

The Effects of Silver Nanoparticles (AgNPs) on Thermophilic Bacteria: Antibacterial, Morphological, Physiological and Biochemical Investigations

Abstract
:1. Introduction
2. Materials and Methods
2.1. Silver Nanoparticles (AgNPs)
2.2. Determination of Silver (Ag) Ion Release from AgNPs
2.3. Strains and Maintenance of Cultures
2.4. Susceptibility of Bacterial Strains to Ag Nanoparticles
2.5. Exploring Cellular Damage through Transmission Electron Microscopy (TEM)
2.6. FT-IR Analysis of Bacterial Biomass Treated with Nanoparticles
2.7. Evaluating Cellular Viability in the Presence of NP Stress
2.8. Quantifying Superoxide Generation in Bacterial Cells under NP-Induced Stress
2.9. Effect of Silver NPs on Enzyme Biosynthesis
2.10. Effect of Silver NPs on Enzyme Secretion and Inhibition
2.11. Statistical Analysis
3. Results and Discussion
3.1. Determination of Silver (Ag) Ion Release from AgNPs
3.2. Phenotypic Characterization of Bacterial Strains
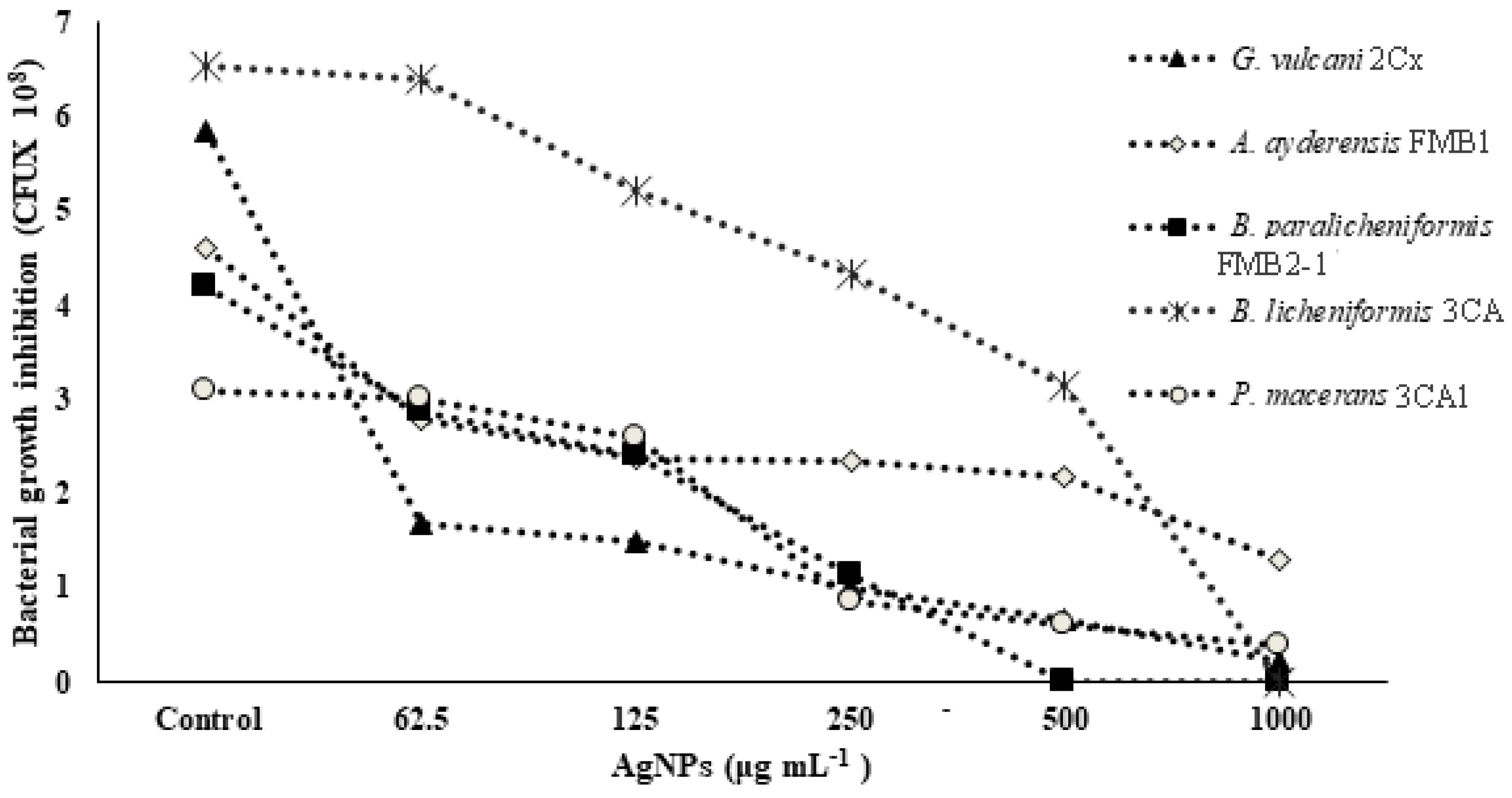
3.3. Bacterial Growth and NP Tolerance/Sensitivity
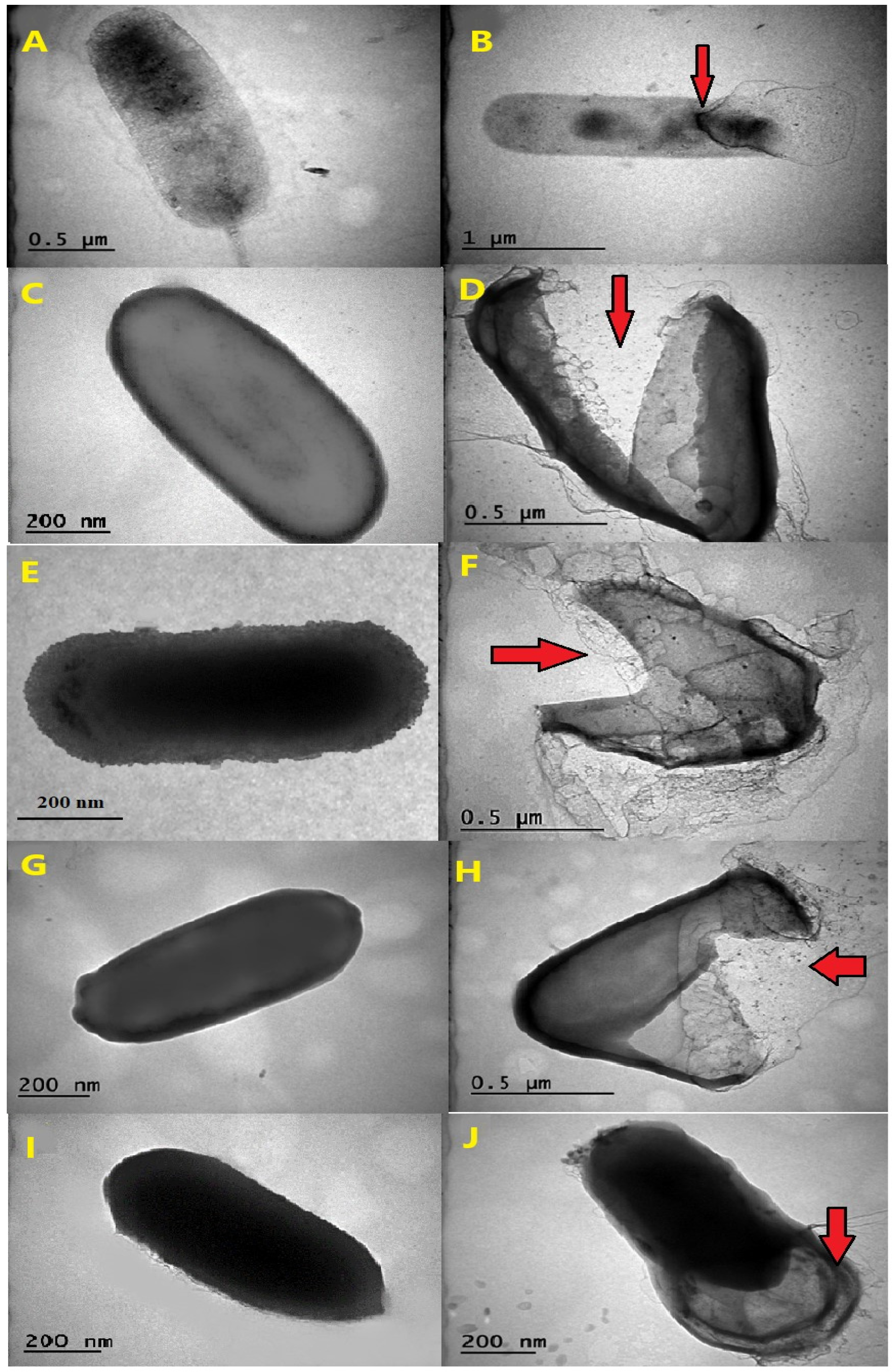
3.4. Exploring Cellular Damage through Transmission Electron Microscopy (TEM)
3.5. Analysis of AgNP-Treated Bacterial Biomass Using FT-IR
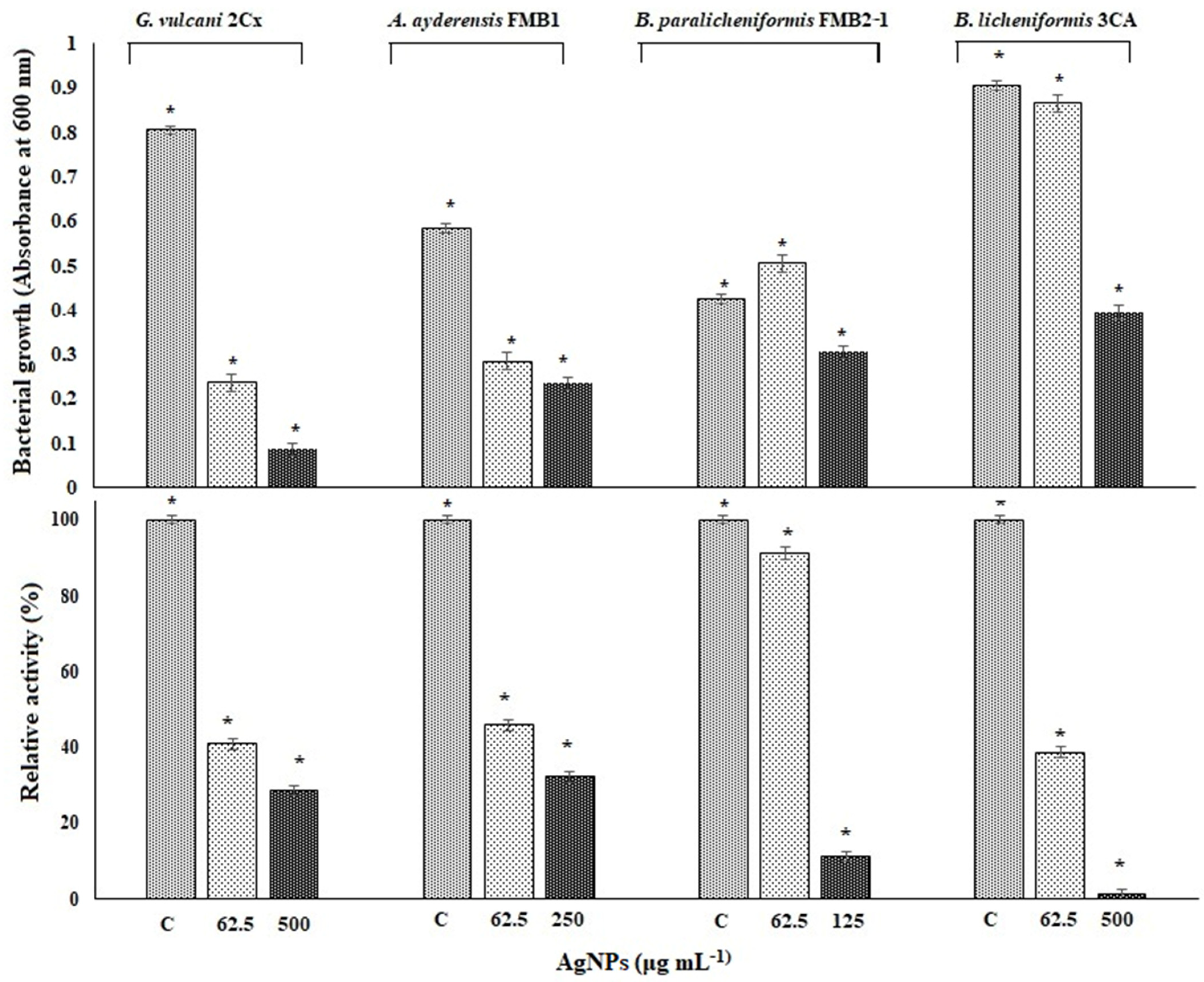
3.6. Evaluating Cellular Viability in the Presence of AgNP Stress
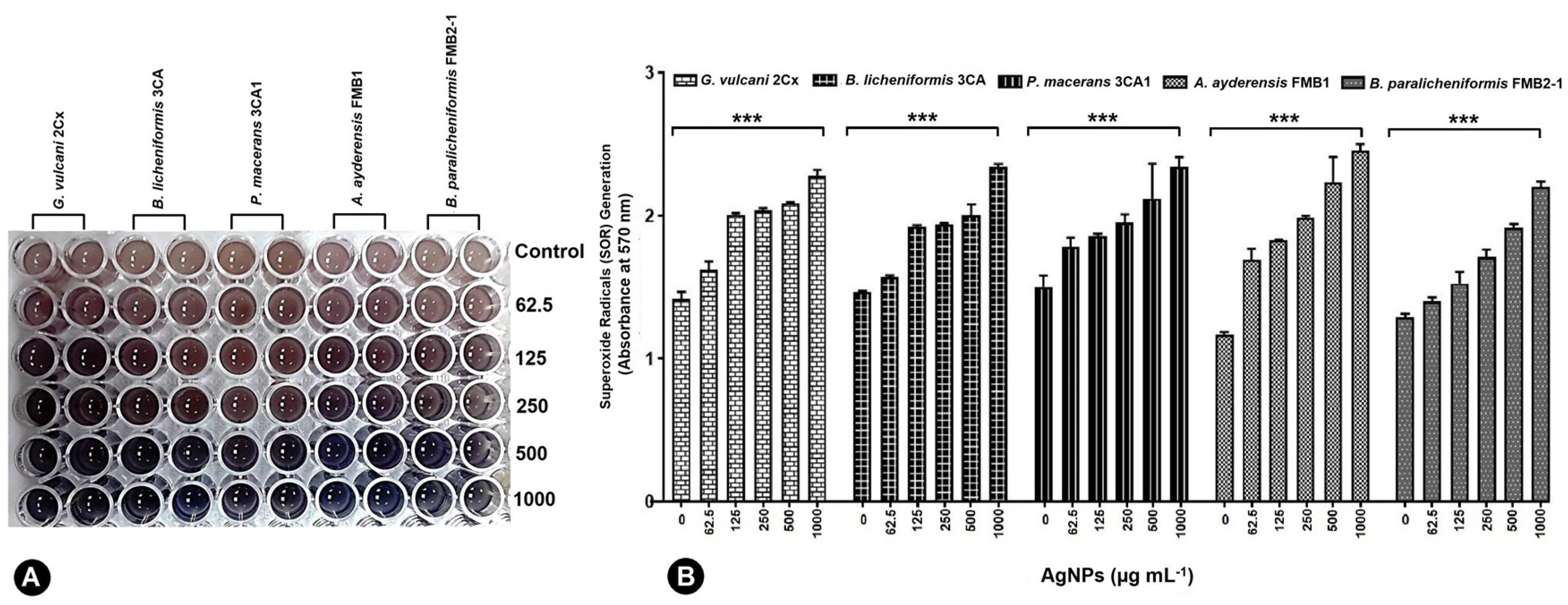
3.7. Quantifying Superoxide Generation in Bacterial Cells under AgNP-Induced Stress
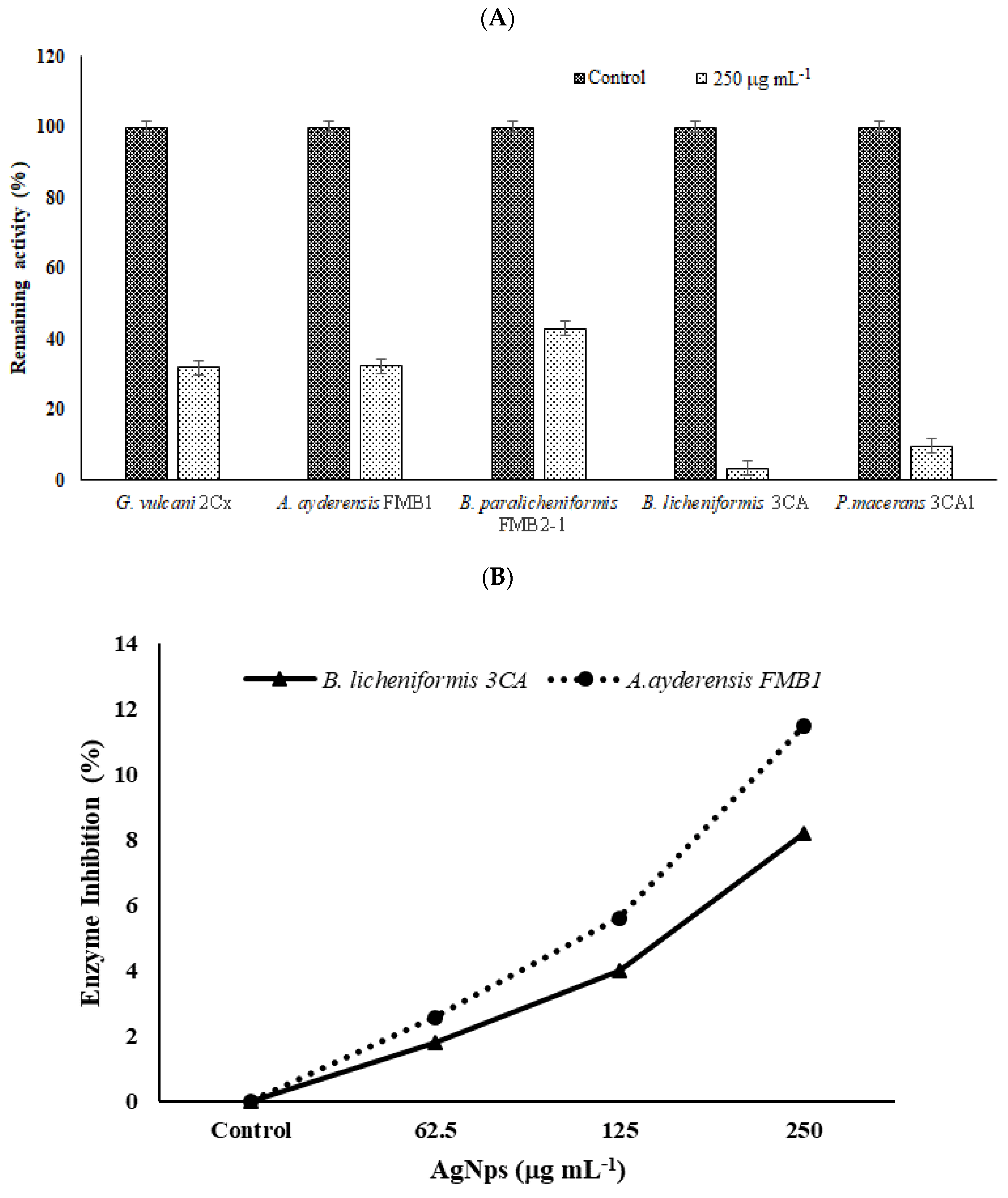
3.8. Effect of Silver NPs on Enzyme Biosynthesis
3.9. Effect of Silver NPs on Enzyme Secretion and Inhibition
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nafisi, S.; Maibach, H.I. Nanotechnology in Cosmetics. In Cosmetic Science and Technology: Theoretical Principles and Applications; Elsevier: Amsterdam, The Netherlands, 2017; pp. 337–369. [Google Scholar] [CrossRef]
- Keller, A.A.; Vosti, W.; Wang, H.; Lazareva, A. Release of Engineered Nanomaterials from Personal Care Products throughout Their Life Cycle. J. Nanopart. Res. 2014, 16, 2489. [Google Scholar] [CrossRef]
- Pourzahedi, L.; Vance, M.; Eckelman, M.J. Life cycle assessment and release studies for 15 nanosilver-enabled consumer products: Investigating hotspots and patterns of contribution. Environ. Sci. Technol. 2017, 51, 7148–7158. [Google Scholar] [CrossRef]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella, M.F.; Rejeski, D.; Hull, M.S. Nanotechnology in the Real World: Redeveloping the Nanomaterial Consumer Products Inventory. Beilstein J. Nanotechnol. 2015, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Forge, D.; Port, M.; Roch, A.; Robic, C.; Vander Elst, L.; Muller, R.N. Magnetic Iron Oxide Nanoparticles: Synthesis, Stabilization, Vectorization, Physicochemical Characterizations, and Biological Applications. Chem. Rev. 2010, 110, 2574. [Google Scholar] [CrossRef]
- Sudha, P.N.; Sangeetha, K.; Vijayalakshmi, K.; Barhoum, A. Nanomaterials History, Classification, Unique Properties, Production and Market. In Emerging Applications of Nanoparticles and Architecture Nanostructures; Elsevier: Amsterdam, The Netherlands, 2018; pp. 341–384. [Google Scholar] [CrossRef]
