
Genomic Insights into the Symbiotic and Plant Growth-Promoting Traits of “Candidatus Phyllobacterium onerii” sp. nov. Isolated from Endemic Astragalus flavescens

Abstract
:1. Introduction
2. Materials and Methods
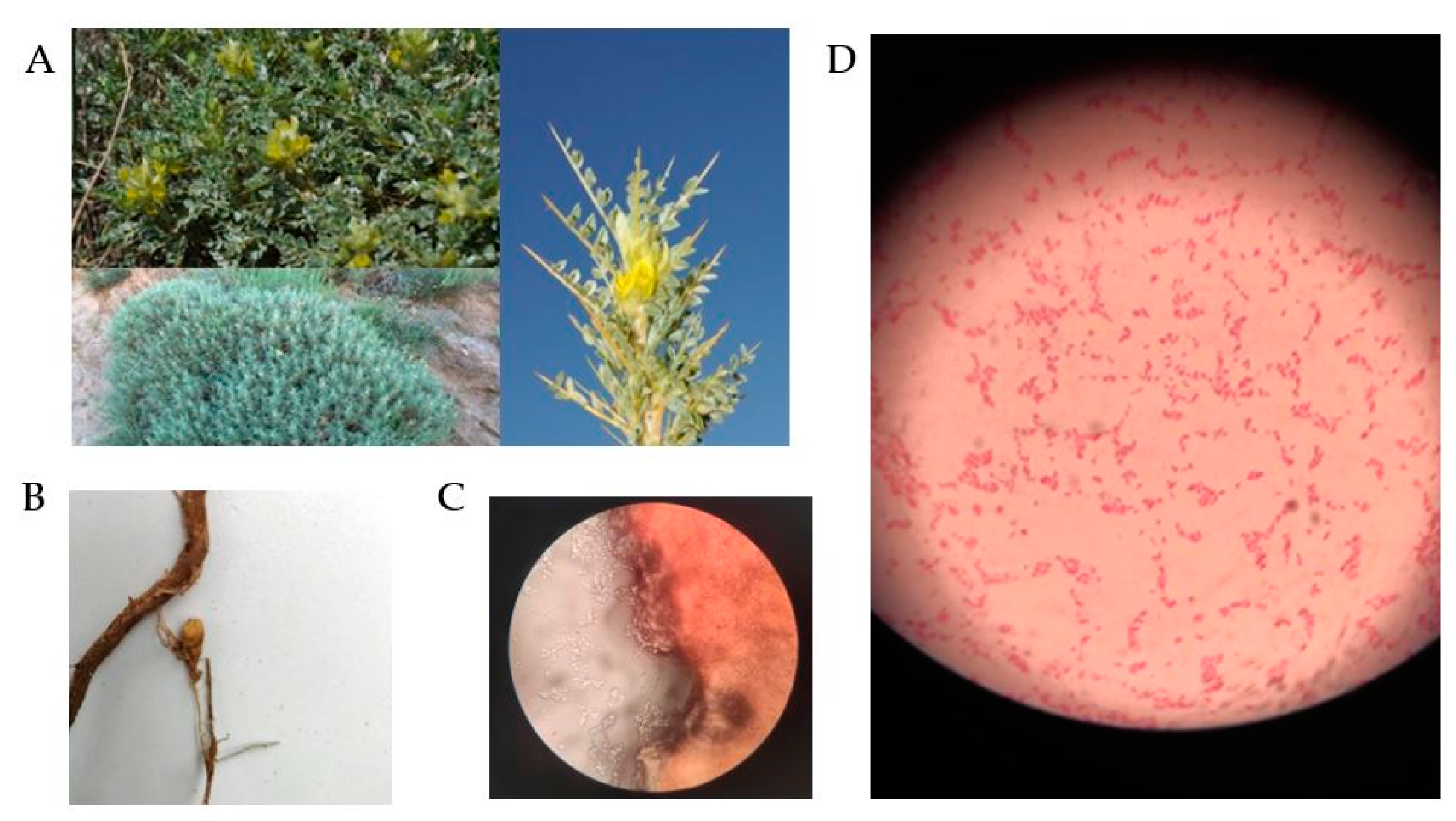
2.1. Sample Collection and Isolation
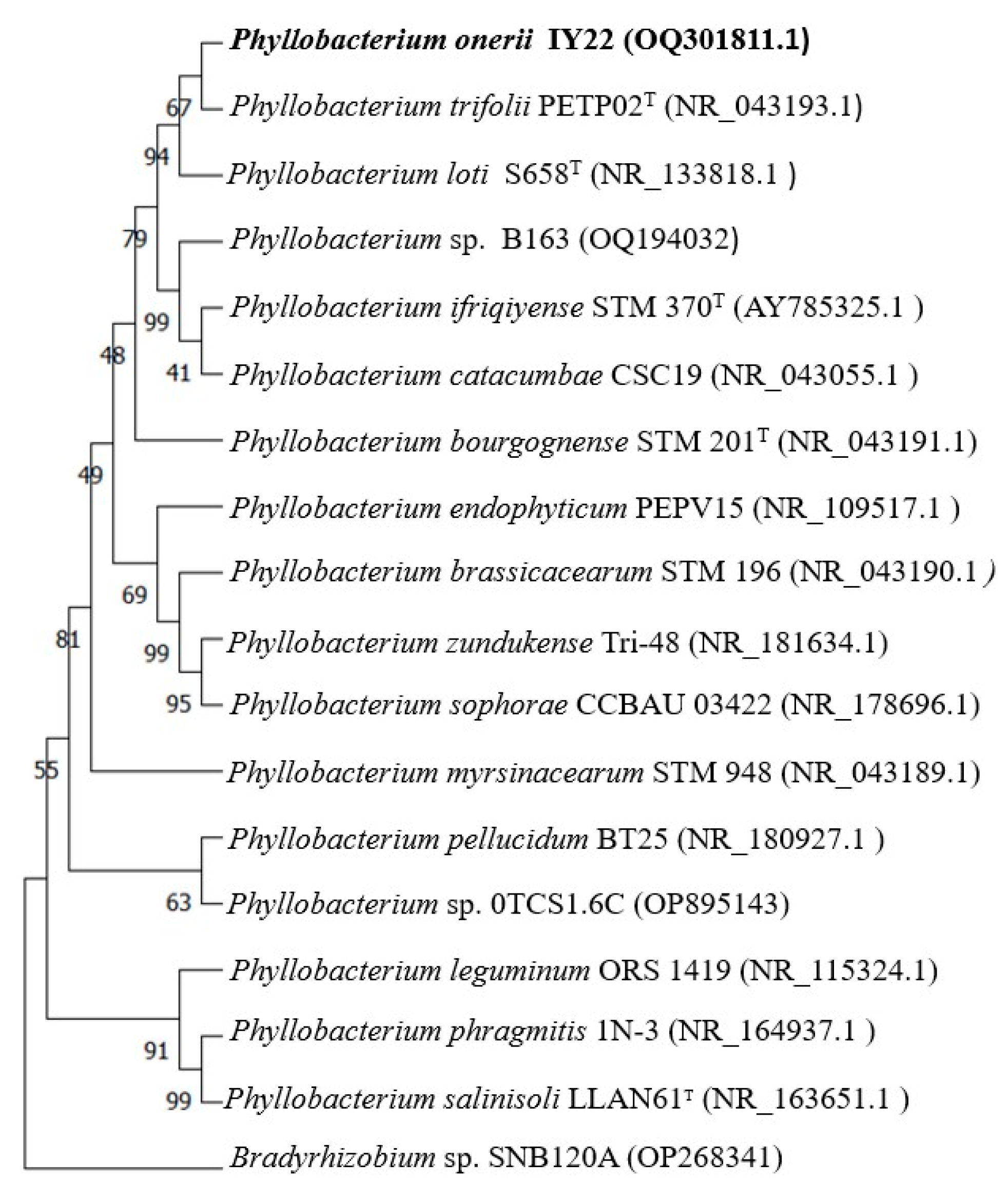
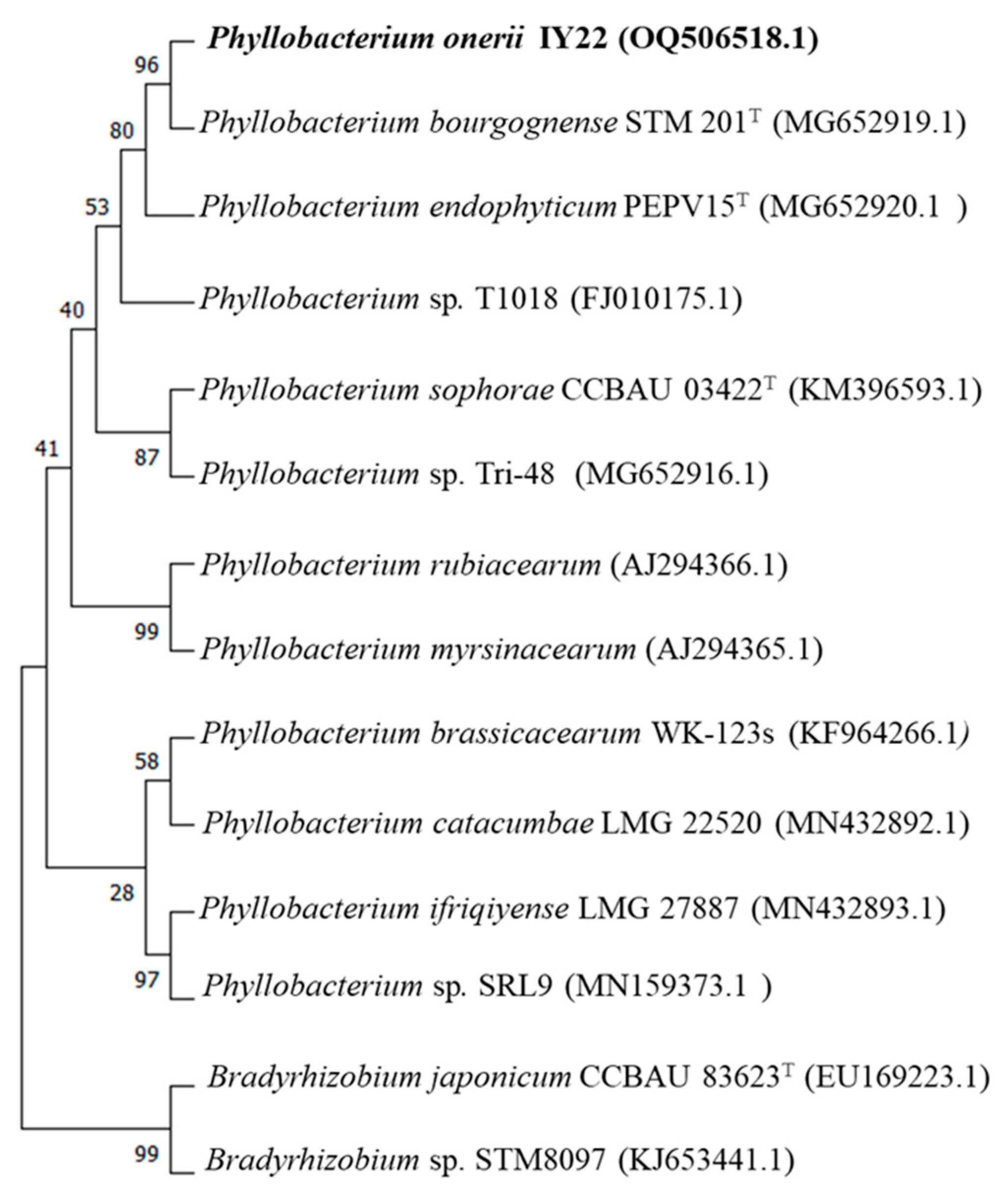
2.2. Phylogeny of 16S rDNA and recA Genes
2.3. Phenotypic Characterization
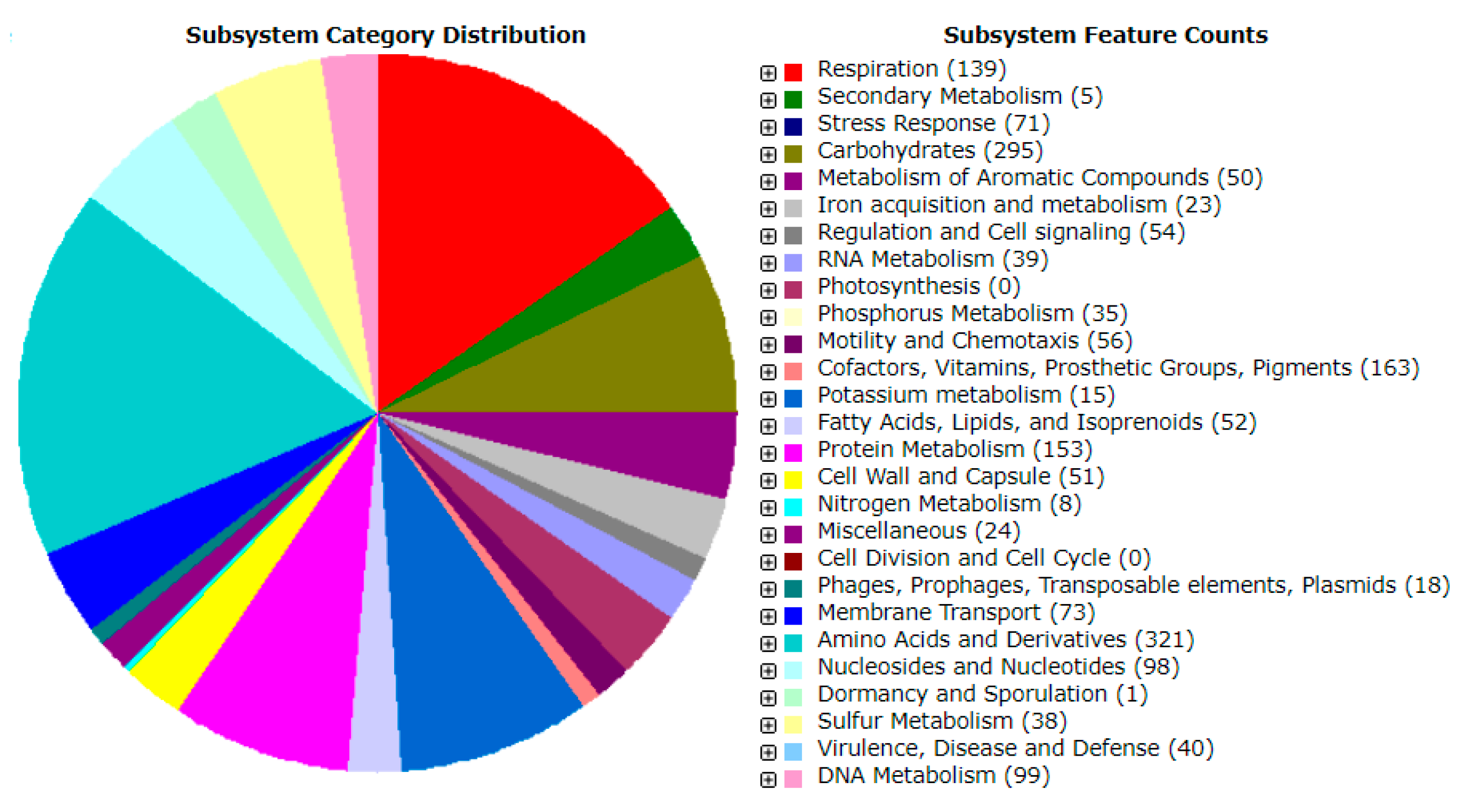
2.4. Genome Analysis
3. Results and Discussion
4. Conclusions
Description of “Candidatus Phyllobacterium onerii” sp. nov.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mantelin, S.; Fischer-Le Saux, M.; Zakhia, F.; Béna, G.; Bonneau, S.; Jeder, H.; de Lajudie, P.; Cleyet-Marel, J.C. Emended description of the genus Phyllobacterium and description of four novel species associated with plant roots: Phyllobacterium bourgognense sp. nov., Phyllobacterium ifriqiyense sp. nov., Phyllobacterium leguminum sp. nov. and Phyllobacterium brassicacearum sp. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Alavi, M.; Miller, T.; Erlandson, K.; Schneider, R.; Belas, R. Bacterial community associated with Pfiesteria-like dinoflagellate cultures. Environ. Microbiol. 2001, 3, 380–396. [Google Scholar] [CrossRef]
- Gonzalez-Bashan, L.E.; Lebsky, V.K.; Hernandez, J.P.; Bustillos, J.J.; Bashan, Y. Changes in the metabolism of the microalga Chlorella vulgaris when coimmobilized in alginate with the nitrogen-fixing Phyllobacterium myrsinacearum. Can. J. Microbiol. 2000, 46, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Laiz, L.; Gonzalez, J.M.; Hernandez-Marine, M.; Valens, M.; Sáiz-Jiménez, C. Phyllobacterium catacumbae sp. nov., a member of the order ‘Rhizobiales’ isolated from Roman catacombs. Int. J. Syst. Evol. Microbiol. 2005, 55, 1487–1490. [Google Scholar] [CrossRef]
- Lambert, B.; Joos, H.; Dierickx, S.; Vantomme, R.; Swings, J.; Kersters, K.; Van Montagu, M. Identification and plant interaction of a Phyllobacterium sp., a predominant rhizobacterium of young sugar beet plants. Appl. Environ. Microbiol. 1990, 56, 1093–1102. [Google Scholar] [CrossRef]
- Mergaert, J.; Cnockaert, M.C.; Swings, J. Phyllobacterium myrsinacearum (subjective synonym Phyllobacterium rubiacearum) emend. Int. J. Syst. Evol. Microbiol. 2002, 52, 1821–1823. [Google Scholar] [CrossRef]
- Knösel, D. Prüfung von bakterien auf Fähigkeit zur Sternbildung. Zentralbl Bakteriol. Parasitenkd. Infekt. Hyg. 1962, 116, 79–100. [Google Scholar]
