1. Introduction
Snakebites are categorized as neglected tropical diseases [
1,
2,
3]. Annually, about 2.7 million people worldwide suffer from snakebites, leading to mortality cases ranging between 81,000 and 138,000 [
4]. Snakebite-related deaths not only attribute fatalities to the venom’s toxicity but also to secondary infections caused by bacteria [
5,
6,
7,
8]. Numerous studies suggest that bacterial species isolated from snakebite wounds correspond to the oral flora of the responsible snake species [
9,
10]. However, wound contamination can also result from the victim’s skin tissue, pre-hospital environments, and inadequate wound management, introducing external bacterial strains unrelated to the snake’s oral flora [
1,
11,
12,
13,
14].
Snake oral saliva contains a diverse bacterial community, including harmless environmental strains and potential pathogenic species harmful to humans [
10,
15,
16]. Following a snakebite, the venom components may lead to necrosis of affected muscles and skin tissues, creating an environment conducive to the growth of bacteria carried by snake oral saliva [
1,
8,
9,
17,
18]. Without proper attention and treatment, even bites from non-venomous snakes can result in bacterial invasion, leading to bacterial infections and the development of various chronic diseases, potentially resulting in death in severe cases [
1,
19,
20,
21]. Snake antivenom therapy may not prevent the subsequent symptoms caused by bacterial infections. Although antibiotics are commonly used to treat bacterial infections, administering them without understanding the specific infection source may be ineffective and contribute to antibiotic resistance, considering the varying sensitivity of bacterial species to antibiotics [
1,
10,
22].
Targeted approaches offer precision and quantification for known target microbes, while non-targeted approaches provide a more comprehensive and unbiased exploration of the microbial landscape. Researchers should use a combination of both approaches to gain a deeper understanding of microbiological systems. In our investigation, we initially utilized a non-targeted approach to analyze the oral microbiota of seven species of wild venomous snakes in Taiwan, employing high-throughput amplicon sequencing of the full-length 16S rRNA gene [
16]. Subsequently, we will conduct a targeted approach focusing on specific oral microbes using a molecular genotyping method. The bacterial species associated with secondary infections potentially resulting from snakebites are diverse and include aerobic Gram-positive bacteria, Gram-negative bacteria, and anaerobic bacteria (see
Supplementary Table S1). Infections caused by Gram-positive bacteria can manifest with symptoms such as abscess formation, cellulitis, sepsis, meningitis, urinary tract infections, bacterial infections at the bite site, and ocular infections.
Enterococcus faecalis, a common pathogenic Gram-positive bacterium found in the gastrointestinal tract, is frequently implicated. Most Gram-negative bacteria are pathogenic, and
Morganella morganii is a major contributor to abscess formation and gastrointestinal bacterial infections. It is also frequently detected as a multi-drug-resistant bacterium carried by snakes in their oral cavity.
Pseudomonas aeruginosa can lead to soft tissue infections in patients, including necrotizing cellulitis, folliculitis, dermatitis, urethritis, and osteomyelitis, and can induce immunosuppressive reactions in the snakebite site, potentially resulting in nosocomial infections. Infections caused by
Aeromonas hydrophila can result in hemorrhagic sepsis and diseases related to diarrhea. Other Gram-negative bacteria can cause various infection symptoms such as gastroenteritis, sepsis, respiratory tract infections, meningitis, diarrhea, fever, and soft tissue infections [
8,
9,
15,
23,
24,
25,
26,
27,
28]. The abundance of the aforementioned four species ranks high among the pathogens identified in our previous study [
16].
The bacterial species carried by the oral flora of snakes can impact not only humans but also infect the host snakes themselves [
21,
26,
29]. The oral cavity of reptiles may sustain wounds from struggling prey during feeding, tooth loss during hunting, or the ingestion of foreign substances such as artificial waste, strings, or fishing nets. These injuries can lead to swelling and inflammatory symptoms [
30,
31,
32]. Inadequate control of environmental temperature and humidity in snake captivity can induce immune suppression in snakes, making them susceptible to the invasion of pathogenic bacteria through the respiratory and gastrointestinal tracts, resulting in systemic infections or even death [
33,
34,
35]. Aerobic Gram-negative bacteria are generally considered the most pathogenic bacterial species for reptiles and are the dominant taxa detected in individuals with oral inflammation. Common genera include
Pseudomonas,
Aeromonas,
Proteus, and
Escherichia [
31,
33,
36]. Among them,
Pseudomonas aeruginosa,
Providencia rettgeri, and
Stenotrophomonas maltophilia are often dominant in the oral cavity of snakes with oral inflammation.