- Martis, E.; Badve, R.; Degwekar, M. Nanotechnology Based Devices and Applications in Medicine: An Overview. Chron. Young Sci. 2012, 3, 68. [Google Scholar] [CrossRef]
- Stampoulis, D.; Sinha, S.K.; White, J.C. Assay-Dependent Phytotoxicity of Nanoparticles to Plants. Environ. Sci. Technol. 2009, 43, 9473–9479. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-R.; Xie, X.-B.; Shi, Q.-S.; Zeng, H.-Y.; OU-Yang, Y.-S.; Chen, Y.-B. Antibacterial Activity and Mechanism of Silver Nanoparticles on Escherichia Coli. Appl. Microbiol. Biotechnol. 2009, 85, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Liu, Z.-G.; Shen, W.; Gurunathan, S. Silver Nanoparticles: Synthesis, Characterization, Properties, Applications, and Therapeutic Approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef]
- Tran, Q.H.; Nguyen, V.Q.; Le, A.-T. Silver Nanoparticles: Synthesis, Properties, Toxicology, Applications and Perspectives. Adv. Nat. Sci. Nanosci. Nanotechnol. 2013, 4, 033001. [Google Scholar] [CrossRef]
- Nie, P.; Zhao, Y.; Xu, H. Synthesis, Applications, Toxicity and Toxicity Mechanisms of Silver Nanoparticles: A Review. Ecotoxicol. Environ. Saf. 2023, 253, 114636. [Google Scholar] [CrossRef]
- Ramalingam, B.; Parandhaman, T.; Das, S.K. Antibacterial effects of biosynthesized silver nanoparticles on surface ultrastructure and nanomechanical properties of gram-negative bacteria viz. Escherichia coli and Pseudomonas aeruginosa. ACS Appl. Mater. Interfaces 2016, 8, 4963–4976. [Google Scholar] [CrossRef]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The Bactericidal Effect of Silver Nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Nunez, N.V.; Villegas, H.H.L.; Turrent, L.C.I.; Padilla, C.R. Silver nanoparticles toxicity and bactericidal effect against methicillin resistant Staphylococcus aureus: Nanoscale does matter. J. Nanobiotechnol. 2009, 5, 2–9. [Google Scholar] [CrossRef]
- Yacamán, M.J.; Ascencio, J.A.; Liu, H.B.; Gardea-Torresdey, J. Structure Shape and Stability of Nanometric Sized Particles. J. Vac. Sci. Technol. 2001, 19, 1091. [Google Scholar] [CrossRef]
- Gao, M.J.; Sun, L.; Wang, Z.Q.; Zhao, Y.B. Controlled synthesis of Ag nanoparticles with different morphologies and their antibacterial properties. Mater. Sci. Eng. C Mater. 2013, 33, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Wen, J.; Xiong, X.; Hu, Y. Shape effect on the antibacterial activity of silver nanoparticles synthesized via a microwave-assisted method. Environ. Sci. Pollut. Res. 2016, 23, 4489–4497. [Google Scholar] [CrossRef]
- Zheng, X.; Huang, H.; Su, Y.; Wei, Y.; Chen, Y. Long-Term Effects of Engineered Nanoparticles on Enzyme Activity and Functional Bacteria in Wastewater Treatment Plants. Water Sci. Technol. 2015, 72, 99–105. [Google Scholar] [CrossRef]
- He, S.; Feng, Y.; Ni, J.; Sun, Y.; Xue, L.; Feng, Y.; Yu, Y.; Lin, X.; Yang, L. Different Responses of Soil Microbial Metabolic Activity to Silver and Iron Oxide Nanoparticles. Chemosphere 2016, 147, 195–202. [Google Scholar] [CrossRef]
- Wang, J.; Shu, K.; Zhang, L.; Si, Y. Effects of Silver Nanoparticles on Soil Microbial Communities and Bacterial Nitrification in Suburban Vegetable Soils. Pedosphere 2017, 27, 482–490. [Google Scholar] [CrossRef]