- Knosel, D.H. Genus IV. Phylloacterium (ex Knosel 1962) nom. rev. (Phyllobacterium Knosel 1962, 96). Bergey’s Man. Syst. Bacteriol. 1984, 1, 254–256. [Google Scholar]
- Valverde, A.; Velazquez, E.; Fernandez-Santos, F.; Vizcaíno, N.; Rivas, R.; Mateos, P.F.; Martínez-Molina, E.; Igual, J.M.; Willems, A. Phyllobacterium trifolii sp. nov., nodulating Trifolium and Lupinus in Spanish soils. Int. J. Syst. Evol. Microbiol. 2005, 55, 1985–1989. [Google Scholar] [CrossRef]
- Flores-Felix, J.D.; Carro, L.; Velazquez, E.; Valverde, A.; Cerda-Castillo, E.; García-Fraile, P.; Rivas, R. Phyllobacterium endophyticum sp. nov., isolated from nodules of Phaseolus vulgaris. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 3, 821–826. [Google Scholar] [CrossRef]
- Sanchez, M.; Ramirez-Bahena, M.H.; Peix, A.; Lorite, M.J.; Sanjuan, J.; Velazquez, E.; Monza, J. Phyllobacterium loti sp. nov. isolated from nodules of Lotus corniculatus. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 3, 781. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.S.; Yan, H.; Ji, Z.J.; Liu, Y.H.; Sui, X.H.; Zhang, X.X.; Wang, E.T.; Chen, W.X. Phyllobacterium sophorae sp. nov., a symbiotic bacterium isolated from root nodules of Sophora flavescens. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 2, 399–406. [Google Scholar] [CrossRef] [PubMed]
- León-Barrios, M.; Ramírez-Bahena, M.H.; Igual, J.M.; Peix, Á.; Velázquez, E. Phyllobacterium salinisoli sp. nov., isolated from a Lotus lancerottensis root nodule in saline soil from Lanzarote. Int. J. Syst. Evol. Microbiol. 2018, 68, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Safronova, V.I.; Sazanova, A.L.; Kuznetsova, I.G.; Belimov, A.A.; Andronov, E.E.; Chirak, E.R.; Popova, J.P.; Verkhozina, A.V.; Willems, A.; Tikhonovich, I.A. Phyllobacterium zundukense sp. nov., a novel species of rhizobia isolated from root nodules of the legume species Oxytropis triphylla (Pall.) Pers. Int. J. Syst. Evol. Microbiol. 2018, 68, 1644–1651. [Google Scholar] [CrossRef]
- Liang, L.X.; Sun, Q.W.; Hui, N.; Zhang, X.X.; Li, L.B.; Liu, L. Phyllobacterium phragmitis sp. nov., an endophytic bacterium isolated from Phragmites australis rhizome in Kumtag Desert. Antonie Leeuwenhoek 2019, 112, 661–668. [Google Scholar] [CrossRef]
- Park, Y.; Ten, L.N.; Maeng, S.; Chang, Y.; Jung, H.Y.; Kim, M.K. Phyllobacterium pellucidum sp. nov., isolated from soil. Arch. Microbiol. 2021, 203, 2647–2652. [Google Scholar] [CrossRef]
