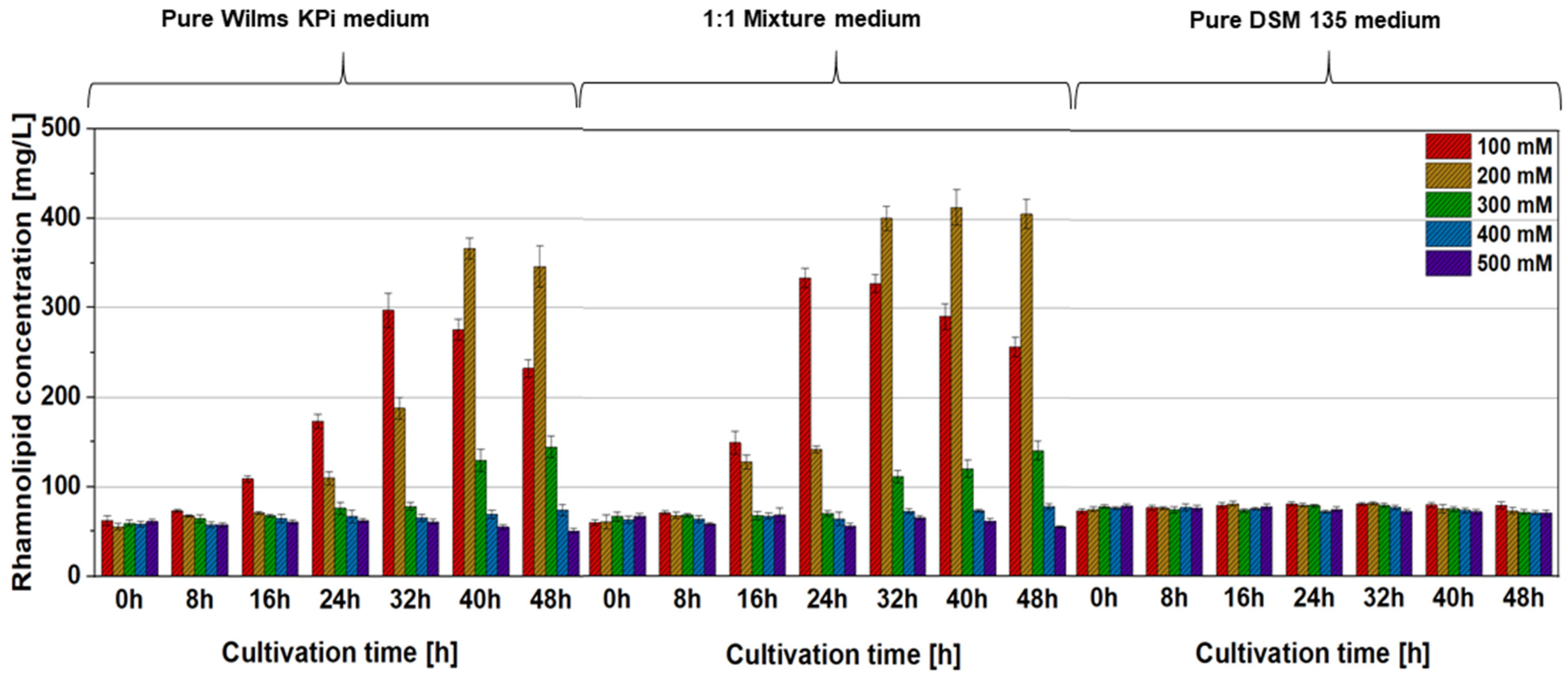
3.3. Production of Mono-Rhamnolipids by P. putida [pVLT31_rhlAB] in Cell-Containing A. woodii [pMTL83251] Spent Medium (Culture Broth)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nikolova, C.; Gutierrez, T. Biosurfactants and Their Applications in the Oil and Gas Industry: Current State of Knowledge and Future Perspectives. Front. Bioeng. Biotechnol. 2021, 9, 626639. [Google Scholar] [CrossRef] [PubMed]
- Nagtode, V.S.; Cardoza, C.; Yasin, H.K.A.; Mali, S.N.; Tambe, S.M.; Roy, P.; Singh, K.; Goel, A.; Amin, P.D.; Thorat, B.R.; et al. Green Surfactants (Biosurfactants): A Petroleum-Free Substitute for Sustainability—Comparison, Applications, Market, and Future Prospects. ACS Omega 2023, 8, 11674–11699. [Google Scholar] [CrossRef] [PubMed]
- Ceresa, C.; Fracchia, L.; Sansotera, A.C.; De Rienzo, M.A.D.; Banat, I.M. Harnessing the Potential of Biosurfactants for Biomedical and Pharmaceutical Applications. Pharmaceutics 2023, 15, 2156. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.F.; Banat, I.M.; Giachini, A.J.; Robl, D. Fungal Biosurfactants, from Nature to Biotechnological Product: Bioprospection, Production and Potential Applications. Bioprocess Biosyst. Eng. 2021, 44, 2003–2034. [Google Scholar] [CrossRef] [PubMed]
- Eras-Muñoz, E.; Farré, A.; Sánchez, A.; Font, X.; Gea, T. Microbial Biosurfactants: A Review of Recent Environmental Applications. Bioengineered 2022, 13, 12365–12391. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, M.; Costa, S.G.V.A.O.; Contiero, J. Rhamnolipid Surfactants: An Update on the General Aspects of These Remarkable Biomolecules. Biotechnol. Prog. 2005, 21, 1593–1600. [Google Scholar] [CrossRef]
- Wittgens, A.; Tiso, T.; Arndt, T.T.; Wenk, P.; Hemmerich, J.; Müller, C.; Wichmann, R.; Küpper, B.; Zwick, M.; Wilhelm, S.; et al. Growth Independent Rhamnolipid Production from Glucose Using the Non-Pathogenic Pseudomonas putida KT2440. Microb. Cell Fact. 2011, 10, 80. [Google Scholar] [CrossRef]
- Lang, S.; Wullbrandt, D. Rhamnose Lipids—Biosynthesis, Microbial Production and Application Potential. Appl. Microbiol. Biotechnol. 1999, 51, 22–32. [Google Scholar] [CrossRef]
- Maier, R.M.; Soberón-Chávez, G. Pseudomonas aeruginosa Rhamnolipids: Biosynthesis and Potential Applications. Appl. Microbiol. Biotechnol. 2000, 54, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Irorere, V.U.; Tripathi, L.; Marchant, R.; McClean, S.; Banat, I.M. Microbial Rhamnolipid Production: A Critical Re-Evaluation of Published Data and Suggested Future Publication Criteria. Appl. Microbiol. Biotechnol. 2017, 101, 3941–3951. [Google Scholar] [CrossRef] [PubMed]
- Fracchia, L.; Ceresa, C.; Franzetti, A.; Cavallo, M.; Gandolfi, I.; Van Hamme, J.; Gkorezis, P.; Marchant, R.; Banat, I.M. Biological Applications of Biosurfactants and Strategies to Potentiate Commercial Production. In Biosurfactants; CRC Press: Boca Raton, FL, USA, 2014; pp. 280–305. [Google Scholar]
