
Antibiofilm Activity and Biocompatibility of Temporin-SHa: A Promising Antimicrobial Peptide for Control of Fluconazole-Resistant Candida albicans

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
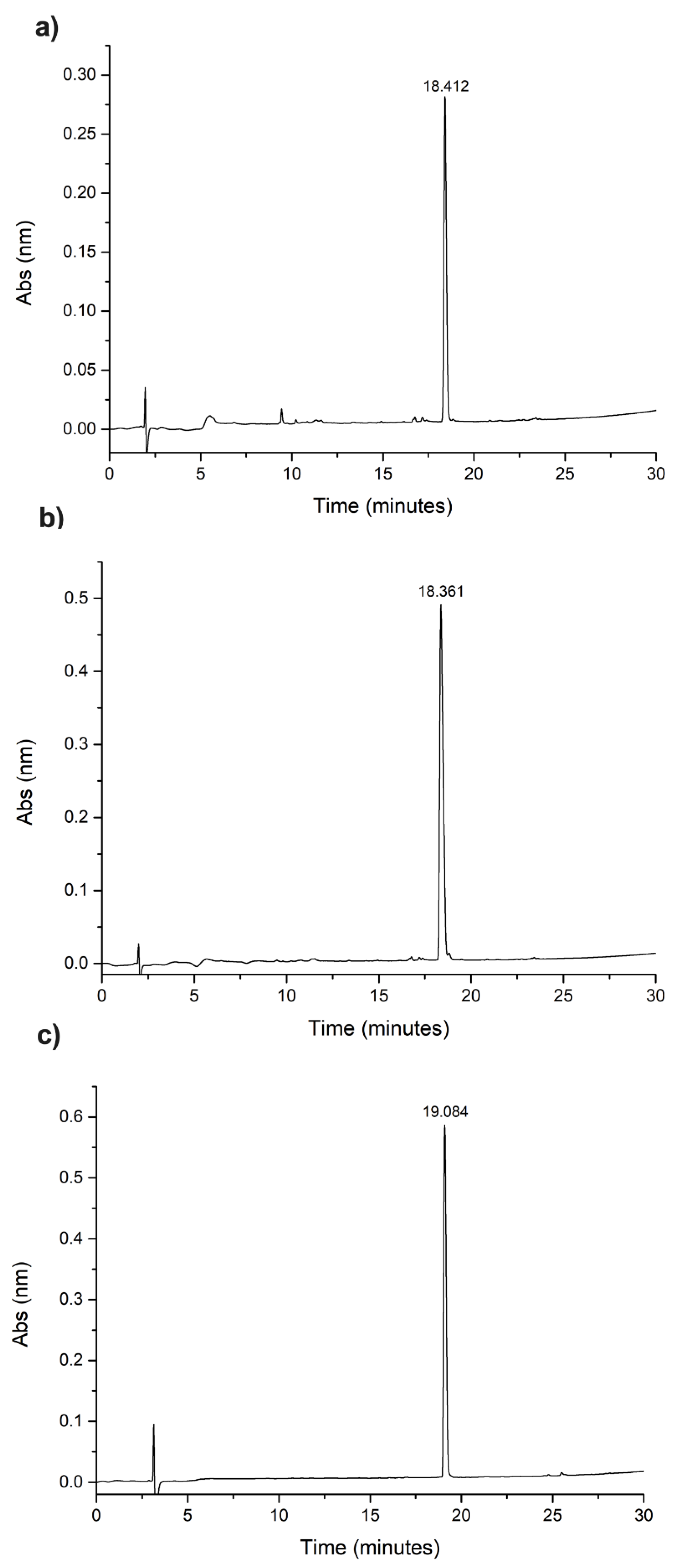
2.1. Synthesis, Cleavage, and Purification of Peptides Hylin−a1, KR−12-a5, and Temporin-SHa
2.2. Antimicrobial Activity of AMPs on Planktonic Cultures of C. albicans
2.2.1. Strains, Microorganism Growth, and Culture Conditions
2.2.2. Survival Curve, Minimum Inhibitory Concentration (MIC), and Minimum Fungicide Concentration (MFC) of AMPs on Planktonic Cultures of C. albicans
2.3. Cytotoxicity Evaluation of Hylin−a1 and KR−12-a5 on Oral Cell Cultures in Normal Oral Keratinocytes (NOK-si) and Human Gingival Fibroblasts (FGH)
2.3.1. Cell Line and Cell Culture
2.3.2. Cell Viability Analysis by alamarBlue™
2.4. Antimicrobial Activity of AMPs against C. albicans Biofilm
2.4.1. Quantification of Total Biofilm Biomass of C. albicans (CaS and CaR) with Crystal Violet Dye
2.4.2. Efficacy of AMPs in the Inhibition of Biofilm Formation and against the Biofilm Formed
2.5. Data Analysis
3. Results
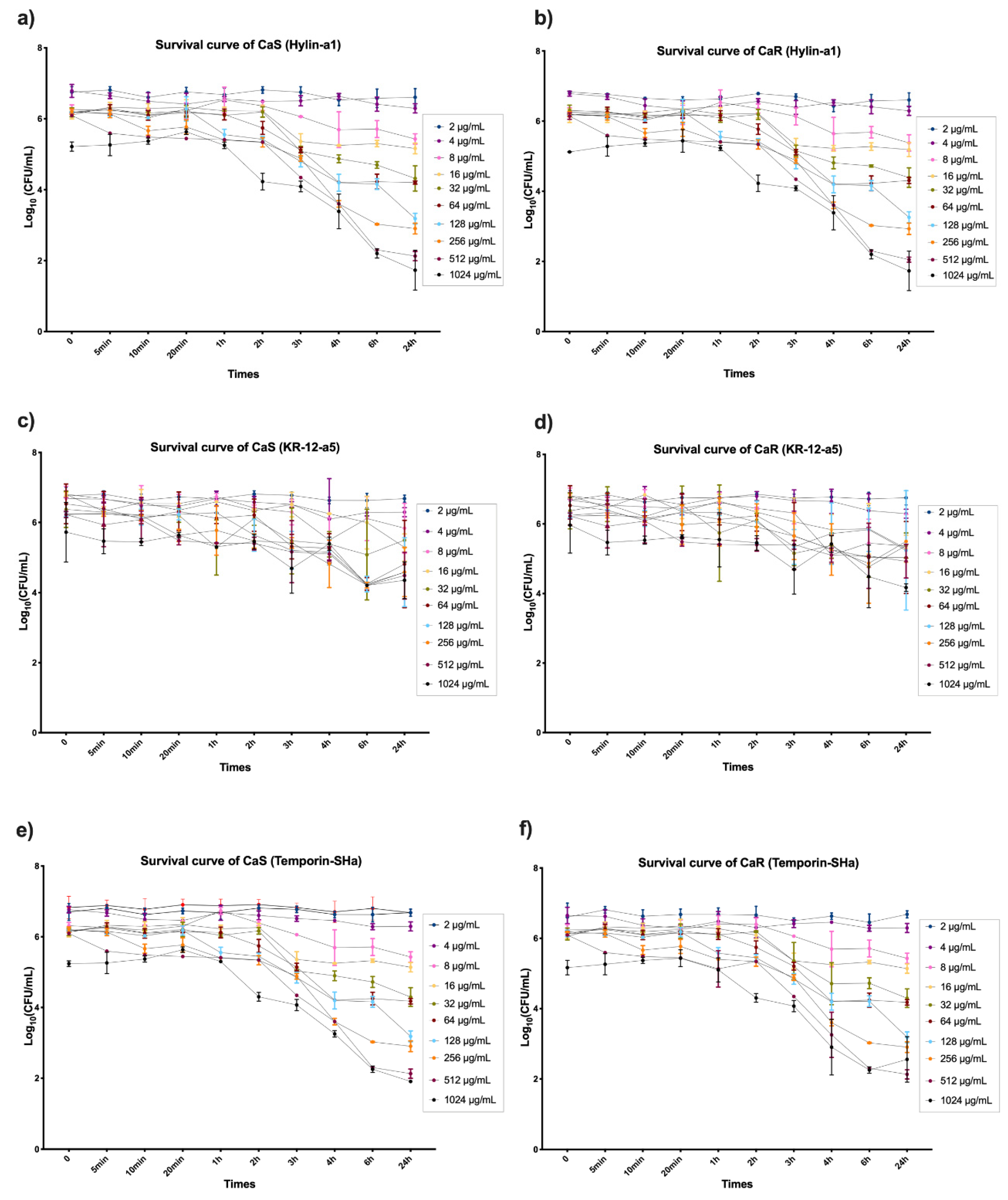
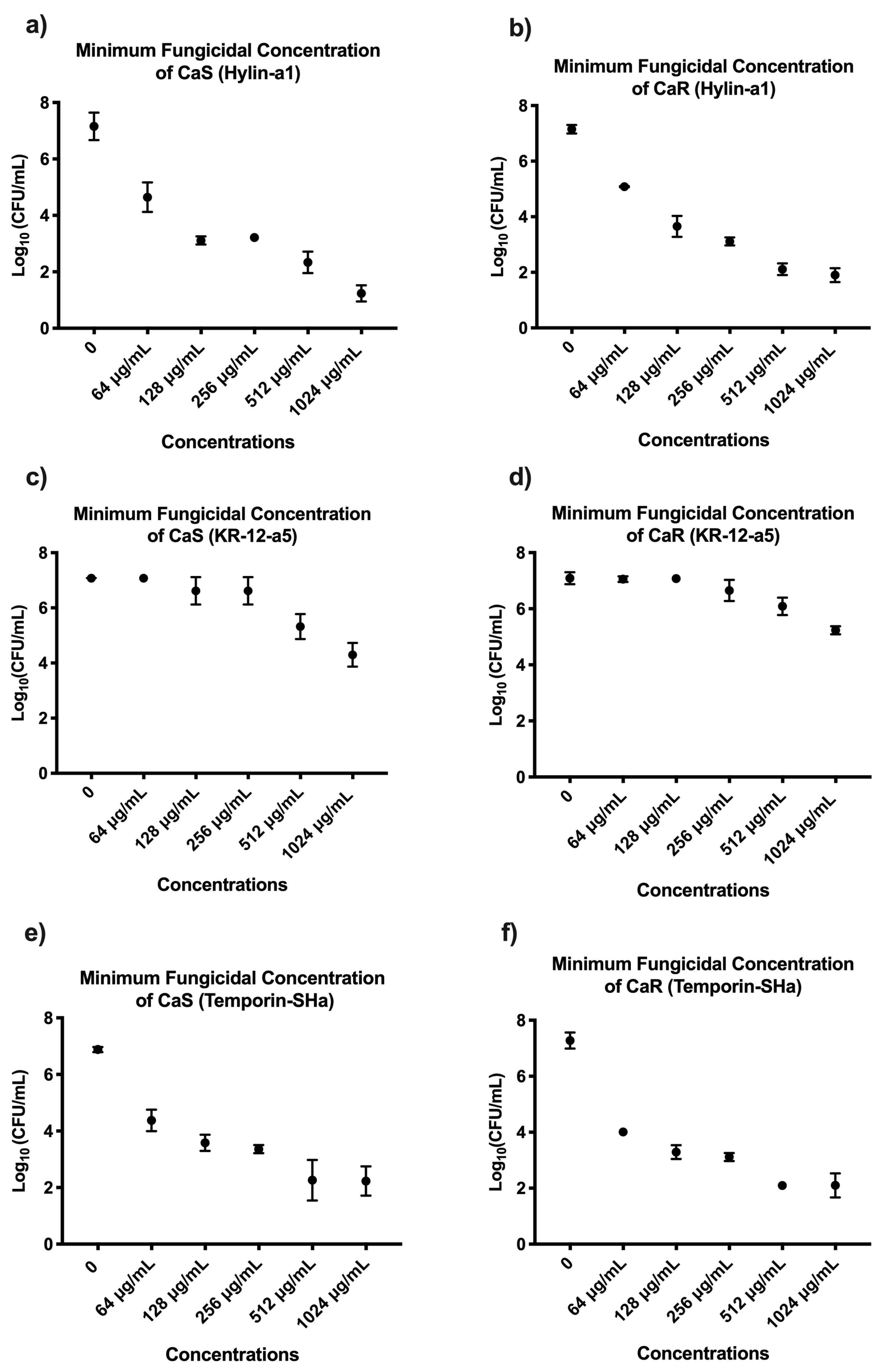
3.1. Survival Curve, Minimum Inhibitory Concentration (MIC), and Minimum Fungicide Concentration (MFC) of AMPs on Planktonic Cultures of C. albicans
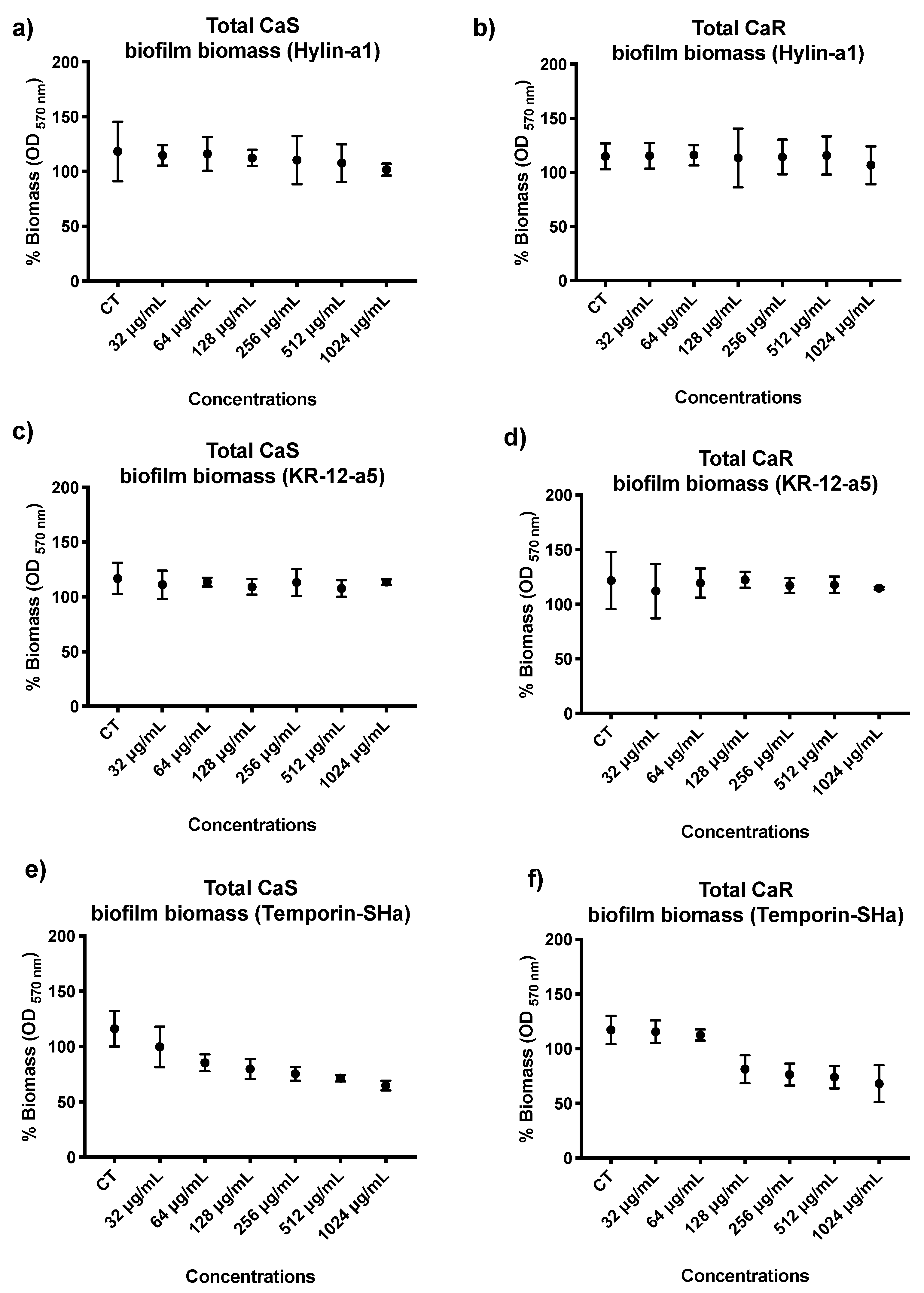
3.2. Quantification of Total Biofilm Biomass of C. albicans (CaS and CaR)
3.3. Efficacy of AMPs in the Inhibition of Biofilm Formation and against the Biofilm Formed
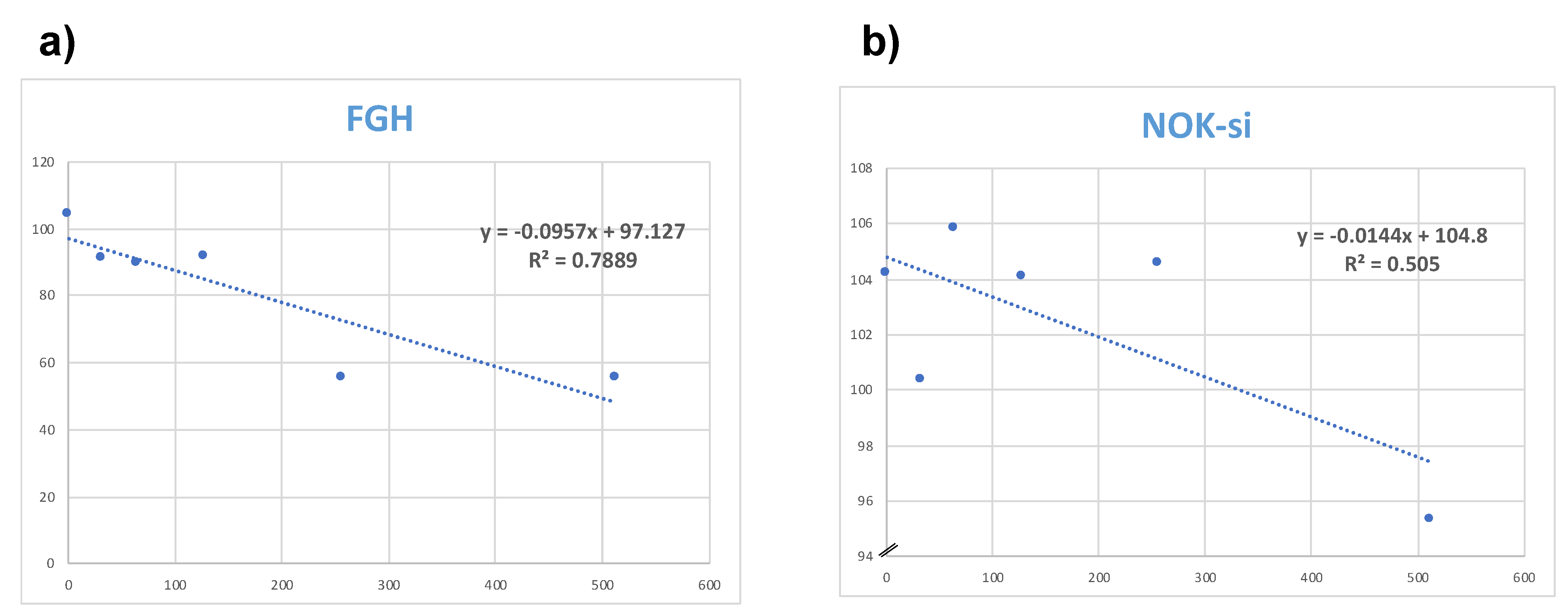