- Lustermans, J.J.; Bjerg, J.J.; Schramm, A.; Marshall, I.P. Phyllobacterium calauticae sp. nov. isolated from a microaerophilic veil transversed by cable bacteria in freshwater sediment. Antonie Leeuwenhoek 2021, 114, 1877–1887. [Google Scholar] [CrossRef]
- Shumilina, J.; Soboleva, A.; Abakumov, E.; Shtark, O.Y.; Zhukov, V.A.; Frolov, A. Signaling in Legume-Rhizobia Symbiosis. Int. J. Mol. Sci. 2023, 24, 17397. [Google Scholar] [CrossRef]
- Singh, J.; Verma, P.K. Role of Nod factor receptors and its allies involved in nitrogen fixation. Planta 2023, 257, 54. [Google Scholar] [CrossRef]
- Mbengue, M.D.; Hervé, C.; Debellé, F. Nod factor signaling in symbiotic nodulation. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2020; Volume 94, pp. 1–39. [Google Scholar] [CrossRef]
- Bouremani, N.; Cherif-Silini, H.; Silini, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N.; Baranov, O.; Belbahri, L. Plant growth-promoting rhizobacteria (PGPR): A rampart against the adverse effects of drought stress. Water 2023, 15, 418. [Google Scholar] [CrossRef]
- Wang, D.; Poinsot, V.; Li, W.; Lu, Y.; Liu, C.; Li, Y.; Xie, K.; Sun, L.; Shi, C.; Peng, H.; et al. Genomic insights and functional analysis reveal plant growth promotion traits of Paenibacillus mucilaginosus G78. Genes 2023, 14, 392. [Google Scholar] [CrossRef]
- Świątczak, J.; Kalwasińska, A.; Wojciechowska, A.; Brzezinska, M.S. Physiological properties and genomic insights into the plant growth-promoting rhizobacterium Brevibacillus laterosporus K75 isolated from maize rhizosphere. J. Sci. Food Agric. 2023, 103, 1432–1441. [Google Scholar] [CrossRef]
- Leontidou, K.; Genitsaris, S.; Papadopoulou, A.; Kamou, N.; Bosmali, I.; Matsi, T.; Madesis, P.; Vokou, D.; Karamanoli, K.; Mellidou, I. Plant growth promoting rhizobacteria isolated from halophytes and drought-tolerant plants: Genomic characterisation and exploration of phyto-beneficial traits. Sci. Rep. 2020, 10, 14857. [Google Scholar] [CrossRef]
- Medina-Cordoba, L.K.; Chande, A.T.; Rishishwar, L.; Mayer, L.W.; Valderrama-Aguirre, L.C.; Valderrama-Aguirre, A.; Gaby, J.C.; Kostka, J.E.; Jordan, I.K. Genomic characterization and computational phenotyping of nitrogen-fixing bacteria isolated from Colombian sugarcane fields. Sci. Rep. 2021, 11, 9187. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, W.; Jiang, Q.; Fei, Z.; Xiao, M. Genome analysis of plant growth-promoting rhizobacterium Pseudomonas chlororaphis subsp. aurantiaca JD37 and insights from comparasion of genomics with three Pseudomonas strains. Microbiol. Res. 2020, 237, 126483. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H. (Ed.) Introduction to Bioinformatics in Microbiology; Springer International Publishing: Cham, Switzerland, 2018; p. 213. [Google Scholar] [CrossRef]