- Chavan, S.; Nadanathangam, V. Effects of Nanoparticles on Plant Growth-Promoting Bacteria in Indian Agricultural Soil. Agronomy 2019, 9, 140. [Google Scholar] [CrossRef]
- Courtois, P.; Rorat, A.; Lemiere, S.; Guyoneaud, R.; Attard, E.; Levard, C.; Vandenbulcke, F. Ecotoxicology of Silver Nanoparticles and Their Derivatives Introduced in Soil with or without Sewage Sludge: A Review of Effects on Microorganisms, Plants and Animals. Environ. Pollut. 2019, 253, 578–598. [Google Scholar] [CrossRef]
- Wu, L.; Zhu, G.; Zhang, X.; Si, Y. Silver Nanoparticles Inhibit Denitrification by Altering the Viability and Metabolic Activity of Pseudomonas stutzeri. Sci. Total Environ. 2020, 706, 135711. [Google Scholar] [CrossRef] [PubMed]
- Mujeebur, R.K.; Fromm, K.M.; Tanveer, F.R.; Giese, B.; Ahamad, F.; Turner, R.J.; Füeg, M.; Marsili, E. Metal Nanoparticle–Microbe Interactions: Synthesis and Antimicrobial Effects. Part. Part. Syst. Charact. 2020, 37, 1900419. [Google Scholar] [CrossRef]
- Wang, L.; Hu, C.; Shao, L. The Antimicrobial Activity of Nanoparticles: Present Situation and Prospects for the Future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed]
- Rahmatpour, S.; Shirvani, M.; Mosaddeghi, M.R.; Nourbakhsh, F.; Bazarganipour, M. Dose–Response Effects of Silver Nanoparticles and Silver Nitrate on Microbial and Enzyme Activities in Calcareous Soils. Geoderma 2017, 285, 313–322. [Google Scholar] [CrossRef]
- Awet, T.T.; Kohl, Y.; Meier, F.; Straskraba, S.; Grün, A.-L.; Ruf, T.; Jost, C.; Drexel, R.; Tunc, E.; Emmerling, C. Effects of Polystyrene Nanoparticles on the Microbiota and Functional Diversity of Enzymes in Soil. Environ. Sci. Eur. 2018, 30, 11. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Li, X.; Yang, K.; Weijun, X.; Wen, Q.; Zhang, Y.; Lihui, Y.; Weng, L.; Teng, Z.; Wang, L. Efficient Bacteria Killing by Cu2WS4 Nanocrystals with Enzyme-like Properties and Bacteria-Binding Ability. ACS Nano 2019, 13, 13797–13808. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, L.S.; Kovel, E.S.; Kydralieva, K.A.; Dzhardimalieva, G.I.; Illés, E.; Tombácz, E.; Kicheeva, A.G.; Kudryasheva, N.S. Effects of Modified Magnetite Nanoparticles on Bacterial Cells and Enzyme Reactions. Nanomaterials 2020, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Huang, J.; Cao, C.; Li, R.; Ma, Y.; Wang, Y. Effects of PVP-Coated Silver Nanoparticles on Enzyme Activity, Bacterial and Archaeal Community Structure and Function in a Yellow-Brown Loam Soil. Environ. Sci. Pollut. Res. 2020, 27, 8058–8070. [Google Scholar] [CrossRef]
- Taş, A.; Keklikcioğlu Çakmak, N.; Agbektaş, T.; Zontul, C.; Özmen, E.; Siliğ, Y. Determination of the impacts of titanium dioxide nanoparticles on a number of xenobiotic-metabolizing enzymes in rat liver. Sci. J. Mehmet Akif Ersoy Univ. 2020, 3, 77–83. [Google Scholar]
- Kolesnikov, S.I.; Timoshenko, A.; Minnikova, T.; Tsepina, N.I.; Kazeev, K.; Yuliya Akimenko, V.; Zhadobin, A.; Shuvaeva, V.; Rajput, V.D.; Mandzhieva, S.; et al. Impact of Metal-Based Nanoparticles on Cambisol Microbial Functionality, Enzyme Activity, and Plant Growth. Plants 2021, 10, 2080. [Google Scholar] [CrossRef]