Aeromonas hydrophila has historically exhibited the highest occurrence rate and is a significant zoonotic pathogen causing infections and death in snakes, commonly distributed in the oral cavity or respiratory tract of snakes [
26,
33,
35,
37,
38]. To mitigate antibiotic resistance and enhance infection recovery rates, a comprehensive understanding of the carried bacterial species is essential for modifying subsequent treatment approaches and determining the appropriate types of antibiotics to administer [
39].
The occurrence of bacterial species and their quantities may vary due to factors such as snake species, season, habitat, snake health, feeding strategies, and prey types [
8,
9,
10,
14,
21,
24,
25,
26,
40]. Even closely related snake species may carry different oral bacterial species if they have different ranges of activity [
10,
14]. Many studies culture bacterial samples on agar plates before bacterial identification, but the cultured bacterial community may differ from the original community [
21,
41,
42,
43,
44,
45]. Non-cultivation methods can identify species that are not suitable for artificial cultivation [
16,
41].
Taiwan is home to six main venomous snake species:
Trimeresurus stejnegeri,
Protobothrops mucrosquamatus,
Naja atra,
Bungarus multicinctus,
Daboia siamensis, and
Deinagkistrodon acutus [
46,
47]. Annually, approximately 800 to 1000 people are bitten by venomous snakes in Taiwan [
48,
49]. Research on these venomous snakes often involves examining the clinical symptoms and wound tissue samples of snakebite patients to identify pathogenic bacterial species. Many previous studies focusing on snakebite patients in Taiwan have primarily used medical records from large hospitals in the northern or central regions for retrospective analysis [
23,
46,
50], with only a few focusing on the southern region [
49]. In wound samples from snakebite patients in Taiwan, Chen et al. [
23] and Huang et al. [
46] found that
M. morganii was the most prevalent among Gram-negative bacteria, followed by
A. hydrophila;
Enterococcus strains were the most common among Gram-positive bacteria. Chung et al. [
49] analyzed the bacterial composition of six major venomous snake oral samples by the culture method, revealing that the most abundant species among Gram-positive bacteria was
E. faecalis, while
P. aeruginosa and
Proteus vulgaris were the most common among Gram-negative bacteria. Further studies, without cultivating bacteria from oral samples, are necessary to comprehend the occurrence of pathogenic bacteria in the oral cavity of Taiwanese snakes. Additionally, there is still limited exploration of the associations between bacterial species and factors such as snake species, region, season, and the body condition of snakes. Moreover, there is a lack of research on the oral bacterial species in non-venomous snakes in Taiwan, which also harbor potential pathogenic bacteria.
This study aims to investigate the occurrence of pathogenic zoonotic bacterial species in the oral cavities of commonly encountered wild snake species in Kaohsiung, southern Taiwan. The targeted bacterial species include E. faecalis, a Gram-positive bacterium, as well as M. morganii, P. aeruginosa, and A. hydrophila, which are Gram-negative bacteria. Polymerase chain reaction (PCR) technology is employed to detect the presence of these four pathogenic microorganisms. Multiple logistic regression analyses are utilized to examine the association between the occurrence of the four bacterial species and various factors, including snake species, sex, snout-vent length, body mass, season, duration between capture and sampling, temperature and rainfall, altitude, artificial ground area around the capture site, and coexisting bacterial species. The findings of this study can serve as a foundation for antibiotic sensitivity testing of bacterial species and facilitate the development of veterinary medicine for wild snakes in Taiwan. This information can, therefore, be valuable in understanding the treatment approaches for secondary infections in humans suffering from snakebites or bacterial infections, whether in captive or wild snakes, stemming from oral microbes.