- De Lajudie, P.M.; Andrews, M.; Ardley, J.; Eardly, B.; Jumas-Bilak, E.; Kuzmanović, N.; Lassalle, F.; Lindström, K.; Mhamdi, R.; Martínez-Romero, E.; et al. Minimal standards for the description of new genera and species of rhizobia and agrobacteria. Int. J. Syst. Evol. Microbiol. 2019, 69, 1852–1863. [Google Scholar] [CrossRef] [PubMed]
- Ormeno-Orrillo, E.; Servín-Garcidueñas, L.E.; Rogel, M.A.; González, V.; Peralta, H.; Mora, J.; Martínez-Romero, J.; Martínez Romero, E. Taxonomy of rhizobia and agrobacteria from the Rhizobiaceae family in light of genomics. Syst. Appl. Microbiol. 2015, 38, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.M. A Manual for the Practical Study of the Root-Nodule Bacteria; Blackwell Scientific: Oxford, UK, 1970. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Ouk Kim, Y.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Sun, H.X.; Zhang, C.; Cheng, L.; Peng, Y.; Deng, Z.; Wang, D.; Wang, Y.; Hu, M.; Liu, W.; et al. Prophage Hunter: An integrative hunting tool for active prophages. Nucleic Acids Res. 2019, 47, W74–W80. [Google Scholar] [CrossRef] [PubMed]
- Ferraz Helene, L.C.; Klepa, M.S.; Hungria, M. New insights into the taxonomy of bacteria in the genomic era and a case study with rhizobia. Int. J. Microbiol. 2022, 2022, 4623713. [Google Scholar] [CrossRef] [PubMed]
- González, V.; Santamaría, R.I.; Bustos, P.; Pérez-Carrascal, O.M.; Vinuesa, P.; Juárez, S.; Martínez-Flores, I.; Cevallos, M.; Brom, S.; Martinez-Romero, E.; et al. Phylogenomic Rhizobium species are structured by a continuum of diversity and genomic clusters. Front. Microbiol. 2019, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
- Manian, S.S.; Gumbleton, R.; O’Gara, F. The role of formate metabolism in nitrogen fixation in Rhizobium Spp. Arch. Microbiol. 1982, 133, 312–317. [Google Scholar] [CrossRef]
- Pickering, B.S.; Oresnik, I.J. Formate-dependent autotrophic growth in Sinorhizobium meliloti. J. Bacteriol. 2008, 190, 6409–6418. [Google Scholar] [CrossRef]
- Vandamme, P.; Sutcliffe, I. Out with the old and in with the new: Time to rethink twentieth century chemotaxonomic practices in bacterial taxonomy. Int. J. Syst. Evol. Microbiol. 2021, 71, 005127. [Google Scholar] [CrossRef]
- Baek, I.; Kim, M.; Lee, I.; Na, S.I.; Goodfellow, M.; Chun, J. Phylogeny trumps chemotaxonomy: A case study involving Turicella otitidis. Front. Microbiol. 2018, 9, 834. [Google Scholar] [CrossRef]