- Grün, A.-L.; Emmerling, C. Long-Term Effects of Environmentally Relevant Concentrations of Silver Nanoparticles on Major Soil Bacterial Phyla of a Loamy Soil. Environ. Sci. Eur. 2018, 30, 31. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.E.; Gass, M.; Muller, K.; Skepper, J.N.; Midgley, P.A.; Welland, M.E. Direct Imaging of Single-Walled Carbon Nanotubes in Cells. Nat. Nanotechnol. 2007, 2, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, S.; Bera, T.; Roy, A.; Dash, D. Characterization of enhanced antibacterial effects of novel silvernanoparticles. Nanotechnology 2007, 18, 225103. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Shen, C.; Ji, Q.; An, H.; Wang, J.; Liu, Q.; Zhang, Z. Food Storage Material Silver Nanoparticles Interfere with DNA Replication Fidelity and Bind with DNA. Nanotechnology 2009, 20, 085102. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.; de Souza, A.R.; Orjuela, G.L.; Da Silva, R.; de Oliveira, T.B.; Rodrigues, A. Applications and Benefits of Thermophilic Microorganisms and Their Enzymes for Industrial Biotechnology. In Gene Expression Systems in Fungi: Advancements and Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 459–492. [Google Scholar] [CrossRef]
- Mehta, R.; Singhal, P.; Singh, H.; Damle, D.; Sharma, A.K. Insight into Thermophiles and Their Wide-Spectrum Applications. 3 Biotech 2016, 6, 81. [Google Scholar] [CrossRef]
- Dumorne, K.; Cordova, D.C.; Astorga-Elo, M.; Renganathan, P. Extremozymes: A Potential Source for Industrial Applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Hasnaa, R.; Temsaah, A.F.; Azmy, M.R.; Amr, E.A.; Walaa, G.H. Isolation and Characterization of Thermophilic Enzymes Producing Microorganisms for Potential Therapeutic and Industrial Use. J. Pure Appl. Microbiol. 2018, 12, 1687–1702. [Google Scholar] [CrossRef]
- Ayisa, T.T.; Oyedokun, N.O.; Ideh, R.; Itoroabasi, J.O.; Ideh, R.R.; Egbi, T.K.; Ahmadu, J.H.; Orji, S.L. Thermophiles and Their Industrial Application: A Review. Afr. J. Sci. Technol. Res. 2022, 4, 63–70. [Google Scholar]
- Özdemir, S.; Kilinc, E.; Poli, A.; Nicolaus, B.; Güven, K. Biosorption of Cd, Cu, Ni, Mn and Zn from Aqueous Solutions by Thermophilic Bacteria, Geobacillus toebii subsp. decanicus and Geobacillus thermoleovorans subsp. stromboliensis: Equilibrium, Kinetic and Thermodynamic Studies. J. Chem. Eng. 2009, 152, 195–206. [Google Scholar] [CrossRef]
- Özdemir, S.; Kilinc, E.; Poli, A.; Nicolaus, B.; Güven, K. Cd, Cu, Ni, Mn and Zn resistance and bioaccumulation by thermophilic bacteria, Geobacillus toebii subsp. decanicus and Geobacillus thermoleovorans subsp. stromboliensis. World J. Microbiol. Biotechnol. 2012, 28, 155–163. [Google Scholar] [CrossRef]
- Alkan, H.; Gul-Guven, R.; Guven, K.; Erdogan, S.; Dogru, M. Biosorption of Cd+2, Cu+2, and Ni+2 Ions by a Thermophilic Haloalkalitolerant Bacterial Strain (KG9) Immobilized on Amberlite XAD-4. Pol. J. Environ. Stud. 2015, 24, 1903–1910. [Google Scholar] [CrossRef] [PubMed]
- Koul, B.; Chaudhary, R.; Taak, P. Extremophilic microbes and their application in bioremediation of environmental contaminants. In Microbe Mediated Remediation of Environmental Contaminants; Woodhead Publishing: Sawston, UK, 2021; pp. 115–128. [Google Scholar] [CrossRef]