3.4. Cytotoxicity Evaluation of Hylin−a1 and KR−12-a5 in Oral Cell Cultures on Normal Oral Keratinocytes (NOK-si) and Human Gingival Fibroblasts (FGH)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- d’Enfert, C. Hidden Killers: Persistence of Opportunistic Fungal Pathogens in the Human Host. Curr. Opin. Microbiol. 2009, 12, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Finkel, J.S.; Mitchell, A.P. Genetic Control of Candida albicans Biofilm Development. Nat. Rev. Microbiol. 2011, 9, 109–118. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial Bloodstream Infections in US Hospitals: Analysis of 24,179 Cases from a Prospective Nationwide Surveillance Study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.S.; Carlisle, P.L.; Kadosh, D. Coevolution of Morphology and Virulence in Candida Species. Eukaryot. Cell 2011, 10, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Santos Fontenelle, R.O.; Brito, E.H.S.; Morais, S.M. Biofilm of Candida albicans: Formation, Regulation and Resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Van Dijck, P.; Sjollema, J.; Camue, B.P.A.; Lagrou, K.; Berman, J.; d’Enfert, C.; Andes, D.R.; Arendrup, M.C.; Brakhage, A.A.; Calderone, R.; et al. Methodologies for In Vitro and In Vivo Evaluation of Efficacy of Antifungal and Antibiofilm Agents and Surface Coatings against Fungal Biofilms. Microb. Cell 2018, 5, 300–326. [Google Scholar] [CrossRef] [PubMed]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between Resistance, Tolerance and Persistence to Antibiotic Treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, P.; Ferrari, S.; Coste, A.T. Antifungal Resistance and New Strategies to Control Fungal Infections. Int. J. Microbiol. 2012, 2012, 713687. [Google Scholar] [CrossRef]