- Gautam, K.K.; Tyagi, V.K. Microbial Surfactants: A Review. J. Oleo Sci. 2006, 55, 155–166. [Google Scholar] [CrossRef]
- Rahman, P.K.S.M.; Gakpe, E. Production, Characterisation and Applications of Biosurfactants—Review. Biotechnology 2008, 7, 360–370. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial Production of Surfactants and Their Commercial Potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Abalos, A.; Pinazo, A.; Infante, M.R.; Casals, M.; García, F.; Manresa, A. Physicochemical and Antimicrobial Properties of New Rhamnolipids Produced by Pseudomonas aeruginosa AT10 from Soybean Oil Refinery Wastes. Langmuir 2001, 17, 1367–1371. [Google Scholar] [CrossRef]
- Soberón-Chávez, G.; Lépine, F.; Déziel, E. Production of Rhamnolipids by Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2005, 68, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Déziel, E.; Lépine, F.; Dennie, D.; Boismenu, D.; Mamer, O.A.; Villemur, R. Liquid Chromatography/Mass Spectrometry Analysis of Mixtures of Rhamnolipids Produced by Pseudomonas aeruginosa Strain 57RP Grown on Mannitol or Naphthalene. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 1999, 1440, 244–252. [Google Scholar] [CrossRef]
- Toribio, J.; Escalante, A.E.; Soberón-Chávez, G. Rhamnolipids: Production in Bacteria Other than Pseudomonas aeruginosa. Eur. J. Lipid Sci. Technol. 2010, 112, 1082–1087. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of Structures, Microbial Origins and Roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef]
- Jarvis, F.G.; Johnson, M.J. A Glycolipide Produced by Pseudomonas aeruginosa. J. Am. Chem. Soc. 1949, 71, 4124–4126. [Google Scholar] [CrossRef]
- Shreve, G.S.; Makula, R. Characterization of a New Rhamnolipid Biosurfactant Complex from Pseudomonas Isolate DYNA270. Biomolecules 2019, 9, 885. [Google Scholar] [CrossRef]
- Shi, S.; Teng, Z.; Liu, J.; Li, T. Conversion of Waste Cooking Oil to Rhamnolipid by a Newly Oleophylic Pseudomonas aeruginosa WO2. Int. J. Environ. Res. Public Health 2022, 19, 1700. [Google Scholar] [CrossRef]
- Chong, H.; Li, Q. Microbial Production of Rhamnolipids: Opportunities, Challenges and Strategies. Microb. Cell Fact. 2017, 16, 137. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Lin, J.; Wang, W.; Li, S. High-Yield Di-Rhamnolipid Production by Pseudomonas aeruginosa YM4 and Its Potential Application in MEOR. Molecules 2019, 24, 1433. [Google Scholar] [CrossRef]
- Soberón-Chávez, G.; González-Valdez, A.; Soto-Aceves, M.P.; Cocotl-Yañez, M. Rhamnolipids Produced by Pseudomonas: From Molecular Genetics to the Market. Microb. Biotechnol. 2021, 14, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Dulcey, C.E.; López de los Santos, Y.; Létourneau, M.; Déziel, E.; Doucet, N. Semi-rational Evolution of the 3-(3-hydroxyalkanoyloxy)Alkanoate (HAA) Synthase RhlA to Improve Rhamnolipid Production in Pseudomonas aeruginosa and Burkholderia glumae. FEBS J. 2019, 286, 4036–4059. [Google Scholar] [CrossRef] [PubMed]