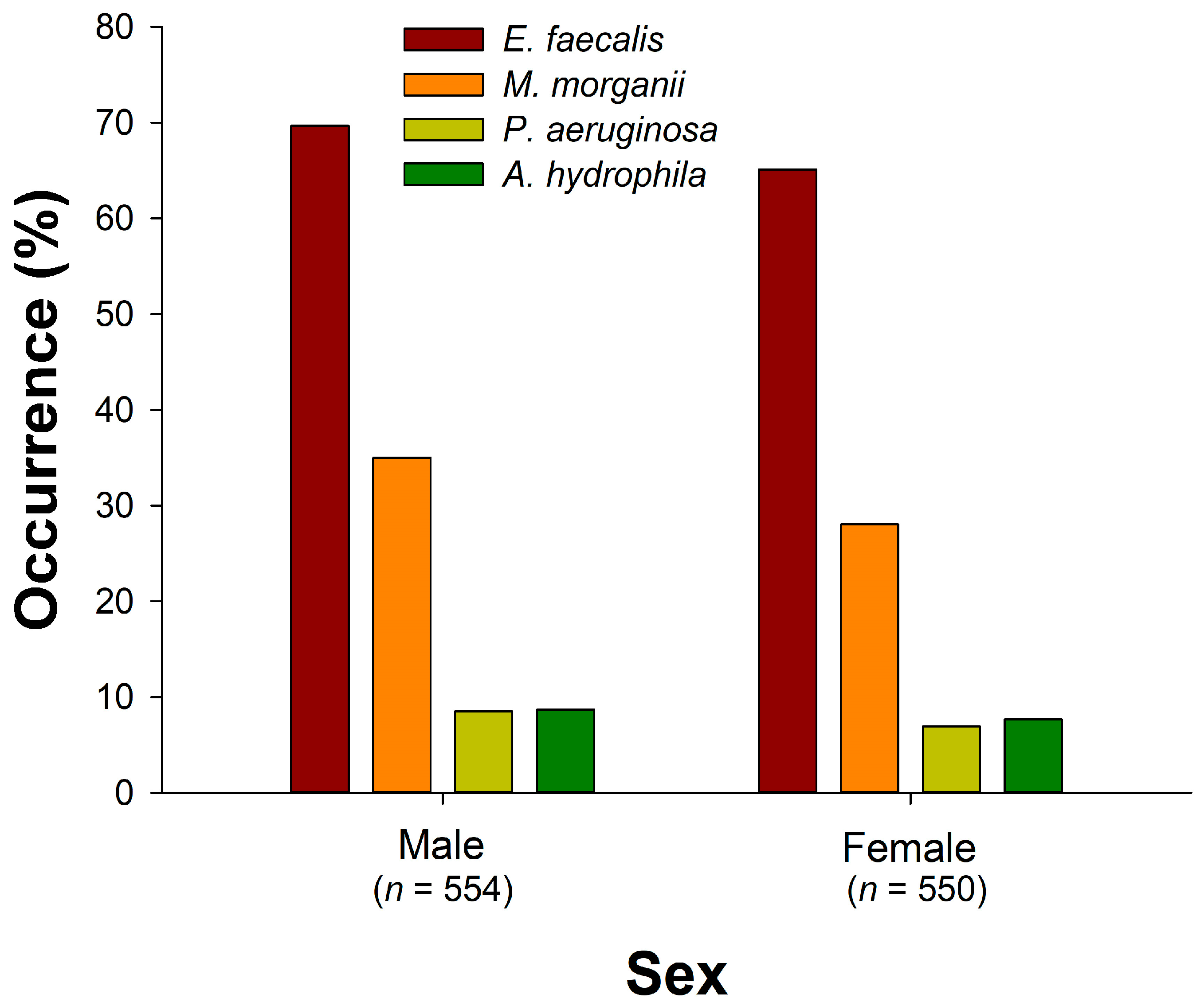
4. Discussion
The factors contributing to bacterial growth in a snake’s oral cavity, aside from venom, may encompass the balance of oral microbiota, the type of prey, environmental factors (humidity, temperature, and substrate composition), the immune system, oral hygiene, and the presence of injury or infection [
8,
9,
10,
14,
21,
24,
25,
26,
40]. This study demonstrates significant associations between the occurrence rates of the four bacterial species and various factors, as summarized in
Table 6. Notably, certain factors like sex, snout-vent length, scaled mass index, artificial ground area, rainfall, and the duration between capture and sampling did not exhibit a significant association with bacterial occurrences. In contrast, the presence of other bacterial species in the oral cavity consistently exhibited a positive association with the occurrence rates of the target bacteria, given that sampling of snakes with compromised health conditions or immune activity was avoided in this study. This finding suggests that pathogenic bacterial species may share growth environments with each other, as seen in previous research [
61]. It also implies that, whether dealing with secondary infections in humans following snake bites or bacterial infections in captive or wild snakes due to oral wounds, multiple pathogenic sources may contribute to the infections. Lin et al. [
62] and Yeh et al. [
63] indicated that
M. morganii and
E. faecalis were the most common sources of polymicrobial infections. Both
E. faecalis and
M. morganii can lead to bacteremia, while
A. hydrophila and
P. aeruginosa may result in urinary tract infections [
23,
46]. The presence of multiple pathogens in infections implies varying degrees of antibiotic resistance, underscoring the importance of thorough evaluation and testing of antibiotic sensitivity in the future. The associated factors for the occurrence rates of bacterial species are discussed as follows:
Some studies suggest that snake venom possesses antibacterial properties [
64,
65,
66,
67,
68], hypothesizing that the venom can protect venomous snakes from bacterial infections carried by prey during feeding. However, conflicting research supports the notion that venomous snakes display a greater diversity of oral bacteria compared to non-venomous ones [
21,
69]. In the context of this study, pit vipers (
P. mucrosquamatus and
T. stejnegeri) exhibit a significantly lower occurrence rate of
E. faecalis, a bacterium whose growth has been demonstrated to be inhibited by venom from vipers and cobras [
70]. In contrast, mildly venomous
B. kraepelini and venomous
B. multicinctus demonstrate a high occurrence rate of
E. faecalis,
P. aeruginosa, and
A. hydrophila. Furthermore, the occurrence rate of
M. morganii in the oral cavity of venomous
N. atra and
T. stejnegeri surpasses that in other snake species, consistent with prior research on patient wounds [
71] and the oral cavities of snakes [
16]. The reasons behind the heightened bacterial occurrence in the above (mildly) venomous species warrant further investigation. Whether the antibacterial properties of venom are related to only specific venom compositions also remains to be confirmed.