- Fan, F.; Liu, Y.; Liu, Y.; Lv, R.; Sun, W.; Ding, W.; Cai, Y.; Li, W.; Liu, X.; Qu, W. Candida albicans Biofilms: Antifungal Resistance, Immune Evasion, and Emerging Therapeutic Strategies. Int. J. Antimicrob. Agents 2022, 60, 106673. [Google Scholar] [CrossRef]
- Dias, L.M.; Ferrisse, T.M.; Medeiros, K.S.; Cilli, E.M.; Pavarina, A.C. Use of Photodynamic Therapy Associated with Antimicrobial Peptides for Bacterial Control: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2022, 23, 3226. [Google Scholar] [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Raja, Z.; André, S.; Abbassi, F.; Humblot, V.; Lequin, O.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D.; et al. Insight into the Mechanism of Action of Temporin-SHa, a New Broad-Spectrum Antiparasitic and Antibacterial Agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [PubMed]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.A.; Farooq, S.; Ali, S.A.; Hameed, A.; Choudhary, M.I.; Shaheen, F. New Analogs of Temporin-LK1 as Inhibitors of Multidrug-Resistant (MDR) Bacterial Pathogens. Synth. Commun. 2018, 48, 1172–1182. [Google Scholar] [CrossRef]
- Kang, J.; Dietz, M.J.; Li, B. Antimicrobial Peptide LL-37 Is Bactericidal against Staphylococcus Aureus Biofilms. PLoS ONE 2019, 14, e0216676. [Google Scholar] [CrossRef] [PubMed]
- Patrzykat, A.; Friedrich, C.L.; Zhang, L.; Mendoza, V.; Hancock, R.E.W. Sublethal Concentrations of Pleurocidin-Derived Antimicrobial Peptides Inhibit Macromolecular Synthesis in Escherichia coli. Antimicrob. Agents Chemother. 2002, 46, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Epand, R.F.; Epand, R.M.; Wang, G. Structural Location Determines Functional Roles of the Basic Amino Acids of KR-12, the Smallest Antimicrobial Peptide from Human Cathelicidin LL-37. RSC Adv. 2013, 3, 19560. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.S.; Ferreira, T.C.G.; Cilli, E.M.; Crusca, E.; Mendes-Giannini, M.J.S.; Sebben, A.; Ricart, C.A.O.; Sousa, M.V.; Fontes, W. Hylin A1, the First Cytolytic Peptide Isolated from the Arboreal South American Frog Hypsiboas Albopunctatus (“Spotted Treefrog”). Peptides 2009, 30, 291–296. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial Peptides from Ranid Frogs: Taxonomic and Phylogenetic Markers and a Potential Source of New Therapeutic Agents. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2004, 1696, 1–14. [Google Scholar] [CrossRef]
- Dias, L.M.; Klein, M.I.; Ferrisse, T.M.; Medeiros, K.S.; Jordão, C.C.; Bellini, A.; Pavarina, A.C. The Effect of Sub-Lethal Successive Applications of Photodynamic Therapy on Candida albicans Biofilm Depends on the Photosensitizer. J. Fungi 2023, 9, 111. [Google Scholar] [CrossRef]
- Hansen, P.R.; Oddo, A. Fmoc Solid-Phase Peptide Synthesis. In Peptide antibodies: Methods in Molecular Biology; Humana Press: New York, NY, USA, 2015; pp. 33–50. [Google Scholar] [CrossRef]
- Kaiser, E.; Colescott, R.L.; Bossinger, C.D.; Cook, P.I. Color Test for Detection of Free Terminal Amino Groups in the Solid-Phase Synthesis of Peptides. Anal. Biochem. 1970, 34, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Panariello, B.H.D.; Klein, M.I.; Pavarina, A.C.; Duarte, S. Inactivation of genes TEC1 and EFG1 in Candida albicans influences extracellular matrix composition and biofilm morphology. J. Oral Microbiol. 2017, 17, 1385372. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Pereira, C.A.; Gorayb-Pereira, A.L.; Menezes Noveletto, J.V.; Jordão, C.C.; Pavarina, A.C. Zerumbone Disturbs the Extracellular Matrix of Fluconazole-Resistant Candida albicans Biofilms. J. Fungi 2023, 9, 576. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K. Erratum for Humphries et al.,“CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests”. J. Clin. Microbiol. 2023, 61, e00739-23. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P. Antifungal Susceptibility of Yeasts and Filamentous Fungi by CLSI Broth Microdilution Testing. In Antifungal Drug Resistance: Methods and Protocols; Springer: New York, NY, USA, 2023; pp. 3–16. [Google Scholar] [CrossRef]
- Kragh, K.N.; Alhede, M.; Rybtke, M.; Stavnsberg, C.; Jensen, P.Ø.; Tolker-Nielsen, T.; Whiteley, M.; Bjarnsholt, T. The Inoculation Method Could Impact the Outcome of Microbiological Experiments. Appl. Environ. Microbiol. 2018, 84, e02264-17. [Google Scholar] [CrossRef] [PubMed]
- da Silva Pimentel, B.N.A.; de Foggi, C.C.; Barbugli, P.A.; de Oliveira, R.C.; de Avila, E.D.; Longo, E.; Vergani, C.E. Antifungal Activity and Biocompatibility of α-AgVO3 Microcrystals: A Promising Material against Oral Candida Disease. Mater. Sci. Eng. C 2020, 108, 110405. [Google Scholar] [CrossRef]
- ISO 10993-5:2009; Biological Evaluation of Medical Devices—Part 5: Tests for in Vitrocytotoxicity. International Organization for Standardization (ISO): Geneva, Switzerland, 2009. Available online: https://www.iso.org/standard/36406.html (accessed on 18 November 2023).
- Viana de Sousa, T.; Carolina Jordão, C.; Augusto Abreu-Pereira, C.; Gorayb Pereira, A.L.; Barbugli, P.A.; Klein, M.I.; Pavarina, A.C. Hydrogen peroxide enhances the efficacy of photodynamic therapy against Candida albicans biofilms. Biofouling. 2023, 39, 94–109. [Google Scholar] [CrossRef]