- Häuβler, S.; Rohde, M.; von Neuhoff, N.; Nimtz, M.; Steinmetz, I. Structural and Functional Cellular Changes Induced by Burkholderia Pseudomallei Rhamnolipid. Infect. Immun. 2003, 71, 2970–2975. [Google Scholar] [CrossRef] [PubMed]
- Bzdyl, N.M.; Moran, C.L.; Bendo, J.; Sarkar-Tyson, M. Pathogenicity and Virulence of Burkholderia Pseudomallei. Virulence 2022, 13, 1945–1965. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Mannaa, M.; Kim, J.; Ra, J.-E.; Kim, S.-M.; Lee, C.; Lee, H.-H.; Seo, Y.-S. The In Vitro and in Planta Interspecies Interactions among Rice-Pathogenic Burkholderia Species. Plant Dis. 2021, 105, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.G.V.A.O.; Déziel, E.; Lépine, F. Characterization of Rhamnolipid Production by Burkholderia glumae. Lett. Appl. Microbiol. 2011, 53, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Ortega, L.; Rojas, C.M. Bacterial Panicle Blight and Burkholderia glumae: From Pathogen Biology to Disease Control. Phytopathology 2021, 111, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Fiechter, A.; Reiser, J. Isolation, Characterization, and Expression in Escherichia coli of the Pseudomonas aeruginosa RhlAB Genes Encoding a Rhamnosyltransferase Involved in Rhamnolipid Biosurfactant Synthesis. J. Biol. Chem. 1994, 269, 19787–19795. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa Las and Rhl Quorum-Sensing Systems in Control of Elastase and Rhamnolipid Biosynthesis Genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef] [PubMed]
- Nickzad, A.; Lépine, F.; Déziel, E. Quorum Sensing Controls Swarming Motility of Burkholderia glumae through Regulation of Rhamnolipids. PLoS ONE 2015, 10, e0128509. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.M.; Hausmann, R. Regulatory and Metabolic Network of Rhamnolipid Biosynthesis: Traditional and Advanced Engineering towards Biotechnological Production. Appl. Microbiol. Biotechnol. 2011, 91, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Reiser, J.; Fiechter, A.; Witholt, B. Production of Pseudomonas aeruginosa Rhamnolipid Biosurfactants in Heterologous Hosts. Appl. Environ. Microbiol. 1995, 61, 3503–3506. [Google Scholar] [CrossRef]
- Nelson, K.E.; Weinel, C.; Paulsen, I.T.; Dodson, R.J.; Hilbert, H.; Martins dos Santos, V.A.P.; Fouts, D.E.; Gill, S.R.; Pop, M.; Holmes, M.; et al. Complete Genome Sequence and Comparative Analysis of the Metabolically Versatile Pseudomonas putida KT2440. Environ. Microbiol. 2002, 4, 799–808. [Google Scholar] [CrossRef]
- Tiso, T.; Wierckx, N.; Blank, L.M. Non-Pathogenic Pseudomonas as Platform for Industrial Biocatalysts; Grunwald, P., Ed.; Jenny Stanford Publishing: Dubai, United Arab Emirates, 2014; ISBN 9780429090264. [Google Scholar]