- Yamano, R.; Yu, J.; Jiang, C.; Harjuno Condro Haditomo, A.; Mino, S.; Sakai, Y.; Sawabe, T. Taxonomic revision of the genus Amphritea supported by genomic and in silico chemotaxonomic analyses, and the proposal of Aliamphritea gen. nov. PLoS ONE 2022, 17. [Google Scholar] [CrossRef]
- Yamano, R.; Yu, J.; Haditomo, A.H.C.; Jiang, C.; Mino, S.; Romalde, J.L.; Kang, K.; Sakai, Y.; Sawabe, T. Genome taxonomy of the genus Thalassotalea and proposal of Thalassotalea hakodatensis sp. nov. isolated from sea cucumber larvae. PLoS ONE 2023, 18, e0286693. [Google Scholar] [CrossRef]
- Chardot, T. A comparison of natural and induced diversity in plant oils. OCL 2022, 29, 34. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Kamisaka, Y.; Maehara, T.; Takaku, H.; Yaoi, K. Identification and characterization of two fatty acid elongases in Lipomyces starkeyi. Appl. Microbiol. Biotechnol. 2020, 104, 2537–2544. [Google Scholar] [CrossRef]
- Czolkoss, S.; Fritz, C.; Hölzl, G.; Aktas, M. Two distinct cardiolipin synthases operate in Agrobacterium tumefaciens. PLoS ONE 2016, 11, e0160373. [Google Scholar] [CrossRef]
- Zamlynska, K.; Komaniecka, I.; Zebracki, K.; Mazur, A.; Sroka-Bartnicka, A.; Choma, A. Studies on lipid A isolated from Phyllobacterium trifolii PETP02T lipopolysaccharide. Antonie Van Leeuwenhoek 2017, 110, 1413–1433. [Google Scholar] [CrossRef]
- Bresson, J.; Vasseur, F.; Dauzat, M.; Labadie, M.; Varoquaux, F.; Touraine, B.; Vile, D. Interact to survive: Phyllobacterium brassicacearum improves Arabidopsis tolerance to severe water Deficit and growth recovery. PLoS ONE 2014, 9, e107607. [Google Scholar] [CrossRef]
- Kechid, M.; Desbrosses, G.; Gamet, L.; Castaings, L.; Varoquaux, F.; Djekoun, A.; Touraine, B. Arabidopsis Growth-Promotion and Root Architecture Responses to the Beneficial Rhizobacterium Phyllobacterium brassicacearum Strain STM196 Are Independent of the Nitrate Assimilatory Pathway. Plants 2022, 11, 128. [Google Scholar] [CrossRef]
- Wulandari, D.; Songwattana, P.; Gressent, F.; Piromyou, P.; Teamtisong, K.; Boonkerd, N.; Giraud, E.; Tittabutr, P.; Teaumroong, N. Nod-Factor structure and functional redundancy of nod genes contribute the broad host range Bradyrhizobium sp. DOA9. Rhizosphere 2022, 22, 100503. [Google Scholar] [CrossRef]
- Peck, M.C.; Fisher, R.F.; Long, S.R. Diverse flavonoids stimulate NodD1 binding to nod gene promoters in Sinorhizobium meliloti. J. Bacteriol. 2006, 188, 5417–5427. [Google Scholar] [CrossRef]