- Mishra, P.; Pandey, C.; Singh, U.; Keshri, A.; Sabaretnam, M. Selection of Appropriate Statistical Methods for Data Analysis. Ann. Card. Anaesth. 2019, 22, 297. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2019, 27, 4–16. [Google Scholar] [CrossRef]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial Peptides and Their Interaction with Biofilms of Medically Relevant Bacteria. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 1044–1060. [Google Scholar] [CrossRef]
- Brunet, K.; Verdon, J.; Ladram, A.; Arnault, S.; Rodier, M.-H.; Cateau, E. Antifungal Activity of [K3]Temporin-SHa against Medically Relevant Yeasts and Moulds. Can. J. Microbiol. 2022, 68, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Olleik, H.; Baydoun, E.; Perrier, J.; Hijazi, A.; Raymond, J.; Manzoni, M.; Dupuis, L.; Pauleau, G.; Goudard, Y.; de La Villéon, B.; et al. Temporin-SHa and Its Analogs as Potential Candidates for the Treatment of Helicobacter Pylori. Biomolecules 2019, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, K.; Dawadi, B.R.; Shrestha, L.B. Role of Biofilm in Bacterial Infection and Antimicrobial Resistance. J. Nepal. Med. Assoc. 2022, 60, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wade, D.; Kurosaka, K.; Wang, Z.Y.; Oppenheim, J.J.; Yang, D. Temporin A and Related Frog Antimicrobial Peptides Use Formyl Peptide Receptor-Like 1 as a Receptor to Chemoattract Phagocytes. J. Immunol. 2004, 173, 2652–2659. [Google Scholar] [CrossRef] [PubMed]
- Jordão, C.C.; Klein, M.I.; Carmello, J.C.; Dias, L.M.; Pavarina, A.C. Consecutive Treatments with Photodynamic Therapy and Nystatin Altered the Expression of Virulence and Ergosterol Biosynthesis Genes of a Fluconazole-Resistant Candida albicans in Vivo. Photodiagnosis Photodyn. Ther. 2021, 33, 102155. [Google Scholar] [CrossRef] [PubMed]