- Wang, Y.; Horlamus, F.; Henkel, M.; Kovacic, F.; Schläfle, S.; Hausmann, R.; Wittgens, A.; Rosenau, F. Growth of Engineered Pseudomonas putida KT2440 on Glucose, Xylose, and Arabinose: Hemicellulose Hydrolysates and Their Major Sugars as Sustainable Carbon Sources. GCB Bioenergy 2019, 11, 249–259. [Google Scholar] [CrossRef]
- Arnold, S.; Henkel, M.; Wanger, J.; Wittgens, A.; Rosenau, F.; Hausmann, R. Heterologous Rhamnolipid Biosynthesis by P. putida KT2440 on Bio-Oil Derived Small Organic Acids and Fractions. AMB Express 2019, 9, 80. [Google Scholar] [CrossRef]
- Drake, H.L.; Daniel, S.L.; Matthies, C.; Küsel, K. Acetogenesis, Acetogenic Bacteria, and the Acetyl-CoA “Wood/Ljungdahl” Pathway: Past and Current Perspectives. In Acetogenesis; Springer: Boston, MA, USA, 1994; pp. 3–60. [Google Scholar]
- Drake, H.L.; Gößner, A.S.; Daniel, S.L. Old Acetogens, New Light. Ann. N. Y. Acad. Sci. 2008, 1125, 100–128. [Google Scholar] [CrossRef]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood–Ljungdahl Pathway of CO2 Fixation. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2008, 1784, 1873–1898. [Google Scholar] [CrossRef]
- Bengelsdorf, F.R.; Dürre, P. Gas Fermentation for Commodity Chemicals and Fuels. Microb. Biotechnol. 2017, 10, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Müller, V. Energy Conservation in Acetogenic Bacteria. Appl. Environ. Microbiol. 2003, 69, 6345–6353. [Google Scholar] [CrossRef] [PubMed]
- Poehlein, A.; Schmidt, S.; Kaster, A.-K.; Goenrich, M.; Vollmers, J.; Thürmer, A.; Bertsch, J.; Schuchmann, K.; Voigt, B.; Hecker, M.; et al. An Ancient Pathway Combining Carbon Dioxide Fixation with the Generation and Utilization of a Sodium Ion Gradient for ATP Synthesis. PLoS ONE 2012, 7, e33439. [Google Scholar] [CrossRef] [PubMed]
- Abubackar, H.N.; Bengelsdorf, F.R.; Dürre, P.; Veiga, M.C.; Kennes, C. Improved Operating Strategy for Continuous Fermentation of Carbon Monoxide to Fuel-Ethanol by Clostridia. Appl. Energy 2016, 169, 210–217. [Google Scholar] [CrossRef]
- Phillips, J.R.; Atiyeh, H.K.; Tanner, R.S.; Torres, J.R.; Saxena, J.; Wilkins, M.R.; Huhnke, R.L. Butanol and Hexanol Production in Clostridium Carboxidivorans Syngas Fermentation: Medium Development and Culture Techniques. Bioresour. Technol. 2015, 190, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ramió-Pujol, S.; Ganigué, R.; Bañeras, L.; Colprim, J. Incubation at 25 °C Prevents Acid Crash and Enhances Alcohol Production in Clostridium Carboxidivorans P7. Bioresour. Technol. 2015, 192, 296–303. [Google Scholar] [CrossRef]
- Hoffmeister, S.; Gerdom, M.; Bengelsdorf, F.R.; Linder, S.; Flüchter, S.; Öztürk, H.; Blümke, W.; May, A.; Fischer, R.-J.; Bahl, H.; et al. Acetone Production with Metabolically Engineered Strains of Acetobacterium woodii. Metab. Eng. 2016, 36, 37–47. [Google Scholar] [CrossRef]