- Gul Guven, R.; Guven, K.; Poli, A.; Nicolaus, B. Purification and Some Properties of a β- galactosidase from The Thermoacidophilic Alicyclobacillus acidocaldarius subsp. rittmannii isolated from Antarctica. Enzym. Microb. Technol. 2007, 40, 1570–1577. [Google Scholar] [CrossRef]
- Gul Guven, R.; Kaplan, A.; Guven, K.; Matpan, F.; Doğru, M. Effects of Various Inhibitors on β-galactosidase Purified from the Thermoacidophilic Alicyclobacillus acidocaldarius subsp. Rittmanni Isolated from Antarctica. Biotechnol. Bioprocess Eng. 2011, 16, 114–119. [Google Scholar] [CrossRef]
- Matpan Bekler, F.; Stougaard, P.; Guven, K.; Gul Guven, R.; Acer, O. Cloning, purification and characterization of a thermostable β-galactosidase from Bacillus licheniformis strain KG9. Cell. Mol. Biol. 2015, 61, 71–78. [Google Scholar] [PubMed]
- Matpan Bekler, F.; Yalaz, S.; Acer, O.; Guven, K. Purification of thermostable β-galactosidase from Anoxybacillus sp. KP1 and estimation of combined effect of some chemicals on enzyme activity using semiparametric errors in variables model. Fresenius Environ. Bull. 2017, 26, 2251–2259. [Google Scholar]
- Tunç, Ş.; Maptan Bekler, F.; Güven, K. Characterization of intracellular β-galactosidase from Bacillus subtilis 4NK and Bacillus paralicheniformis 5NK isolated from a hot water spring and effects of various inhibitors on enzyme activity. Biotech Stud. 2021, 30, 71–78. [Google Scholar] [CrossRef]
- Dong, F.; Valsami-Jones, E.; Kreft, J.U. New, rapid method to measure dissolved silver concentration in silver nanoparticle suspensions by aggregation combined with centrifugation. J. Nanopart. Res. 2016, 18, 259. [Google Scholar] [CrossRef] [PubMed]
- Matpan Bekler, F.; Yalaz, S.; Guven, K.; Gul Guven, R. Isolation and characterization of thermophilic bacteria from hot spring in Çermik, Diyarbakır. In Proceedings of the International Eurasian Conference on Biological and Chemical Sciences (EurasianBioChem 2020), Ankara, Turkey, 19–20 April 2020; pp. 132–139. [Google Scholar]
- Matpan Bekler, F.; Yalaz, S.; Güven, K. Molecular Characterisation and Numerical Analysis of Novel Moderately Thermophile Anoxybacillus sp. FMB1. Rom. Biotechnol. Lett. 2018, 23, 13964–13975. [Google Scholar]
- Matpan Bekler, F.; Yalaz, S.; Gul Guven, R.; Guven, K. (2019). Optimization of the Thermostable Alkaline and Ca-dependent a-Amylase Production from Bacillus paralicheniformis by Statistical Modeling. J. Serbian Chem. Soc. 2019, 84, 1093–1104. [Google Scholar] [CrossRef]
- Guven, K.; Togrul, S.; Uyar, F.; Ozant, S.; De Pomerai, D.I. A Comparative Study of Bioassays Based on Enzyme Biosynthesis in Escherichia coli and Bacillus subtilis Exposed to Heavy Metals and Organic Pesticides. Enzym. Microb. Technol. 2003, 32, 658–664. [Google Scholar] [CrossRef]
- Guven, K.; Yolcu, M.; Gul-Guven, R.; Erdogan, S.; Pomerai, D.D. The Effects of Organic Pesticides on Inner Membrane Permeability in Escherichia coli ML35. Cell Biol. Toxicol. 2005, 21, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, Applications and Toxicities. Arab. J. Chem. 2017, 12, 908–931. [Google Scholar] [CrossRef]
- Ameen, F.; Alsamhary, K.; Alabdullatif, J.A.; ALNadhari, S. A review on metal-based nanoparticles and their toxicity to beneficial soil bacteria and fungi. Ecotoxicol. Environ. Saf. 2021, 213, 112027. [Google Scholar] [CrossRef]