- Panariello, B.H.D.; Klein, M.I.; Mima, E.G.D.O.; Pavarina, A.C. Fluconazole Impacts the Extracellular Matrix of Fluconazole-Susceptible and -Resistant Candida albicans and Candida Glabrata Biofilms. J. Oral. Microbiol. 2018, 10, 1476644. [Google Scholar] [CrossRef] [PubMed]
- Caiaffa, K.S.; Massunari, L.; Danelon, M.; Abuna, G.F.; Bedran, T.B.L.; Santos-Filho, N.A.; Spolidorio, D.M.P.; Vizoto, N.L.; Cilli, E.M.; Duque, C. KR−12-A5 Is a Non-Cytotoxic Agent with Potent Antimicrobial Effects against Oral Pathogens. Biofouling 2017, 33, 807–818. [Google Scholar] [CrossRef]
- Jiao, Y.; Tay, F.R.; Niu, L.; Chen, J. Advancing Antimicrobial Strategies for Managing Oral Biofilm Infections. Int. J. Oral. Sci. 2019, 11, 28. [Google Scholar] [CrossRef]
- Cao, Y.; Yin, H.; Wang, W.; Pei, P.; Wang, Y.; Wang, X.; Jiang, J.; Luo, S.-Z.; Chen, L. Killing Streptococcus mutans in Mature Biofilm with a Combination of Antimicrobial and Antibiofilm Peptides. Amino Acids 2020, 52, 1–14. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, X.; Jiao, Z.; Peng, J.; Zhou, L.; Yang, L.; Huang, M.; Tian, C.; Guo, G. The Antimicrobial Peptide AMP-17 Derived from Musca Domestica Inhibits Biofilm Formation and Eradicates Mature Biofilm in Candida albicans. Antibiotics 2022, 11, 1474. [Google Scholar] [CrossRef]
- Galdiero, E.; de Alteriis, E.; De Natale, A.; D’Alterio, A.; Siciliano, A.; Guida, M.; Lombardi, L.; Falanga, A.; Galdiero, S. Eradication of Candida albicans Persister Cell Biofilm by the Membranotropic Peptide GH625. Sci. Rep. 2020, 10, 5780. [Google Scholar] [CrossRef] [PubMed]