- Mook, A.; Beck, M.H.; Baker, J.P.; Minton, N.P.; Dürre, P.; Bengelsdorf, F.R. Autotrophic Lactate Production from H2 + CO2 Using Recombinant and Fluorescent FAST-Tagged Acetobacterium woodii Strains. Appl. Microbiol. Biotechnol. 2022, 106, 1447–1458. [Google Scholar] [CrossRef]
- Geinitz, B.; Hüser, A.; Mann, M.; Büchs, J. Gas Fermentation Expands the Scope of a Process Network for Material Conversion. Chem. Ing. Tech. 2020, 92, 1665–1679. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Autotrophy at the Thermodynamic Limit of Life: A Model for Energy Conservation in Acetogenic Bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef]
- Arslan, K.; Schoch, T.; Höfele, F.; Herrschaft, S.; Oberlies, C.; Bengelsdorf, F.; Veiga, M.C.; Dürre, P.; Kennes, C. Engineering Acetobacterium woodii for the Production of Isopropanol and Acetone from Carbon Dioxide and Hydrogen. Biotechnol. J. 2022, 17, 2100515. [Google Scholar] [CrossRef]
- Bertani, G. Studies on Lysogenesis. I. The Mode of Phage Liberation by Lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef]
- Tschech, A.; Pfennig, N. Growth Yield Increase Linked to Caffeate Reduction in Acetobacterium woodii. Arch. Microbiol. 1984, 137, 163–167. [Google Scholar] [CrossRef]
- Wolin, E.A.; Wolin, M.J.; Wolfe, R.S. Formation of Methane by Bacterial Extracts. J. Biol. Chem. 1963, 238, 2882–2886. [Google Scholar] [CrossRef] [PubMed]
- Wilms, B.; Hauck, A.; Reuss, M.; Syldatk, C.; Mattes, R.; Siemann, M.; Altenbuchner, J. High-cell-density Fermentation for Production of L-carbamoylase Using an Expression System Based on the Escherichia coli RhaBAD Promoter. Biotechnol. Bioeng. 2001, 73, 95–103. [Google Scholar] [CrossRef] [PubMed]
- de Lorenzo, V.; Eltis, L.; Kessler, B.; Timmis, K.N. Analysis of Pseudomonas Gene Products Using LacIq/Ptrp-Lac Plasmids and Transposons That Confer Conditional Phenotypes. Gene 1993, 123, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Minton, N.P. A Modular System for Clostridium Shuttle Plasmids. J. Microbiol. Methods 2009, 78, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Song, Y.; Jeong, Y.; Cho, B.-K. Analysis of the Core Genome and Pan-Genome of Autotrophic Acetogenic Bacteria. Front. Microbiol. 2016, 7, 1531. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W. Nickel and the Carbon Cycle. J. Inorg. Biochem. 2007, 101, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Buetow, L.; Smith, T.K.; Dawson, A.; Fyffe, S.; Hunter, W.N. Structure and Reactivity of LpxD, the N-Acyltransferase of Lipid A Biosynthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 4321–4326. [Google Scholar] [CrossRef]
- Stark, C.; Münßinger, S.; Rosenau, F.; Eikmanns, B.J.; Schwentner, A. The Potential of Sequential Fermentations in Converting C1 Substrates to Higher-Value Products. Front. Microbiol. 2022, 13, 907577. [Google Scholar] [CrossRef]
- Ljungdahl, L.G.; Andreesen, J.R. Formate Dehydrogenase, a Selenium-Tungsten Enzyme from Clostridium thermoaceticum. Methods Enzymol. 1978, 53, 360–372. [Google Scholar]