- Seol, J.W.; Hur, T.Y.; Jung, Y.H.; Kang, S.J.; Park, S.Y. Evaluation of bactericidal capacity of silver ion against bovine mastitis pathogens. J. Vet. Clin. 2010, 27, 252–256. [Google Scholar]
- Sintubin, L.; De Gusseme, B.; Van der Meeren, P. The antibacterial activity of biogenic silver and its mode of action. Appl. Microbiol. Biotechnol. 2011, 91, 153–162. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, L.; Si, Y.; Shu, K. Size-Dependent Cytotoxicity of Silver Nanoparticles to Azotobacter Vinelandii: Growth Inhibition, Cell Injury, Oxidative Stress and Internalization. PLoS ONE 2018, 13, e0209020. [Google Scholar] [CrossRef]
- Ahmed, B.; Ameen, F.; Rizvi, A.; Ali, K.; Sonbol, H.; Zaidi, A.; Khan, M.S.; Musarrat, J. Destruction of Cell Topography, Morphology, Membrane, Inhibition of Respiration, Biofilm Formation, and Bioactive Molecule Production by Nanoparticles of Ag, ZnO, CuO, TiO2, and al2O3 toward Beneficial Soil Bacteria. ACS Omega 2020, 5, 7861–7876. [Google Scholar] [CrossRef]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic Basis of Antimicrobial Actions of Silver Nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef]
- Dovbeshko, G. FTIR Spectroscopy Studies of Nucleic Acid Damage. Talanta 2000, 53, 233–246. [Google Scholar] [CrossRef]
- Hou, Y.C.; Zhao, P.; Zhang, F.; Yang, S.; Rady, A.; Wijewardane, N.K.; Huang, J.; Li, M. Fourier-Transform Infrared Spectroscopy and Machine Learning to Predict Amino Acid Content of Nine Commercial Insects. Food Sci. Technol. 2022, 42, e100821. [Google Scholar] [CrossRef]
- Mordechai, S.; Sahu, R.K.; Hammody, Z.; Mark, S.; Kantarovich, K.; Guterman, H.; Podshyvalov, A.; Goldstein, J.; Argov, S. Possible Common Biomarkers from FTIR Microspectroscopy of Cervical Cancer and Melanoma. J. Microsc. 2004, 215, 86–91. [Google Scholar] [CrossRef]
- Nandiyanto, A.B.D.; Andika, R.; Aziz, M.; Riza, L.S. Working Volume and Milling Time on the Product Size/Morphology, Product Yield, and Electricity Consumption in the Ball-Milling Process of Organic Material. Indones. J. Sci. Technol. 2018, 3, 82. [Google Scholar] [CrossRef]
- Brisebois, P.P.; Arnold, A.A.; Chabre, Y.M.; Roy, R.; Marcotte, I. Comparative Study of the Interaction of Fullerenol Nanoparticles with Eukaryotic and Bacterial Model Membranes Using Solid-State NMR and FTIR Spectroscopy. Eur. Biophys. J. 2012, 41, 535–544. [Google Scholar] [CrossRef]
- Jackson, M.; Sowa, M.G.; Mantsch, H.H. Infrared spectroscopy: A new frontier in medicine. Biophys. Chem. 1997, 68, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Szalontai, B.; Nishiyama, Y.; Gombos, Z.; Murata, N. Membrane dynamics as seen by Fourier transform infrared spectroscopy in a cyanobacterium, Synechocystis PCC 6803: The effects of lipid unsaturation and the protein-to-lipid ratio. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1509, 409–419. [Google Scholar] [CrossRef]
- Banyay, M.; Sarkar, M.; Gräslund, A. A library of IR bands of nucleic acids in solution. Biophys. Chem. 2003, 104, 477–488. [Google Scholar] [CrossRef]