- Vignoli Muniz, G.S.; De la Torre, L.I.; Duarte, E.L.; Lorenzón, E.N.; Cilli, E.M.; Balan, A.; Lamy, M.T. Interaction of Synthetic Antimicrobial Peptides of the Hylin A1 Family with Models of Eukaryotic Structures: Zwitterionic Membranes and DNA. Biochem. Biophys. Rep. 2020, 24, 100827. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Mukherjee, P.K.; Leidich, S.D.; Faddoul, F.F.; Hoyer, L.L.; Douglas, L.J.; Ghannoum, M.A. Antifungal Resistance of Candidal Biofilms Formed on Denture Acrylic In Vitro. J. Dent. Res. 2001, 80, 903–908. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classifications | % of Viability |
|---|---|
| non-cytotoxic | <25% |
| slightly cytotoxic | 25–50% |
| moderately cytotoxic | 50–75% |
| severely cytotoxic | >75% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, L.M.; Cilli, E.M.; Medeiros, K.S.; Brasil, M.C.O.d.A.; Marin, L.M.; Siqueira, W.L.; Pavarina, A.C. Antibiofilm Activity and Biocompatibility of Temporin-SHa: A Promising Antimicrobial Peptide for Control of Fluconazole-Resistant Candida albicans. Microorganisms 2024, 12, 99. https://doi.org/10.3390/microorganisms12010099
Dias LM, Cilli EM, Medeiros KS, Brasil MCOdA, Marin LM, Siqueira WL, Pavarina AC. Antibiofilm Activity and Biocompatibility of Temporin-SHa: A Promising Antimicrobial Peptide for Control of Fluconazole-Resistant Candida albicans. Microorganisms. 2024; 12(1):99. https://doi.org/10.3390/microorganisms12010099
Chicago/Turabian StyleDias, Luana Mendonça, Eduardo Maffud Cilli, Karine Sousa Medeiros, Maria Carolina Oliveira de Arruda Brasil, Lina Maria Marin, Walter L. Siqueira, and Ana Claudia Pavarina. 2024. "Antibiofilm Activity and Biocompatibility of Temporin-SHa: A Promising Antimicrobial Peptide for Control of Fluconazole-Resistant Candida albicans" Microorganisms 12, no. 1: 99. https://doi.org/10.3390/microorganisms12010099