- Geelen, D.; Leyman, B.; Mergaert, P.; Klarskov, K.; Van Montagu, M.; Geremia, R.; Holsters, M. NodS is an S-adenosyl-l-methionine-dependent methyltransferase that methylates chitooligosaccharides deacetylated at the non-reducing end. Mol. Microbiol. 1995, 17, 387–397. [Google Scholar] [CrossRef]
- Stacey, G. Bradyrhizobium japonicum nodulation genetics. FEMS Microbiol. Lett. 1995, 127, 1–9. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, N.; Li, S.; Brunel, B.; Wang, J.; Feng, Y.; Yang, T.; Zong, X. Genotypic composition and performance of pea-nodulating rhizobia from soils outside the native plant-host range. Front. Microbiol. 2023, 14, 1201140. [Google Scholar] [CrossRef]
- Kalita, M.; Małek, W.; Coutinho, T.A. Putative novel Bradyrhizobium and Phyllobacterium species isolated from root nodules of Chamaecytisus ruthenicus. Syst. Appl. Microbiol. 2020, 43, 126056. [Google Scholar] [CrossRef]
- Jimenez-Vicente, E.; Yang, Z.Y.; Del Campo, J.S.M.; Cash, V.L.; Seefeldt, L.C.; Dean, D.R. The NifZ accessory protein has an equivalent function in maturation of both nitrogenase MoFe protein P-clusters. J. Biol. Chem. 2019, 294, 6204–6213. [Google Scholar] [CrossRef]
- Smith, A.D.; Jameson, G.N.; Dos Santos, P.C.; Agar, J.N.; Naik, S.; Krebs, C.; Frazzon, J.; Dean, D.R.; Huynh, B.H.; Johnson, M.K. NifS-mediated assembly of [4Fe− 4S] clusters in the N-and C-terminal domains of the NifU scaffold protein. Biochemistry 2005, 44, 12955–12969. [Google Scholar] [CrossRef]
- Ribbe, M.W.; Hu, Y.; Hodgson, K.O.; Hedman, B. Biosynthesis of nitrogenase metalloclusters. Chem. Rev. 2014, 114, 4063–4080. [Google Scholar] [CrossRef]
- Maio, N.; Rouault, T.A. Iron–sulfur cluster biogenesis in mammalian cells: New insights into the molecular mechanisms of cluster delivery. Biochim. Biophys. Acta 2015, 1853, 1493–1512. [Google Scholar] [CrossRef]
- Li, Q.; Li, Y.; Li, X.; Chen, S. Identification of genes involved in Fe–S cluster biosynthesis of nitrogenase in Paenibacillus polymyxa WLY78. Int. J. Mol. Sci. 2021, 22, 3771. [Google Scholar] [CrossRef]
- Yu, Z.; Yang, G.; Liu, X.; Wang, Y.; Zhuang, L.; Zhou, S. Complete genome sequence of the nitrogen-fixing bacterium Azospirillum humicireducens type strain SgZ-5T. Stand. Genom. Sci. 2018, 13, 28. [Google Scholar] [CrossRef]
- Koirala, A.; Brözel, V.S. Phylogeny of nitrogenase structural and assembly components reveals new insights into the origin and distribution of nitrogen fixation across bacteria and archaea. Microorganisms 2021, 9, 1662. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Source | Reference |