Our study reveals a positive correlation between the occurrence rate of
E. faecalis and environmental temperature, aligning with previous findings that
E. faecalis thrives in warmer temperatures [
72]. Since
E. faecalis predominantly inhabits the gastrointestinal tract of animals, the oral microbiota of snakes is influenced by the excretion of prey during feeding [
73]. The increased activity of snakes due to higher environmental temperatures may lead to an elevated frequency of predation [
60], indirectly contributing to the higher occurrence rate of
E. faecalis.
M. morganii exhibits the highest occurrence rate among Gram-negative bacteria in this study. This observation aligns with findings from both clinical research on patient wounds and snake oral microbiota studies [
23,
46,
50]. Bacteriomic analysis reveals that
M. morganii ranks among the 10 most abundant bacterial species in oropharyngeal samples of both
N. atra and
T. stejnegeri [
16].
M. morganii, known to inhabit the gastrointestinal tract of animals, is also widely distributed in the natural environment, including soil, water sources, and sewage [
74,
75]. The higher occurrence rate of
M. morganii in the oral cavity of male
N. atra and
B. multicinctus may be attributed to the larger activity range seen in males of some snake species [
76,
77], increasing the likelihood of contact with pathogens.
P. aeruginosa primarily inhabits human feces and sewage, with less occurrence in natural environments such as soil and streams [
78]. This study exhibited the lowest occurrence rate among bacterial species, contrary to previous research on Taiwanese snake oral bacteria that used culture-based methods, identifying
P. aeruginosa as the Gram-negative species with the highest occurrence rate [
49]. This discrepancy may stem from the limitation that microbial compositions cultured on agar plates may not entirely represent the original oral microbiota, as indicated by several studies [
21,
41,
42,
43,
44,
45]. Studies have shown that
P. aeruginosa outbreaks are more extensive in hospital areas than in non-hospital settings [
78]. This suggests that the
P. aeruginosa identified in clinical studies, particularly in patients with wound infections [
23], may predominantly originate from nosocomial infections. Additionally, previous research investigating the seasonal association of
P. aeruginosa using human samples demonstrated a peak in its occurrence during cooler months (October to March) [
79], which aligns with our findings indicating a higher occurrence rate of
P. aeruginosa in winter.
This study has several limitations: (1) The investigation did not analyze the impact of dietary factors on the incidence rates of bacteria in the sampled snake species due to their complex and overlapping feeding habits. Subsequent research could concentrate on snake species with simpler and non-overlapping diets to ascertain whether snake feeding habits significantly influence bacterial occurrence rates. (2) Previous studies [
31,
33,
36] have identified Gram-negative bacteria, particularly
P. aeruginosa,
P. rettgeri, and
S. maltophilia, as dominant strains in the oral cavity of snakes with oral inflammation. However, our study did not sample the oral bacterial community of unhealthy or sick snakes. Future studies should include samples from snakes exhibiting infectious symptoms to explore variations in oral microbiomes under different health conditions. (3) The bacterial strains selected for this study were aerobic or facultatively anaerobic. Future research on the oral bacterial communities of wild snakes should also consider investigating other types of pathogenic bacteria, including anaerobic strains such as
B. fragilis and
Clostridium spp.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}