- Martin, W.; Russell, M.J. On the Origin of Biochemistry at an Alkaline Hydrothermal Vent. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1887–1926. [Google Scholar] [CrossRef]
- Popov, A.L.; Zholobak, N.M.; Balko, O.I.; Balko, O.B.; Shcherbakov, A.B.; Popova, N.R.; Ivanova, O.S.; Baranchikov, A.E.; Ivanov, V.K. Photo-Induced Toxicity of Tungsten Oxide Photochromic Nanoparticles. J. Photochem. Photobiol. B 2018, 178, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Matharu, R.K.; Ciric, L.; Ren, G.; Edirisinghe, M. Comparative Study of the Antimicrobial Effects of Tungsten Nanoparticles and Tungsten Nanocomposite Fibres on Hospital Acquired Bacterial and Viral Pathogens. Nanomaterials 2020, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Strigul, N.; Koutsospyros, A.; Arienti, P.; Christodoulatos, C.; Dermatas, D.; Braida, W. Effects of Tungsten on Environmental Systems. Chemosphere 2005, 61, 248–258. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Description | Source |
|---|---|---|
| pVLT31 | Expression vector with the IPTG-inducible tac promoter, TetR | De Lorenzo et al., (1993) [59] |
| pVLT31_rhlAB | Same as pVLT31, rhlAB genes of P. aeruginosa PAO1 under the control of the tac promoter | Wittgens et al., (2011) [7] |
| pMTL83251 | The backbone for homologous recombination vectors, CmR | Heap et al., (2009) [60] |
| Primer | Sequence (5′ → 3′) |
|---|---|
| SK_awo_Fwd1 | GGTCCATGTCGAGTCAGTCC |
| SK_awo_Rev1 | GCGCCATGCATCGCATGAAC |
| SK_ppu_Fwd1 | GGCATCTGGCGCAAGATTGC |
| SK_ppu_Rev1 | CCATGTCGTCCAGGTGGCGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widberger, J.; Wittgens, A.; Klaunig, S.; Krämer, M.; Kissmann, A.-K.; Höfele, F.; Baur, T.; Weil, T.; Henkel, M.; Hausmann, R.; et al. Recombinant Production of Pseudomonas aeruginosa Rhamnolipids in P. putida KT2440 on Acetobacterium woodii Cultures Grown Chemo-Autotrophically with Carbon Dioxide and Hydrogen. Microorganisms 2024, 12, 529. https://doi.org/10.3390/microorganisms12030529
Widberger J, Wittgens A, Klaunig S, Krämer M, Kissmann A-K, Höfele F, Baur T, Weil T, Henkel M, Hausmann R, et al. Recombinant Production of Pseudomonas aeruginosa Rhamnolipids in P. putida KT2440 on Acetobacterium woodii Cultures Grown Chemo-Autotrophically with Carbon Dioxide and Hydrogen. Microorganisms. 2024; 12(3):529. https://doi.org/10.3390/microorganisms12030529
Chicago/Turabian StyleWidberger, Jonas, Andreas Wittgens, Sebastian Klaunig, Markus Krämer, Ann-Kathrin Kissmann, Franziska Höfele, Tina Baur, Tanja Weil, Marius Henkel, Rudolf Hausmann, and et al. 2024. "Recombinant Production of Pseudomonas aeruginosa Rhamnolipids in P. putida KT2440 on Acetobacterium woodii Cultures Grown Chemo-Autotrophically with Carbon Dioxide and Hydrogen" Microorganisms 12, no. 3: 529. https://doi.org/10.3390/microorganisms12030529