- Saraeva, I.; Tolordava, E.; Yushina, Y.; Sozaev, I.; Sokolova, V.; Khmelnitskiy, R.; Sheligyna, S.; Pallaeva, T.; Pokryshkin, N.; Khmelenin, D.; et al. Direct Bactericidal Comparison of Metal Nanoparticles and Their Salts against S. aureus Culture by TEM and FT-IR Spectroscopy. Nanomaterials 2022, 12, 3857. [Google Scholar] [CrossRef]
- Azim, A.A.; Aksel, H.; Zhuang, T.; Mashtare, T.; Babu, J.P.; Huang, G.T.J. Efficacy of 4 Irrigation Protocols in Killing Bacteria Colonized in Dentinal Tubules Examined by a Novel Confocal Laser Scanning Microscope Analysis. J. Endod. 2016, 42, 928–934. [Google Scholar] [CrossRef]
- Javvaji, P.K.; Dhali, A.; Francis, J.; Kolte, A.P.; Mech, A.; Roy, S.C.; Mishra, A.K.; Bhatta, R. An Efficient Nitroblue Tetrazolium Staining and Bright-Field Microscopy Based Method for Detecting and Quantifying Intracellular Reactive Oxygen Species in Oocytes, Cumulus Cells and Embryos. Front. Cell Dev. Biol. 2020, 8, 764. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.A.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J.; Ischiropoulos, H. Measuring Reactive Oxygen and Nitrogen Species with Fluorescent Probes: Challenges and Limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef]
- Patlolla, A.K.; Hackett, D.; Tchounwou, P.B. Silver Nanoparticle-Induced Oxidative Stress-Dependent Toxicity in Sprague-Dawley Rats. Mol. Cell. Biochem. 2014, 399, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Sun, J.; Chang, X.; Li, W.; Li, J.; Niu, S.; Kong, L.; Zhang, T.; Wu, T.; Tang, M.; et al. Biodistribution and Organ Oxidative Damage Following 28 Days Oral Administration of Nanosilver With/without Coating in Mice. J. Appl. Toxicol. 2020, 40, 815–831. [Google Scholar] [CrossRef]
- Kim, S.; Lee, H.-S.; Ryu, D.-S.; Young Choi, S.; Lee, D.-S. Antibacterial Activity of Silver-Nanoparticles against Staphylococcus aureus and Escherichia coli. Korean J. Microbiol. Biotechnol. 2011, 39, 77–85. [Google Scholar]
- Wu, D.; Fan, W.; Kishen, A.; Gutmann, J.L.; Fan, B. Evaluation of the Antibacterial Efficacy of Silver Nanoparticles against Enterococcus faecalis Biofilm. J. Endod. 2014, 40, 285–290. [Google Scholar] [CrossRef]
- Matpan Bekler, F.; Yalaz, S.; Gul Guven, R.; Acer, O.; Guven, K. Characterization of Thermostable β-Galactosidase from Anoxybacillus ayderensis and Optimal Design for Enzyme Inhibition using Semiparametric EIV Models. Online J. Sci. Technol. 2018, 8, 32–38. [Google Scholar]
- Yalaz, S.; Matpan Bekler, F.; Acer, O. Combined effects of some chemicals on b-galactosidase activity using a new semiparametric errors in variables model. Banat’s J. Biotechnol. 2019, 10, 42–50. [Google Scholar] [CrossRef]
- Acer, Ö.; Matpan Bekler, F. Characterization of a Thermally Stable β-galactosidase Produced by Thermophilic Anoxybacillus sp. AH1. Turk. J. Nat. Sci. 2021, 10, 130–136. [Google Scholar] [CrossRef]
- Jung, W.K.; Koo, H.C.; Kim, K.W.; Shin, S.; Kim, S.H.; Park, Y.H. Antibacterial Activity and Mechanism of Action of the Silver Ion in Staphylococcus Aureus and Escherichia Coli. Appl. Environ. Microbiol. 2008, 74, 2171–2178. [Google Scholar] [CrossRef]
- Rai, M.K.; Deshmukh, S.D.; Ingle, A.P.; Gade, A.K. Silver Nanoparticles: The Powerful Nanoweapon against Multidrug-Resistant Bacteria. J. Appl. Microbiol. 2012, 112, 841–852. [Google Scholar] [CrossRef]