|---|---|---|
| P. myrsinacearum | Ardisia crispa, Germany | [7] |
| P. rubiacearum (junior subjective synonym, P. myrsinacearum) | Pavetta leaf nodules, Belgium | [8] |
| P. trifolii | Trifolium pratense, Spain | [9] |
| P. catacumbae | Roman catacombs of Saint Callixtus, Italy | [4] |
| P. bourgognense | Brassica napus, France | [1] |
| P. brassicacearum | Brassica napus, France | [1] |
| P. ifriqiyense | Astragalus algerianus and Lathyrus numidicus, Tunisia | [1] |
| P. leguminum | Argyrolobium uniflorum and Astragalus algerianus, Tunisia | [1] |
| P. endophyticum | Phaseolus vulgaris, Spain | [10] |
| P. loti | Lotus corniculatus, Uruguay | [11] |
| P. sophorae | Sophora flavescens, China | [12] |
| P. salinisoli | Lotus lancerottensis, Lanzarote | [13] |
| P. zundukense | Oxytropis triphylla (Pall.) Pers, Russia | [14] |
| P. phragmitis | Phragmites australis, China | [15] |
| P. pellucidum | soil, Korea | [16] |
| P. calauticae | sediment, Denmark | [17] |
| Characteristic | P. onerii IY22 (This Study) | P. trifolii PETP02T [9] | P. bourgognense STM201T [1] |
|---|---|---|---|
| Sucrose | + | − | nd |
| Trehalose | + | − | w |
| Rhamnose | + | w | + |
| Raffinose | + | − | + |
| Citrate assimilation | + | − | + |
| Indole production | + | − | − |
| Attribute | P. onerii (GCA_028331445.1) | P. trifolii (GCA_014192095.1) | P. bourgognense (GCA_003337575.1) |
|---|---|---|---|
| Contigs | 65 | 69 | 61 |
| GC Content | 56.37 | 56.4 | 56.5 |
| Plasmids | 0 | 0 | 0 |
| Contig L50 | 5 | 9 | 7 |
| Genome Length | 6,010,116 bp | 6,306,857 bp | 5,618,596 |
| Contig N50 | 438,728 | 236,202 | 228,269 |
| Chromosomes | 1 | 1 | 1 |
| CDS | 6470 | 6715 | 5995 |
| tRNA | 46 | 41 | 46 |
| rRNA | 3 | 3 | 3 |
| Protein Features | Number |
|---|---|
| Hypothetical proteins | 2534 |
| Proteins with functional assignments | 3936 |
| Proteins with EC number assignments | 1232 |
| Proteins with GO assignments | 1087 |
| Proteins with pathway assignments | 952 |
| Proteins with PATRIC genus-specific family (PLfam) assignments | 4252 |
| Proteins with PATRIC cross-genus family (PGfam) assignments | 4701 |
| Genes | Function of Gene Product or Identity | Length (aa) | Acc. Number |
|---|---|---|---|
| nodA | N-acyltransferase | 351 | WP_271896533 |
| nodC | N-acetylglucosaminyltransferase | 423 | WP_271898196.1 |
| nodD | LysR family transcriptional regulator | 298 | WP_271898124.1 |
| nodS | SAM-dependent methyltransferase | 281 | WP_271898982.1 |
| nodN | Nodulation protein N (MaoC family dehydratase) | 159 | WP_271898282.1 |
| nodZ | GDP-L-fucose synthetase | 333 | WP_271892710.1 |
| nifU | Fe-S cluster biogenesis protein | 192 | WP_114431505.1 NZ_JAQMHX010000004.1 217482-218060 |
| nifH/frxC | 4Fe-4S binding protein | 666 | WP_271899276.1 |
| nifS | Cysteine desulfurase | 391 | WP_271893808.1 |
| sufA | Fe-S cluster assembly protein | 124 | WP_271893816.1 |
| sufB * | Fe-S cluster assembly protein | 509 | WP_112530944.1 NZ_JAQMHX010000001.1 1104702-1106231 |
| sufC * | Fe-S cluster assembly ATPase | 251 | WP_271893810.1 NZ_JAQMHX010000001.1 1106527-1107282 |
| sufD | Fe-S cluster assembly protein | 423 | WP_271893811.1 |
| sufE * | Sulfur acceptor protein | 146 | WP_224505514.1 NZ_JAQMHX010000001.1 576195-576635 |
| sufS | Cysteine desulfurase | 413 | WP_271893813.1 |
| NtrB | Nitrogen metabolism regulation gene | 384 | WP_271896598.1 |
| NtrC * | Nitrogen metabolism regulation gene | 486 | WP_271896598.1 |
| NtrX * | Nitrogen metabolism regulation gene | 486 | WP_224506676.1 NZ_JAQMHX010000005.1 410594-412054 (+) |
| GlnK * | Nitrogen regulatory protein P-II | 112 | WP_008123000 NZ_JAQMHX010000012.1 108434-108772 (+) |
| glnD | [Protein-PII] uridylyltransferase | 937 | WP_271895489.1 |
| cysE | Serine acetyltransferase | 274 | WP_271896446.1 |
| fixA | Electron transfer flavoprotein beta subunit | 250 | WP_271897294.1 |
| fixB | Electron transfer flavoprotein alfa subunit | 309 | WP_271897292.1 |
| fixC | Flavoprotein-ubiquinone oxidoreductase | 555 | WP_271893276.1 |
| fixj | Response regulator low-oxygen conditions | 124 | WP_271898449.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eren Eroğlu, A.E.; Eroğlu, V.; Yaşa, İ. Genomic Insights into the Symbiotic and Plant Growth-Promoting Traits of “Candidatus Phyllobacterium onerii” sp. nov. Isolated from Endemic Astragalus flavescens. Microorganisms 2024, 12, 336. https://doi.org/10.3390/microorganisms12020336
Eren Eroğlu AE, Eroğlu V, Yaşa İ. Genomic Insights into the Symbiotic and Plant Growth-Promoting Traits of “Candidatus Phyllobacterium onerii” sp. nov. Isolated from Endemic Astragalus flavescens. Microorganisms. 2024; 12(2):336. https://doi.org/10.3390/microorganisms12020336
Chicago/Turabian StyleEren Eroğlu, Asiye Esra, Volkan Eroğlu, and İhsan Yaşa. 2024. "Genomic Insights into the Symbiotic and Plant Growth-Promoting Traits of “Candidatus Phyllobacterium onerii” sp. nov. Isolated from Endemic Astragalus flavescens" Microorganisms 12, no. 2: 336. https://doi.org/10.3390/microorganisms12020336