- Holt, K.B.; Bard, A.J. Interaction of silver(I) ions with the respiratory chain of Escherichia coli: An electrochemical and scanning electro-chemical microscopy study of the antimicrobial mechanism of micromolar Ag+. Biochemistry 2005, 44, 13214–13223. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Han, C.; Wang, X.; Zheng, Y.; Li, Q.; Hu, X.; Sun, H. The progress of silver nanoparticles in the antibacterial mechanism, clinical application and cytotoxicity. Mol. Biol. Rep. 2012, 39, 9193–9201. [Google Scholar] [CrossRef] [PubMed]
- Pareek, V.; Gupta, R.; Panwar, J. Do Physico-Chemical Properties of Silver Nanoparticles Decide Their Interaction with Biological Media and Bactericidal Action? A Review. Mater. Sci. Eng. C 2018, 90, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Zheng, J. Antibacterial Activity of Silver Nanoparticles: Structural Effects. Adv. Healthc. Mater. 2018, 7, 1701503. [Google Scholar] [CrossRef]
- Sondi, I.; Salopek-Sondi, B. Silver Nanoparticles as Antimicrobial Agent: A Case Study on E. coli as a Model for Gram-Negative Bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef]
- Ahmad, A.; Wei, Y.; Syed, F.; Tahir, K.; Rehman, A.U.; Khan, A.; Ullah, S.; Yuan, Q. The Effects of Bacteria-Nanoparticles Interface on the Antibacterial Activity of Green Synthesized Silver Nanoparticles. Microb. Pathog. 2017, 102, 133–142. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AgNP Characteristics | |
|---|---|
| Purity\ | 99.5% (metal basis) |
| Average particle size | 35 nm |
| Specific surface area | 20–30 m2/g |
| Particle morphology | Spherical |
| Appearance | Gray powder |
| Bacterial Strains | Shape | Spore Forming | Pigmentation | Optimum Temperature (°C) | Optimum pH | Reference |
|---|---|---|---|---|---|---|
| G. vulcani 2Cx | rod | + | light yellow | 55 | 7.0 | [53] |
| B. licheniformis 3CA | rod | + | creamy white | 55 | 8.0 | [53] |
| P. macerans 3CA1 | rod | + | creamy white | 50 | 7.0 | [53] |
| A. ayderensis FMB1 | rod | + | yellow/orange | 50 | 7.0 | [54] |
| B. paralicheniformis FMB2-1 | rod | + | creamy white | 50 | 7.0 | [55] |
| Bacterial Strains | AgNPs (μg mL−1) | |
|---|---|---|
| MIC | MBC | |
| G. vulcani 2Cx | 500 | 1000 |
| B. licheniformis 3CA | 1000 | 1500 |
| P. macerans 3CA1 | 500 | 1000 |
| A. ayderensis FMB1 | 1000 | 1500 |
| B. paralicheniformis FMB2-1 | 500 | 1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahan, I.; Matpan Bekler, F.; Tunç, A.; Güven, K. The Effects of Silver Nanoparticles (AgNPs) on Thermophilic Bacteria: Antibacterial, Morphological, Physiological and Biochemical Investigations. Microorganisms 2024, 12, 402. https://doi.org/10.3390/microorganisms12020402
Jahan I, Matpan Bekler F, Tunç A, Güven K. The Effects of Silver Nanoparticles (AgNPs) on Thermophilic Bacteria: Antibacterial, Morphological, Physiological and Biochemical Investigations. Microorganisms. 2024; 12(2):402. https://doi.org/10.3390/microorganisms12020402
Chicago/Turabian StyleJahan, Israt, Fatma Matpan Bekler, Ahmed Tunç, and Kemal Güven. 2024. "The Effects of Silver Nanoparticles (AgNPs) on Thermophilic Bacteria: Antibacterial, Morphological, Physiological and Biochemical Investigations" Microorganisms 12, no. 2: 402. https://doi.org/10.3390/microorganisms12020402